
城市公园生境类型对鸟类群落的影响
2015-02-06 03:54丁由中吴时英唐海明李宏庆王小明王正寰
生态学报 2015年12期
杨 刚,王 勇,许 洁, 丁由中,吴时英,唐海明, 李宏庆,王小明, 马 波,王正寰,*
1. 华东师范大学生命科学学院, 上海 200062 2 华东师范大学上海市城市化过程与恢复重点实验室, 上海 200241 3 上海市浦东新区林业站, 上海 201210 4 上海科技馆,上海 200127 5 上海市野生动植物保护管理站, 上海 200023
城市公园生境类型对鸟类群落的影响
杨 刚1,2,王 勇1,2,许 洁1,2, 丁由中1,2,吴时英3,唐海明3, 李宏庆1,王小明4, 马 波5,王正寰1,2,*
1. 华东师范大学生命科学学院, 上海 200062 2 华东师范大学上海市城市化过程与恢复重点实验室, 上海 200241 3 上海市浦东新区林业站, 上海 201210 4 上海科技馆,上海 200127 5 上海市野生动植物保护管理站, 上海 200023
2011 年12 月—2012 年11 月,在上海世纪公园和滨江森林公园对鸟类群落和植物群落进行调查,通过对12 个植被变量进行主成分分析,将两个公园分为8 种不同的生境类型。结果表明:2 个公园生境构成存在显著差异,滨江森林公园灌木层植物发达的生境(Habitat with developed shrub layer, S型)以及灌木层和地被层植物都发达的生境(Habitat with developed tree layer and shrub layer, T+S型)数量显著多于世纪公园,世纪公园地被层发达的生境(Habitat with developed ground cover layer, G型)以及乔木层和地被层植物都发达的生境(Habitat with developed tree layer and ground cover layer, T+G型)数量显著多于滨江森林公园。世纪公园不同生境中鸟种数差异显著,而滨江森林公园中差异不显著。2 个公园有24 种共有鸟种,对共有鸟种生境利用率的配对t检验结果表明,滨江森林公园鸟类生境利用率显著高于世纪公园。对2 个公园共有生境类型中鸟种数进行分析,发现滨江森林公园鹟科(Muscicapidae)鸟类种数显著大于世纪公园。根据以上结果,上海城市公园不同生境类型对鸟类群落结构存在显著影响。因此,建议在规划和建设大型城市公园时,应构建植被分层结构复杂的生境,多样化种植各类乔木,林下多样化搭配灌木。在保留供游客休憩草坪区域的同时种植各类草本植物,以此提高鸟类生境利用率,增加城市公园的鸟类多样性。
城市公园;生境类型;鸟类群落;植被结构;上海
城市化致使生物多样性丧失是一个全球性的生态问题[1]。上海作为中国城市化率最高的城市[2],城市化导致的生物群落组成变化尤其明显[3]。鸟类作为生物多样性的重要组成部分,是城市生态系统健康的重要指标类群[4]。研究城市化对城市鸟类群落组成的影响已成为城市生态学研究的热点[5]。
城市公园绿地是城市鸟类的主要栖息地[6],为鸟类直接或间接的提供食物。国内外的一些学者已经开始对城市鸟类和城市绿地的关系进行了探讨和研究。在景观尺度上,Sandstrom等[7]和McKinney[8]的研究表明,城市化导致鸟类群落的组成发生变化,自然林地中常见的食虫鸟类、筑地面巢或者树洞巢鸟类沿城市化梯度种类和数量都呈减少趋势。陈水华等[9]对城市鸟类群落组成的研究表明,园林鸟类群落呈现出不完全的嵌套格局, 分布于物种数较少的园林中的物种多数也分布在物种数较多的园林中。王彦平等[10]的研究也证实了杭州20 块绿地中60 种繁殖鸟和越冬鸟类显著符合嵌套结构。在微生境尺度上,林地成熟度以及灌木盖度、高度对鸟类群落的重要性明显[11],而林下带的洁净度与鸟类多度呈显著的负相关[12]。在公园树种的搭配上,Melles等[13]指出针叶林和浆果灌木与城市公园鸟类丰富度存在显著正相关。Fontana等[14]研究表明,城市绿地植被结构,尤其是乔木结构复杂度与鸟类丰富度和多样性呈显著正相关。陈水华等[15]对杭州公园鸟类群落的研究表明,杭州城市鸟类对园林栖息地具有较强的选择性,园林栖息地间的异质性以及鸟类物种与栖息地结构的密切关系是园林鸟类选择性分布的主要原因。
上海鸟类资源较丰富,从20 世纪初至今共整理记录了438 种鸟类,其中85.2%的鸟类为迁徙鸟类[16],因此对上海野生鸟类资源的保护不仅对本地生物多样性的提高存在重要意义,而且保护了所有迁徙路线途径上海的野生鸟类。上海绿地的建设近年来不断加快,截止2012 年,上海公共绿地面积人均13.1 m2,森林覆盖率达到12.58%[17]。然而一直以来,公园的植被配置主要考虑景观和休闲的需要[18],其作为野生动物栖息地的生态服务功能未得到足够关注。因此,研究上海城市公园作为野生鸟类栖息地的植被结构是有效改善高度城市化地区鸟类多样性水平的重要基础研究,对于城市公园作为鸟类栖息地生态服务功能的实现具有现实的指导意义。
本研究选择上海市2 个典型的大型城市公园——上海世纪公园和上海滨江森林公园作为研究地点,运用数值分类的方法对两个公园的生境类型进行划分,旨在解决如下3个科学问题:(1)鸟种数和鸟类数量在2个公园的不同生境类型中是否存在显著差异;(2)2 个公园共有鸟种的生境利用率是否存在显著差异;(3)2 个公园共有生境类型中鸟类群落组成是否存在显著差异。通过回答以上问题进而归纳出生境植被组成差异对鸟类群落结构的影响。该研究将为大型城市公园合理配置植被,提高其作为城市野生动物尤其是野生鸟类栖息地的生态服务功能提供理论基础和建议。
1 研究方法
1.1 研究地点的选择
上海(31°14′N, 121°29′E) 属于亚热带季风气候,是中国的经济和工业中心,2012年上海市城市化率已超过89%,常住人口达到2300 多万[2]。进入21 世纪以来,上海不断加强城市绿地建设的力度,公园绿地面积从2008 年的1153 hm2增长到2010 年的1687 hm2[17]。上海城市公园建设有两种主要类型,一种是在苗圃林地基础上改建而成的森林公园,另一种是根据园林景观要求兴建而成的公园。滨江森林公园和世纪公园是这两类公园的典型代表。
滨江森林公园位于上海市浦东新区高桥镇,占地面积约120 hm2,由始建于1985 年的三岔港苗圃改建而成,并在2007 年对公众开放。常见的木本植物包括:落羽杉 (Taxodiumdistichum)、广玉兰(Magnoliagrandiflora)、樟(Cinnamomumcamphora)、女贞(Ligustrumlucidum)、枫杨(Pterocaryastenoptera)、池杉(T.ascendens)、棕榈(Trachycarpusfortunei)、海桐(Pittosporumtobira)、八角金盘(Fatsiajaponica)、罗汉松(Podocarpusmacrophyllus)等。常见的草本包括:狗牙根(Cynodondactylon)、诸葛菜(Orychophragmus violaceus)、红花酢浆草(Oxaliscorymbosa)、虎耳草(Saxifragastolonifera)等。
世纪公园位于上海市浦东新区花木行政文化中心,占地面积约140 hm2,于2000 年公园对外开放。常见的木本植物包括:樟(C.camphora)、落羽杉(T.distichum)、二球悬铃木(Platanusacerifolia)、枫杨(P.stenoptera)、榉树(Zelkovaserrata)、黄连木(Pistaciachinensis)、槐(Sophorajaponica)、罗汉松(P.macrophyllus)、湿地松(Pinuselliottii)、夹竹桃(Neriumindicum)等。常见的草本植物包括:狗牙根(C.dactylon)、早熟禾(Poaannua)、乌蔹莓(Cayratiajaponica)、小蓬草(Conyzacanadensis)等。
1.2 样点设置
根据系统取样原则[19],通过在数字地图上设置抽样网格的方法,将网格的交叉点作为调查样点。具体的做法是在2 个公园中分别随机设置抽样起点,使用ArcGIS 9.2生成抽样网格和备选样点。然后,分别从2 个公园的备选样点中随机抽取20 个作为该公园最终的调查样点。抽取的随机样点中只有避开道路、建筑以及其他公园中不可到达区域、且间距≥100 m的样点可以作为调查样点。因此,上述随机抽样可进行多轮,以最终确定调查样点。本研究涉及的植被样方调查和鸟类调查均以这些调查样点作为样方中心。
1.3 植被调查
2012 年6 月—8 月,采用样方法对2 个公园进行植被调查。以调查样点为中心,设置20 m×20 m的大样方,对样方内所有乔木层植物的种类和数量进行统计。在20 m×20 m的样方内随机选择5 个4 m×4 m的中样方和5 个1 m×1 m小样方。在中样方中对灌木层植物的种类和数量进行调查,在小样方中对地被层植物的种类和数量进行调查。植被的分层结构依据植物地上部分高度划分(乔木层h(株高)>2.5 m;灌木层 0.5 m≤h≤2.5 m;地被层h<0.5 m),然后收集如下植被变量信息:
乔木层盖度(COVt),大样方内乔木层植物的盖度;
灌木层盖度(COVs),中样方内灌木层植物的盖度除以中样方数目;
乔木层多样性(DIVt),大样方内乔木层植物的多样性指数,多样性指数采用Shannon-Wiener指数,计算公式为:
灌木层多样性(DIVs),中样方内灌木层植物的多样性指数,计算公式同乔木层多样性指数;
乔木层丰富度(RICt),大样方内乔木层植物的物种数;
灌木层丰富度(RICs),中样方内灌木层植物的物种数;
地被层丰富度(RICg),小样方内地被层植物的物种数;
乔木层均匀度(EVEt),大样方内乔木层植物的均匀度指数,计算公式为:
灌木层均匀度(EVEs),中样方内灌木层植物的均匀度指数,计算公式同乔木层均匀度指数;
乔木层多度(ABUt),大样方内乔木层植物的个体数目;
灌木层多度(ABUs),中样方内灌木层植物的个体数目;
地被层多度(ABUg),小样方内地被层植物的个体数目。
按照20 个大样方覆盖面积计算,世纪公园植被调查抽样强度为3.47%、滨江森林公园为3.41%。
1.4 鸟类调查
在2011 年12 月至2012 年的11 月间,选择晴朗无风的工作日,在07:00—10:00和16:00—18:00 两个时间段,采用样点法进行鸟类调查。调查时,研究人员到达样点后先静止5 min,然后以样点为中心,记录25 m半径内所有看到、听到的鸟类和数量,每个样点观察时间为5 min,整个研究期间调查人员固定以控制系统误差[20]。在各样点收集如下鸟类数据:
鸟种数(RICb),调查所记录到的鸟类物种数;
鸟类数量(ABUb),调查所记录到的鸟类个体数,单位为次/只;
每种生境类型的平均鸟类数量(AABU),每种生境的所有鸟类个体数除以该类型生境的数目。
1.5 数据分析
1.5.1 两个公园生境类型的数值分类和鸟类分布差异分析
用主成分分析(PCA)对两个公园40 个样方的植被变量进行降维分析。PCA处理之前对所有数据进行标准化。采用最大方差法旋转,截取特征值(Eigenvalue)≥1的主成分用于降维[11]。在各主成分中选取载荷量(loading)绝对值≥0.7的变量作为该主成分的显著变量。截取累积方差(Cumulative variance)超过总方差60%的主成分。按照主成分象限排列组合的结果,计算出所有可能的生境类型,并对2 个公园的样方进行生境分类。使用卡方适合性检验,比较2 个公园中不同生境中鸟种数(RICb)、鸟类数量(ABUb)和每种生境平均鸟类数量(AABU)是否存在显著差异。
1.5.2 共有陆生鸟种的生境类型利用差异比较
统计2 个公园中共有陆生鸟种对可利用生境类型的利用比例(某一鸟种利用的生境类型数/该公园生境类型总数)。经单样本Kolmogorov-Smirnov 检验确认数据符合正态分布,因此使用配对样本t检验比较共有鸟种在2 个公园中生境类型的使用差异。
1.5.3 共有生境类型中陆生鸟种组成的差异比较
统计2 个公园共有生境类型中出现的陆生鸟类种数及其科属分类。数据经对数转换后符合正态分布,因此采用配对样本t检验,分析2 个公园共有生境类型中鸟种数(RICb)差异的著差性。
2 结果与分析2.1 公园鸟类物种组成
本次调查共记录到鸟类58 种4312 次/只,分属10 目23 科。其中,世纪公园共记录到鸟类7 目20 科40 种2618 次/只,滨江森林公园共记录到鸟类8 目20 科47 种1694 次/只,两个公园共有鸟种24 种,其中陆生鸟类21 种,分属2 目12 科,共有陆生鸟类分别占世纪公园鸟类数量的82.6%(2163/2618),滨江森林公园鸟类数量的83.3%(1411/1694)。
2.2 公园生境类型组成
PCA结果显示,前4个主成分(PC1—PC4)的特征值均大于1,累积方差占总方差的77.2%。第1主成分(PC1)的显著变量分别为COVs,DIVs,RICs和EVEs,主要反映了灌木层的盖度和多样性信息。第2主成分(PC2)的显著变量为ABUt,主要反应乔木层的乔木数量。第3主成分(PC3)的显著变量为DIVt和EVEt,反应乔木层的多样性信息。第4主成分(PC4)的显著变量为RICg和ABUg,反应地被层的丰富度和多度信息(表1)。由于所有载荷量绝对值≥0.7的变量均为正值,因此只有样方主成分得分为正时,表示样方包含该主成分信息。根据样方在PC1—PC4上的得分情况绘制图1和图2,PC1正方向代表包含发达灌木层的生境(S),PC2正方向和PC3正方向均代表包含发达乔木层的生境(T),PC4正方向代表包含发达地被层的生境(G)。将2 个公园的样方按照上述象限组合标准进行排列组合,40 个样方可以划分为8 种生境类型,包括乔木层植物发达的生境(T型)、灌木层植物发达的生境(S型)、地被层植物发达的生境(G型)、乔木层和灌木层植物都发达的生境(T+S型)、乔木层和地被层植物都发达的生境(T+G型)、灌木层和地被层植物都发达生境(S+G型)、植被分层结构复杂生境(T+S+G型)和人工修饰生境(D型)(表2)。生境类型名称,数目及植被特征描述见表2。卡方检验结果表明,2 个公园不同类型生境数量存在显著差异(X2=16.98, df=7,P=0.018)。世纪公园缺乏S型生境,可利用的生境有7 种,T+G型生境最丰富。而滨江森林公园缺乏G型和T+G型生境,可利用的生境有6 种,T+S型生境最丰富(表2)。2 个公园共有的生境类型有5 种,分别为T型,T+S型,S+G型,T+S+G型和D型。

表1 植被变量的主成分分析结果Table 1 Principal component analysis of the variables that responded to the vegetation composition
2.3 生境类型和鸟类群落结构的关系
卡方适合性检验结果表明,世纪公园鸟种数(RICb,X2=23.28, df=6,P<0.001),鸟类数量(ABUb,X2=1567.23, df=6,P< 0.001)和每种生境类型的平均鸟类数量(AABU,X2=79.65, df=6,P< 0.001)在不同类型生境的分布均存在显著差异。世纪公园T+G型生境RICb最高,T+S型生境RICb最低;T+G型生境ABUb最高,S+G型生境ABUb最低;D型生境AABU最高,S+G型生境AABU最低。滨江森林公园ABUb(X2=543.07, df=5,P< 0.001)和AABU(X2=25.47, df=5,P< 0.001)在不同生境的分布均存在显著差异,而RICb在不同生境的分布没有显著差异(X2=7.547, df=5,P= 0.183)。滨江森林公园T+S型生境的ABUb最高,S+G型生境的ABUb最低;S型生境AABU最高,D型生境AABU最低;RICb在各种生境的分布无显著差异。因此,滨江森林公园鸟类群落对生境的利用更加均匀,而世纪公园鸟类群落对生境的利用选择性较强。
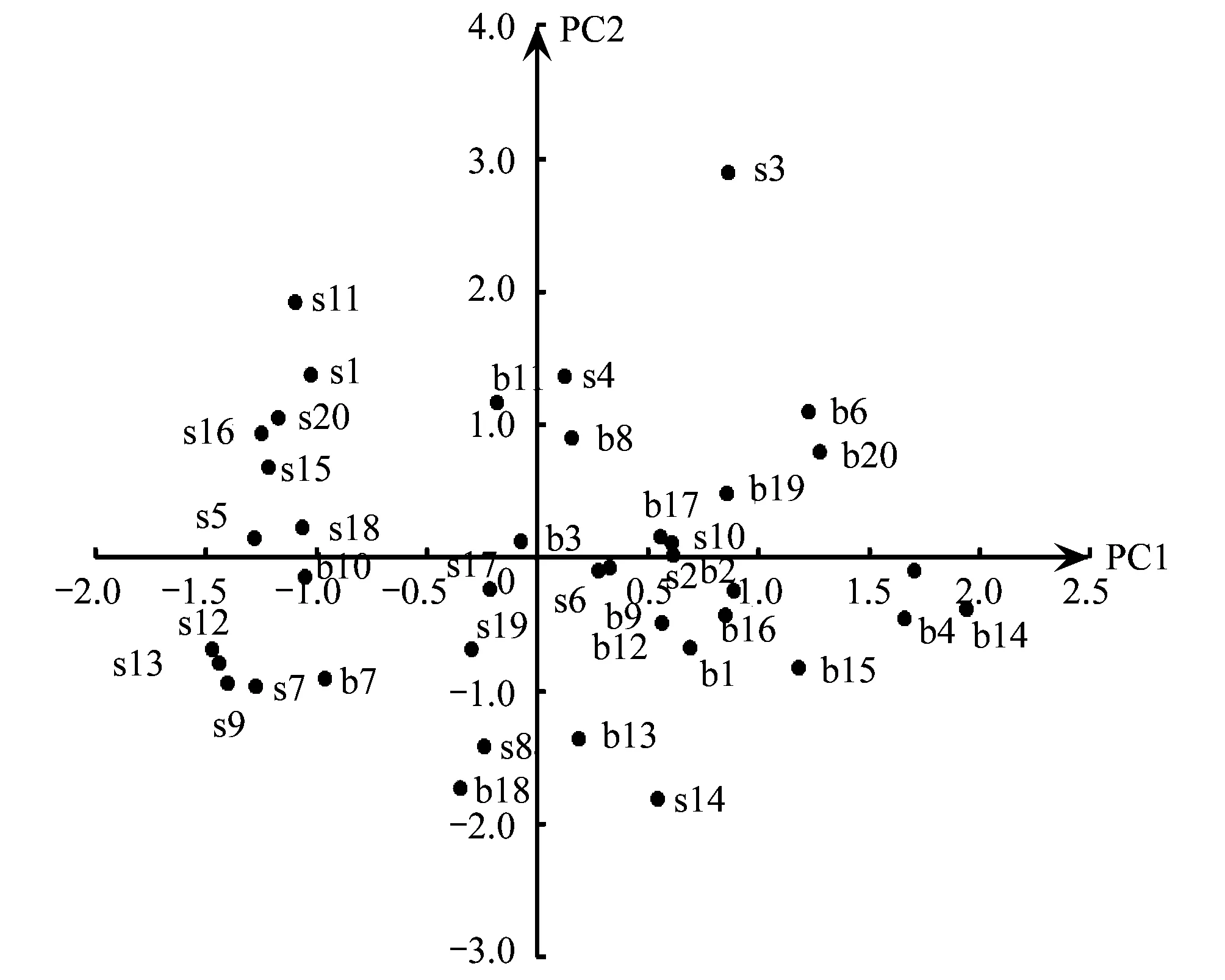
图1 公园样方在PCA(Principal Component Analysis)第1轴(PC1)和第2轴(PC2)上的排序
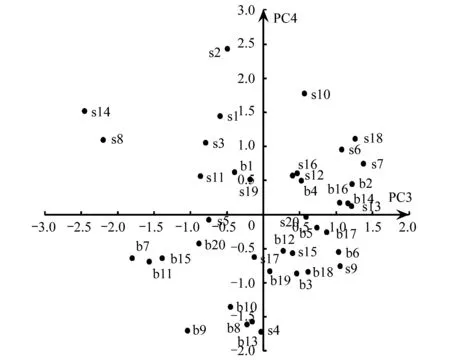
图2 公园样方在PCA第3轴(PC3)和第4轴(PC4)上的排序
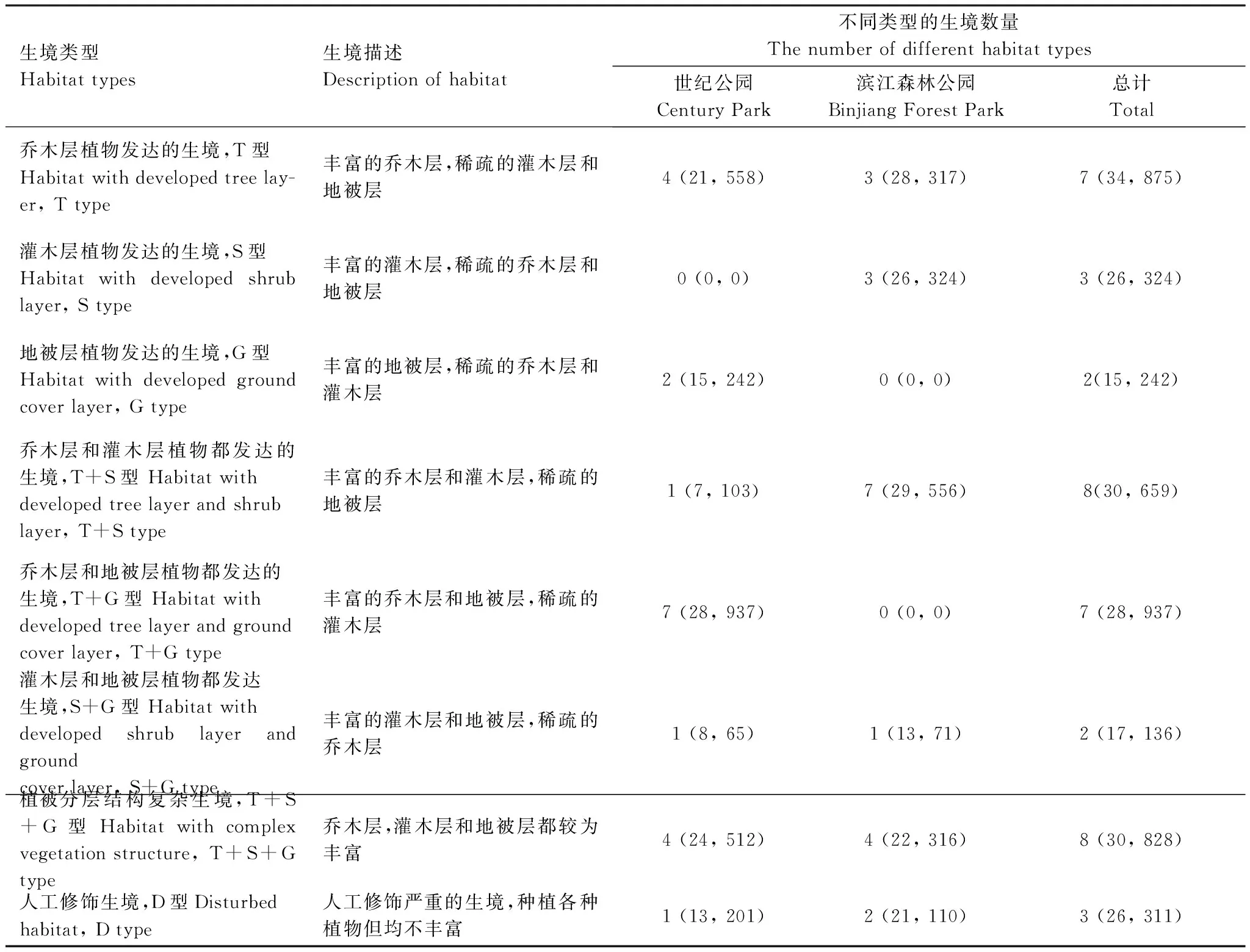
表2 基于主成分分析结果的公园生境分类Table 2 Classification of habitat types in urban parks based on Principal Component Analysis
2.4 2个公园共有陆生鸟种的生境类型利用差异比较
为了比较2 个公园共有鸟种的生境利用差异,本研究统计了2 个公园共有鸟种对可选择生境的利用比例(某一鸟种选择的生境类型数/该公园可选择的生境总数)。该比例进行对数转换后符合正态分布,进行配对t检验,结果表明共有鸟种对滨江森林公园的利用比例显著高于世纪公园(t=2.247, df=23,P=0.035, 表3)。
表3 两个公园24种共有鸟种对各生境类型的利用比例(某一鸟种利用的生境类型数/该公园可利用的生境总数)比较
Table 3 The comparison of the utilization ratio of the available habitat by the species recorded in both parks(the habitat types actually used/ the available habitat types in the park)

共有鸟种Commonbirdspecies共有鸟种对各生境类型的利用比例Theutilizationratiooftheavailablehabitatbythespeciesrecordedinbothparks世纪公园CenturyPark滨江森林公园BinjiangForestPark白腹鸫Turduspallidus4/75/6白鹡鸰Motacillaalba7/76/6白鹭Egrettagarzetta*2/72/6白头鹎Pycnonotussinensis7/76/6斑鸫Turdusnaumanni2/76/6大山雀Parusmajor3/76/6戴菊Regulusregulus1/72/6黑尾蜡嘴雀Eophonamigratoria5/76/6红胁蓝尾鸲Tarsigercyanurus2/74/6虎斑地鸫Zootheradauma2/73/6黄腹山雀Parusvenustulus2/71/6黄喉鹀Emberizaelegans2/71/6黄腰柳莺Phylloscopusproregulus1/71/6灰背鸫Turdushortulorum2/73/6灰头鹀Emberizaspodocephala2/73/6灰喜鹊Cyanopicacyana7/74/6山斑鸠Streptopeliaorientalis2/75/6树鹨Anthushodgsoni2/72/6乌鸫Turdusmerula7/76/6小Tachybapusruficollis*5/72/6夜鹭Nycticoraxnycticorax*5/74/6珠颈斑鸠Streptopeliachinensis7/76/6棕背伯劳Laniusschach5/76/6棕头鸦雀Paradoxorniswebbianus4/73/6
*表示在陆生生境中记录到的水鸟,这些鸟种不计入统计分析
2.5 2 个公园共有生境类型中陆生鸟种组成的差异比较
对2 个公园共有生境类型的鸟类物种数进行对数转换后符合正态分布,利用配对t检验,发现2 个公园共有生境类型的鸟种数之间无显著差异(t=2.045,df=4,P= 0.110)。但2 个公园共有生境类型中鹟科(Muscicapidae)鸟种数(RICb)存在显著差异(t=2.804,df=4,P= 0.049),滨江森林公园共有生境中鹟科鸟类RICb显著多于世纪公园。在世纪公园中鹟科鸟类仅出现在T+S+G型生境,而在滨江森林公园中则出现在T型、T+S型、T+S+G型和D型生境中。其余各科鸟类RICb之间并无显著差异。
3 讨论
3.1 公园生境结构对鸟类群落的影响
由于2 个公园在构建景观过程中均重视乔木植物的保护与种植[21- 23],因此两个公园发达乔木层生境都很丰富。世纪公园T+G型生境最为丰富(7/20),T型(4/20)和T+S+G型(4/20)生境也较为丰富。这3 种生境中记录到鸟种数和鸟类数量占世纪公园总数的90.0%(36/40)和76.7%(2007/2618)。滨江森林公园T+S型生境最为丰富(7/20),T+S+G型(4/20)和T型(3/20)生境也较为丰富。这3 种生境中记录到鸟种数和鸟类数量占滨江森林公园总数的87.2%(41/47)和70.2%(1189/1694)(表2)。因此,多数鸟类选择包含发达乔木层的生境,这一点与Evans等[24]的研究是一致的。另外,乔木层是最重要的植被结构之一,由于其为鸟类提供了栖息、筑巢、隐蔽和觅食的场所[25- 26],因此,在城市绿地中乔木层能显著增加鸟类丰富度和鸟类多样性水平[27- 28]。
2 个公园的生境类型组成存在显著差异。差异之一是世纪公园发达灌木层生境较少。世纪公园包含发达灌木层的生境有T+S型(1/20)、S+G型(1/20)和T+S+G型(4/20),这3 种生境中记录到的鸟种数和鸟类数量占世纪公园总数的62.5%(25/40)和26.0%(680/2618)。而滨江森林公园包含发达灌木层的生境有S型(3/20)、T+S型(7/20)、S+G型(1/20)和T+S+G型(4/20),这4 种生境中记录到鸟种数和鸟类数量占滨江森林公园总数的76.6%(36/47)和74.8%(1267/1694)(表2)。滨江森林公园比世纪公园拥有更加丰富的灌木栖息地,因此在滨江森林公园灌木生境中鸟类资源更加丰富,并且存在仅在此种生境中出现的鸟种。例如,乌灰鸫(Turduscardis)仅在滨江森林公园S型生境中发现,北灰鹟(Muscicapadauurica)和田鹀(Emberizarustica)仅滨江森林公园在T+S型生境中发现。Melles[13]和Hedblom[29]的研究发现灌木丰富的生境对鸟类丰富度和多度,尤其是森林鸟类的多度具有显著影响。同时灌木层盖度和多样性水平也会显著提高鸟类多样性水平[30]。
2 个公园的生境类型组成存在的另一差异在于滨江森林公园发达地被层生境较少。世纪公园包含发达地被层的生境有G型(2/20) 、T+G型(7/20)、S+G型(1/20)和T+S+G型(4/20),这4 种生境中记录到的鸟种数和鸟类数量占世纪公园总数的92.5%(37/40)和67.1%(1756/2618)。而滨江森林公园包含发达地被层的生境有S+G型(1/20)和T+S+G型(4/20),这2 种生境中记录到鸟种数和鸟类数量占滨江森林公园总数的51.1%(24/47)和22.8%(387/1694)(表2)。世纪公园比滨江森林公园拥有更加丰富的地被层栖息地,因此世纪公园地被层生境中鸟类资源更加丰富。地被层为部分鸟类提供食物来源[13],在本研究中,灰椋鸟(Sturnuscineraceus)和丝光椋鸟(S.sericeus)等仅在世纪公园中发现的鸟类其觅食行为主要发生在地被层,关于植被垂直结构对鸟类群落的影响另文发表。
3.2 鸟类在公园中的生境利用率的差异
鸟种数在世纪公园各类生境中的分布存在显著差异,而在滨江森林公园中无显著差异。同时,滨江森林公园共有鸟种对可利用生境的利用比例显著高于世纪公园(表3)。世纪公园鸟类的分布比滨江森林公园更加集中。世纪公园90.0% (36/40) 的鸟种数和61.8% (1617/2618) 的鸟类数量分布在复杂生境中(由2 个及其以上植被层次组成的生境),而滨江森林公园这一比例只有68.1% (32/47)和55.7% (943/1694)。通过对2 个公园木本植物多样性的比较,滨江森林公园乔木多样性和灌木多样性均显著大于世纪公园(F=13.901,P<0.001;F=7.390,P=0.002)。滨江森林公园比世纪公园木本植物多样性更高,因此生境垂直结构更加丰富。这就使得滨江森林公园的鸟类分布更加分散。Roth[31]的研究也证明了植被多样性与鸟类均匀度呈显著的正相关。世纪公园鹟科鸟类仅在T+S+G型生境中出现,而在滨江森林公园T型、T+S型、T+S+G型和D型生境中均有所记录,结合2 个公园木本植物多样性的差异,鹟科鸟类倾向于选择更加植被多样性更高且植被分层结构复杂的生境。
另一方面,城市化水平对鸟类的生境选择也具有显著影响[32],城市化水平较高的区域鸟类对生境的选择更加单一,稀有鸟种减少,伴人种增加并且生物量增加[14]。世纪公园比滨江森林公园更靠近市中心,因此鸟种数少于滨江森林公园而鸟类数量多于滨江森林公园。
3.3 大型城市公园建设的现状及建议
城市景观公园中往往具有多种景观类型,不同景观的密集排列和组合,产生了丰富多样的边界[33],因此生境类型丰富[34]。因此,通过合理配置植被使多种景观类型所产生丰富的边缘效应,以此来提高公园鸟类的多样性水平是城市公园生态服务功能的重要体现。由于乔木层是最重要的植被结构之一,为多数鸟类提供了栖息、运动和觅食的场所,因此应多样化种植各类乔木[35],如针叶林和阔叶林相以及不同果期乔木结合,具体树种搭配参考王勇等[36]。由于灌木层生境较少,世纪公园缺少了一些仅在灌木层生境生活的鸟类,因此,从增加鸟类多样性角度,建议公园在乔木层林下多样化搭配灌木树种。由于地被层是鸟类食物来源地之一,建议在保留供游客休憩草坪区域的同时多样化种植草本植物,同时通过草坪分区产生更加丰富的边界以提高鸟类多样性[37- 38]。
致谢:调查工作得到上海世纪公园和滨江森林公园全体工作人员的帮助, 华东师范大学何娅博士、杨会涛博士、刘旭博士、毕玉科、周青梅、陈珏、杨应远和顾孙龙对论文写作给予帮助,特此致谢。
[1] Grimm N B, Faeth S H, Golubiewski N E, Redman C L, Wu J G, Bai X M, Briggs J M. Global change and the ecology of cities. Science, 2008, 319(5864): 756- 760.
[2] 牛文元. 中国新型城市化报告2012. 北京: 科学出版社, 2012: 103- 110.
[3] 赵斌, 唐礼俊, 吴千红, 陈家宽. 上海市生物多样性信息管理系统的建立和应用. 生物多样性, 2000, 8(2): 233- 237.
[4] Roberge J M, Angelstam P. Indicator species among resident forest birds-A cross-regional evaluation in northern Europe. Biological Conservation, 2006, 130(1): 134- 147.
[5] Rotenberry J T, Wiens J A. Foraging patch selection by shrubsteppe sparrows. Ecology, 1998, 79(4): 1160- 1173.
[6] 栾晓峰. 上海鸟类群落特征及其保护规划研究 [D]. 上海: 华东师范大学, 2003.
[7] Sandström U G, Angelstam P, Mikusiński G. Ecological diversity of birds in relation to the structure of urban green space. Landscape and Urban Planning, 2006, 77(1/2): 39- 53.
[8] McKinney M L. Effects of urbanization on species richness: a review of plants and animals. Urban Ecosystems, 2008, 11(2): 161- 176.
[9] 陈水华, 丁平, 郑光美, 王玉军. 园林鸟类群落的岛屿性格局. 生态学报, 2005, 25(4): 657- 663.
[10] Wang Y P, Ding P, Chen S H, Zheng G M. Nestedness of bird assemblages on urban woodlots: Implications for conservation. Landscape and Urban Planning, 2013, 111: 59- 67.
[11] Díaz L. Influences of forest type and forest structure on bird communities in oak and pine woodlands in Spain. Forest Ecology and Management, 2006, 223(1/3): 54- 65.
[12] Heyman E. Clearance of understory in urban woodlands: Assessing impact on bird abundance and diversity. Forest Ecology and Management, 2010, 260(1): 125- 131.
[13] Melles S, Glenn S, Martin K. Urban bird diversity and landscape complexity: Species-environment associations along a multiscale habitat gradient. Conservation Ecology, 2003, 7(1): 5- 5.
[14] Fontana S, Sattler T, Bontadina F, Moretti M. How to manage the urban green to improve bird diversity and community structure. Landscape and Urban Planning, 2011, 101(3): 278- 285.
[15] 陈水华, 丁平, 范忠勇, 郑光美. 城市鸟类对斑块状园林栖息地的选择性. 动物学研究, 2002, 23(1): 31- 38.
[16] 蔡音亭, 唐仕敏, 袁晓, 王吉衣, 马志军. 上海市鸟类记录及变化. 复旦学报: 自然科学版, 2011, 50(3): 334- 343.
[17] 上海市统计局. 上海统计年鉴(2012). (2013-01- 19) [2013-05- 24]. http://www.stats-sh.gov.cn/tjnj/tjnj2012.htm.
[18] Bachin R F. Cultivating unity: The changing role of parks in urban America. Places-a Forum of Design for the Public Realm, 2003, 15(3): 12- 17.
[19] Volpato G H, Lopes E V, Mendonca L B, Bocon R, Bisheimer M V, Serafini P P, Anjos L. The use of the point count method for bird survey in the Atlantic forest. Zoologia, 2009, 26(1): 74- 78.
[20] Shwartz A, Shirley S, Kark S. How do habitat variability and management regime shape the spatial heterogeneity of birds within a large Mediterranean urban park? Landscape and Urban Planning, 2008, 84(3/4): 219- 229.
[21] 任梦非, 朱祥明. 上海滨江森林公园规划设计研究. 中国园林, 2007, (2): 21- 27.
[22] 万福绪, 董波, 陈敏. 上海滨江森林公园景观空间格局分析. 中国园林, 2008, (7): 61- 65.
[23] 王震威. 上海世纪公园生态景观功能提升的实践析要 [D]. 上海: 上海交通大学, 2009.
[24] Evans K L, Newson S E, Gaston K J. Habitat influences on urban avian assemblages. Ibis, 2009, 151(1): 19- 39.
[25] Mills G S, Dunning J B Jr, Bates J M. Effects of urbanization on breeding bird community structure in southwestern USA desert habitats. Condor, 1989, 91(2): 416- 428.
[26] Munyenyembe F, Harris J, Hone J, Nix H. Determinants of bird populations in an urban area. Australian Journal of Ecology, 1989, 14(4): 549- 557.
[27] Clergeau P, Savard J P L, Mennechez G, Falardeau G. Bird abundance and diversity along an urban-rural gradient: A comparative study between two cities on different continents. Condor, 1998, 100(3): 413- 425.
[28] Palomino D, Carrascal L M. Urban influence on birds at a regional scale: A case study with the avifauna of northern Madrid province. Landscape and Urban Planning, 2006, 77(3): 276- 290.
[29] Hedblom M, Söderström B. Landscape effects on birds in urban woodlands: an analysis of 34 Swedish cities. Journal of Biogeography, 2010, 37(7): 1302- 1316.
[30] Leston L F V, Rodewald A D. Are urban forests ecological traps for understory birds? An examination using Northern cardinals. Biological Conservation, 2006, 131(4): 566- 574.
[31] Roth R R. Spatial heterogeneity and bird species diversity. Ecology, 1976, 57(4): 773- 782.
[32] Blair R B, Johnson E M. Suburban habitats and their role for birds in the urban-rural habitat network: points of local invasion and extinction? Landscape Ecology, 2008, 23(10): 1157- 1169.
[33] Pellissier V, Cohen M, Boulay A, Clergeau P. Birds are also sensitive to landscape composition and configuration within the city centre. Landscape and Urban Planning, 2012, 104(2): 181- 188.
[35] 陈水华, 丁平, 郑光美, 诸葛阳. 岛屿栖息地鸟类群落的丰富度及其影响因子. 生态学报, 2002, 22(2): 141- 149.
[36] 王勇, 许洁, 杨刚, 李宏庆, 吴时英, 唐海明, 马波, 王正寰. 城市公共绿地常见木本植物组成对鸟类群落的影响. 生物多样性, 2014, 22(2): 196- 207.
[37] Carbó-Ramírez P, Zuria I. The value of small urban greenspaces for birds in a Mexican city. Landscape and Urban Planning, 2011, 100(3): 213- 222.
[38] Strohbach M W, Haase D, Kabisch N. Birds and the city: urban biodiversity, land use, and socioeconomics. Ecology and Society, 2009, 14(2): 31- 31.
The influence of habitat types on bird community in urban parks
YANG Gang1,2, WANG Yong1,2,XU Jie1,2, DING Youzhong1,2, WU Shiying3, TANG Haiming3, LI Hongqing1, WANG Xiaoming4, MA Bo5, WANG Zhenghuan1,2,*
1SchoolofLifeSciences,EastChinaNormalUniversity,Shanghai200062,China2ShanghaiKeyLaboratoryofUrbanizationandEcologicalRestoration,EastChinaNormalUniversity,Shanghai200241,China3ForestryStationofPuDongNewDistrict,Shanghai201210,China4ShanghaiScienceandTechnologyMuseum,Shanghai200127,China5DepartmentofWildlifeProtectionandManagementAdministration,Shanghai200023,China
The bird communities and vegetation communities in Shanghai Century Park and Bingjiang Forest Park were respectively surveyed from December 2011 to November 2012. In accordance with the result of principal components analysis of 12 vegetation variables, the vegetation in the two parks was grouped into 8 types including T type (habitat with developed tree layer), S type (habitat with developed shrub layer), G type (habitat with developed ground cover layer), T+S type (habitat with developed tree layer and shrub layer), T+G type (habitat with developed tree layer and ground cover layer), S+G type (habitat with developed shrub layer and ground cover layer), T+S+G type (habitat with complex vegetation structure), D type (disturbed habitat). Chi-square test suggested that the S type and T+S type habitat in Binjiang Forest Park was significantly more abundant than those in Century Park while G type and T+G type habitat in Century Park was significantly more abundant than those in Binjiang Forest Park. Developed tree layer habitat was abundant both in the two parks and More than 70% of the bird richness and bird abundance were recorded in these habitat. Bird richness was significantly different in habitat types in Century Park, but the richness difference was not significant in Binjiang Forest Park. There were 24 common bird species in the two parks, which were indicated to have performed higher utility rate in Binjiang Park than those in Century Park in accordance with paired sample t test result. Analysis of the bird richness in common habitat types of the two parks showed that the richness of Muscicapidae in Binjiang Park was significantly higher than that of their counterparts in Century Park. Due to the forgoing, we concluded that (1) trees were one of the most important vegetation components that enhanced the bird species richness and diversity in urban green spaces; (2) shrubs were usually treated as being as important as trees for birds living in urban areas, where they provided some forest birds with nesting sites; (3) ground cover layer provided the ground foraging groups with feeding resources. Our research emphasized that habitat types significantly influenced the structure of bird communities in the urban parks in Shanghai. Therefore, we suggest that hierarchical vegetation types should be adopted while planning and constructing large-scale urban parks so that the bird diversity will be improved. Furthermore, in order to perform higher utility rate of urban birds, diversified shrubs and different kinds of herbaceous plant should be planted under the tree layer.
urban parks; habitat types; bird community; vegetation structure; Shanghai
上海市绿化和市容管理局专项基金、浦东新区科技发展基金(PKJ2011- N020)
2013- 09- 11;
2014- 07- 02
10.5846/stxb201309112250
*通讯作者Corresponding author.E-mail: zhwang@bio.ecnu.edu.cn
杨刚,王勇,许洁, 丁由中,吴时英,唐海明, 李宏庆,王小明, 马波,王正寰.城市公园生境类型对鸟类群落的影响.生态学报,2015,35(12):4186- 4195.
Yang G, Wang Y,Xu J, Ding Y Z, Wu S Y, Tang H M, Li H Q, Wang X M, Ma B, Wang Z H.The influence of habitat types on bird community in urban parks.Acta Ecologica Sinica,2015,35(12):4186- 4195.
猜你喜欢
现代装饰(2022年6期)2022-12-17
湖南大学学报(自然科学版)(2022年8期)2022-09-02
大众摄影(2021年3期)2021-03-22
江淮法治(2020年9期)2020-07-11
故事作文·低年级(2019年8期)2019-08-20
乡村地理(2018年1期)2018-07-06
现代园艺(2018年3期)2018-02-10
浙江林业(2016年5期)2016-06-14
十几岁(2016年1期)2016-04-09
杭州(2015年9期)2015-12-21
