
鼎湖山针阔叶混交林4种优势树种树干液流特征
2015-02-06 03:54欧阳旭黄德卫刘世忠张德强李跃林
生态学报 2015年12期
程 静, 欧阳旭, 黄德卫, 刘世忠, 张德强, 李跃林,*
1 中国科学院华南植物园, 广州 510650 2 中国科学院大学, 北京 100049 3 中国林业科学研究院热带林业实验中心, 凭祥 532600
鼎湖山针阔叶混交林4种优势树种树干液流特征
程 静1,2, 欧阳旭1,2, 黄德卫1,3, 刘世忠1, 张德强1, 李跃林1,*
1 中国科学院华南植物园, 广州 510650 2 中国科学院大学, 北京 100049 3 中国林业科学研究院热带林业实验中心, 凭祥 532600
运用Granier热消散式探针法,对鼎湖山自然保护区针阔叶混交林4种优势树种(马尾松、木荷、锥栗和广东润楠)的树干液流密度进行了长期连续观测,并同步监测林分的环境因子(光合有效辐射(PAR)、空气温度(T)、空气湿度(RH)、土壤含水量)。结果表明:(1)4种优势树种的边材面积(As)与胸径(DBH)均存在显著相关关系(P<0.05);(2)各优势树种树干液流均呈现“昼高夜低”单峰曲线,且液流速率存在明显的季节性差异;(3)无论湿季还是干季,光合有效辐射(PAR)和水汽压亏缺(VPD) 均为控制蒸腾的主要驱动因子;(4) 4种优势树种湿季平均日蒸腾量高于干季,马尾松、木荷、锥栗、广东润楠湿季平均日蒸腾量分别为29.52、39.29、30.40、9.41 kgH2O/d,干季分别为20.91、24.84、24.26、8.43 kgH2O/d,干季和湿季的平均日蒸腾量(kgH2O/d)大小均为木荷>锥栗>马尾松>广东润楠,这种种间差异是由边材面积大小和树种本身的生物学特性共同决定的。
Granier热消散式探针; 针阔叶混交林; 边材面积; 树干液流; 日蒸腾
树干液流(sap flow)是指在叶片蒸腾拉力作用下,植物体木质部内的水分自根部运输到叶片中的过程,在树干上升的液流中,有99.8% 以上消耗于蒸腾[1],一般用木质部边材液流量表征树木的蒸腾耗水量[2]。因此,树干木质部液流速度及液流量的大小制约着整株树的蒸腾量[3]。目前国际上广泛运用的树干液流测定法主要有三类:组织热平衡法、热脉冲法、热消散式探针法[4]。其中Granier热消散式探针法由于具有保持树木在自然条件下,基本不破坏树木正常生长状态,可以连续测定树干液流量等优点,被认为是目前测定乔木蒸腾作用最准确的方法[5- 6]。
目前我国应用Granier热消散式探针法对树干液流的研究较多[7- 10],但大量的研究主要集中于单一树种的人工林[7- 8],少有的自然林研究主要集中于液流与环境因子关系的探讨[10- 12],对自然林蒸腾的研究甚少[12]。在我国南亚热带鼎湖山自然保护区,黄德卫等[10]已对该区海拔约50 m的典型地带性植被常绿针阔混交林进行相关研究,揭示其优势树种马尾松(Pinusmanssoniana)、木荷(Schimasuperba)、锥栗(Castanopsischinensis)、广东润楠(Machiluskwangtungensis) 树干液流的基本规律及其与环境因子之间的关系。然而不同海拔高度引起的环境因子差异必然导致林分水分利用效率产生变化,因此本研究期望在前期研究基础上进一步揭示不同海拔的针阔叶混交林优势树种的树干液流特征,为估算整个区域针阔叶混交林森林生态系统的蒸腾量奠定基础,为深入研究自然林水分平衡和林地水分利用提供理论和方法依据。
1 实验材料与方法
1.1 样地与样树概况
鼎湖山国家级自然保护区(112°30′39″—112°33′41″E,23°09′21″—23°11′30″N)位于广东省肇庆市境内,总面积1155 hm2,属低山丘陵地貌,最高峰鸡笼山海拔1000.3 m。本区属典型南亚热带湿润季风气候,年平均气温21 ℃,最热月(7月)和最冷月(1月)平均气温分别为28.0 ℃、12.0 ℃,此区年均降雨量1956 mm,年平均相对湿度为81.5%,全年干湿季明显,降雨量的76%发生在湿季的4—9月[13- 15]。
鼎湖山主要植被类型为季风常绿阔叶林、针阔叶混交林和马尾松林。本文研究的针阔混交林位于保护区内五棵松,林地海拔320 m左右,坡向东偏南,坡度约10°,由人工种植或自然生长的马尾松群落被先锋阔叶树种入侵后自然发展演变而成,是马尾松林向季风常绿阔叶林演替发展的一个过渡类型。林下灌木主要有山血丹(Ardisialindleyana)、九节(Psychotriarubra)、锈叶新木姜子(Neolitseacambodiana)等;林下草本植物以芒萁(Dicranopterispedata)、双盖蕨(Diplaziumdonianum)为主。在针阔叶混交林选取30 m×40 m样地,根据样地实际情况,选取树干圆满通直、生长状况良好且无病虫害的主要优势树种马尾松、锥栗、广东润楠各3棵、木荷5棵为测定对象,样树树形基本特征如表1。

表1 样树基本特征Table 1 Basic characteristics of the sampling trees
1.2 树干液流密度的测定
综合气象资料,采用Granier热扩散探针法于2010年7月(湿季)和11月(干季)[10]对14株样树液流密度进行连续测定。分别将每组探针上下相距10—15 cm安装于树干北面(避免阳光直射)[10]胸高1.3 m处。为防止雨水接触探针,导致测定产生误差,在探针外覆盖泡沫盒,并包裹防辐射薄膜,减小温度波动。上探针供以12 V直流电压持续加热(0.2 W),下探针作为参照不加热。两探针之间的温差电势应用数据采集器DL2e(Delta-T Devices, England)自动记录和存储(每10 s测读1次数据,存储每30 min的平均值)。根据Granier建立的经验公式将温差电势转化为液流密度[16]:
Js= 119 × [(ΔTm-ΔT) /ΔT]1.231
(1)
式中,ΔTm为上、下探针之间的最大昼夜温差,ΔT为瞬时温差,Js为瞬时液流密度(g H2O m- 2s-1) 转换为液流密度值。该公式是Ganier经过多年在多种树木进行研究总结出的经验公式,适用于任何树种[17]。
1.3 边材面积的确定
在实际操作中,只能通过测定边材厚度求算边材面积。出于对自然保护区植被保护的原则,并避免对样树造成伤害,因此对研究地区采取抽样调查的办法(不包括待测液流的样树),每种树种选取6—8棵,量取胸径,采用直径5 mm生长锥于树干北面胸高1.3 m处钻取木栓,考虑到林内郁闭度大,树干圆满通直,主要以北面木栓为主,量取胸高处边材厚度,计算边材面积并建立边材面积与胸径的关系式[7]:
As=m(DBH)n
(2)
式中,As代表边材面积,DBH为胸径,m、n为参数。
1.4 环境因子的测定
微型气象观测仪安装于样地内高约10 m处的空旷地铁架上,以避免树木等障碍物影响数据准确性。无线电子测量记录器(Der elektronische Funk-Messlogger funky_Clima,德国)用于测量气温(T)和空气相对湿度(RH)。Li-cor光合有效辐射传感器(LI-COR Environmental,美国)用于对光合有效辐射进行连续监测。土壤10 cm和30 cm处安插Em50-ECH2O型土壤含水量传感器,环境因子数据采集和存储频率均与树干液流相同。为综合表达温度和空气相对湿度的协同效应采用VPD(vapor pressure deficiency)这一指标,采用下式计算VPD[18]:
es(T) =a×exp[bT/(T+c)]
(3)
VPD =es(T)-ea=es(T) (1-RH)
(4)
式中,es(T)代表T温度下的饱和水汽压(kPa),a、b、c为参数,分别取值为:0.611 kPa、 17.502、 240.97 ℃,T为温度(℃),ea为实际水汽压(kPa),RH为相对湿度,VPD为叶片和空气之间的水汽压亏缺(kPa)。
1.5 数据处理
本文运用Excel2003软件进行数据处理,用SPSS17.0统计软件对优势树种树干液流进行偏相关和回归分析,并运用Sigmaplot10.0绘图。
2 结果与分析
2.1 树种边材面积与胸径的关系
边材作为水分的输导组织,其面积大小与水分的传导能力直接相关[19]。边材面积是计算液流和整树蒸腾的关键参数。图1为根据各优势树种胸径与边材面积相关关系拟合回归方程而绘制的散点图。结果表明,胸径和边材之间存在着高度相关的幂函数关系,4优势树种马尾松、木荷、锥栗、广东润楠的判定系数R2值分别为:0.99、0.96、0.93、0.81,由此可见,各公式均能非常好地拟合胸径与边材面积的关系。
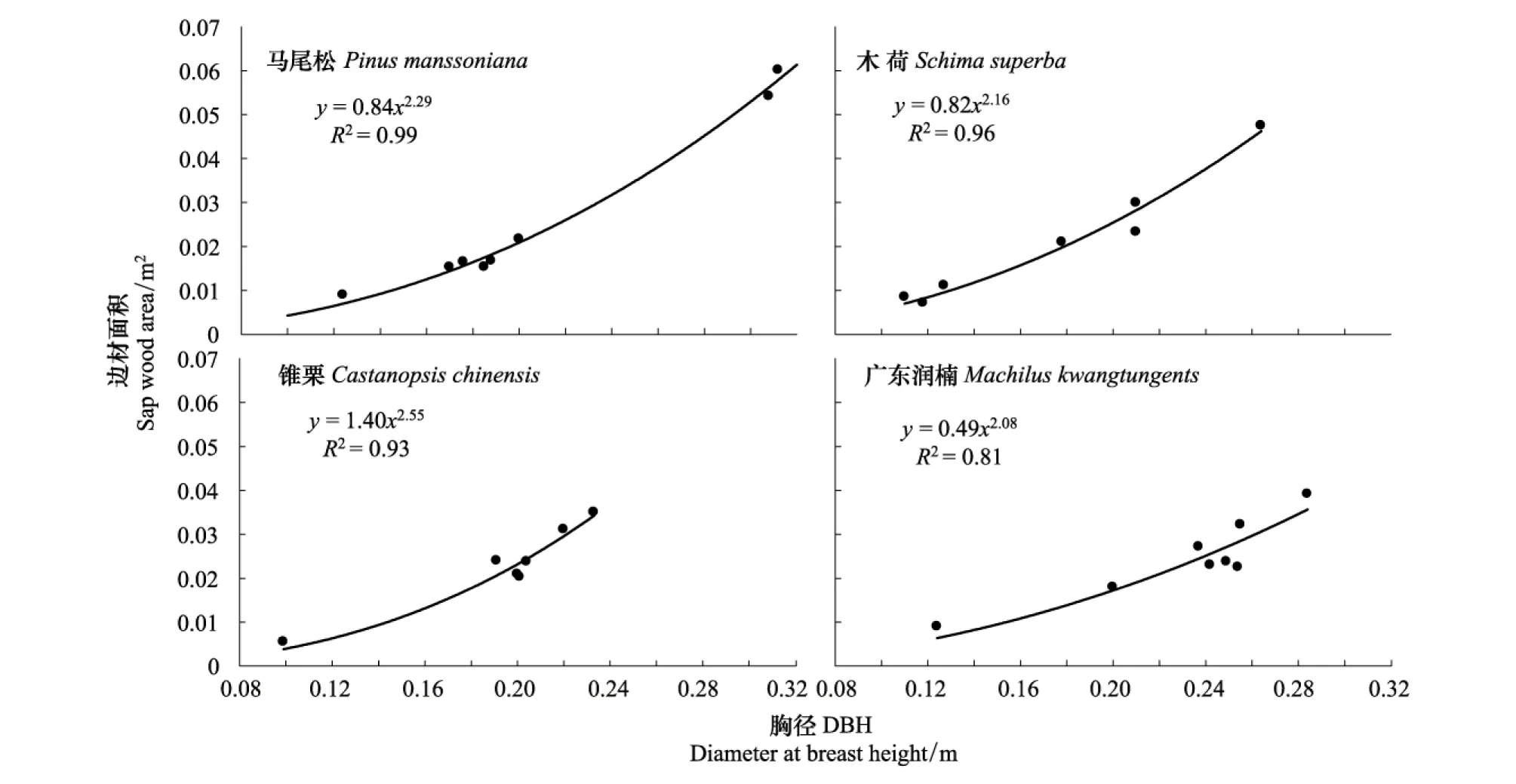
图1 优势树种边材面积与胸径关系
2.2 不同优势树种液流日变化
分别将同一树种湿季(7月)和干季 (11月)各单日液流速率数据进行每30 min平均,如图2所示,无论是湿季(7月)还是干季(11月),马尾松、木荷、锥栗、广东润楠的树干液流均呈现典型的 “昼高夜低”单峰曲线。湿季(7月)液流速率呈现明显“宽峰”型曲线。干季(11月)高峰时间段较短,整体趋于“尖峰”型曲线。各优势树种液流日变化速率存在明显的季节性差异,主要表现在液流启动时间、到达峰值时间、峰值区间差异以及液流下降速率。湿季(7月)液流在6:30—7:00启动,马尾松与广东润楠液流启动时间滞后木荷和锥栗约30 min。干季(11月)液流启动时间为7:30—8:00,锥栗液流启动约在7:30,马尾松与木荷滞后锥栗约30 min。随着太阳辐射的增强,气温逐渐升高,空气相对湿度降低,冠层气孔导度不断升高,液流速率逐渐增加。湿季(7月)峰值出现在14:00—14:30之间,滞后干季(11月)约30—120 min。7月平均液流峰值大小依次为锥栗(30.76 gH2O m-2s-1)>木荷(27.51 g H2O m-2s-1)>马尾松(26.66 gH2O m-2s-1)>广东润楠(23.69 gH2O m-2s-1)。11月平均液流峰值大小依次为锥栗(28.56 gH2O m-2s-1)>广东润楠(25.14 gH2O m-2s-1)>马尾松(25.02 gH2O m-2s-1)>木荷(21.62 gH2O m-2s-1),除广东润楠外,各优势树种湿季液流峰值均大于干季。湿季液流在20:00—21:00下降到最低值,滞后干季约1 h,干季液流下降速率较湿季迅速。夜间存在微弱液流,液流量相对稳定,变化幅度小。
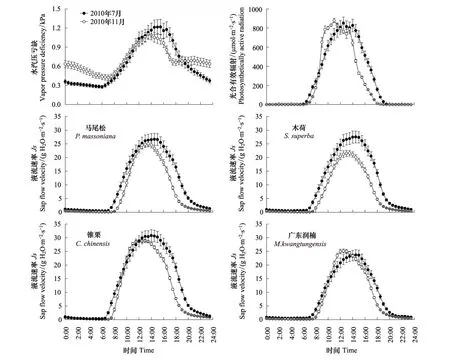
图2 干湿季光合有效辐射(PAR)、水汽压亏缺(VPD)及优势树种树干液流速率的日变化
2.3 树干液流对蒸腾驱动因子的响应
植物蒸腾是一个包括物理学机理和叶片生物学特性的过程,强度主要取决于土壤可利用水,液态水转化为水汽压所必须的能量以及叶片内外水汽压梯度[20],这三个因素可以分别由土壤含水量、PAR和VPD来衡量[21]。为了直观表达液流与蒸腾驱动因子的变化特征,绘制出了湿季(7月)与干季(11月)PAR、VPD变化曲线图,如图2所示。PAR、VPD与液流速率的变化一致,均呈现出单峰型曲线, PAR峰值区间存在明显波动,呈多峰型曲线。湿季(7月)土壤含水量变化较大,无明显规律,土壤含水量为(0.27±0.0004)m3/m3;干季(11月)土壤含水量呈明显下降趋势,下降幅度较小,土壤含水量为(0.21±0.0002)m3/m3。
环境因子对树干液流的影响是瞬时的、变动的,外界环境因子的波动会在树体内部的液流上反应出来[22]。本文将液流日变化序列与蒸腾驱动因子进行偏相关分析,结果如表2所示。从偏相关系数来看,在湿季(7月)和干季(11月),4种优势树种树干液流与PAR、VPD均呈现极显著正相关关系。湿季(7月)木荷树干液流与土壤含水量呈极显著负相关关系,干季(11月)4种优势树种均与土壤含水量呈极显著负相关关系。从偏相关系数大小来看, PAR、VPD为控制液流的主要驱动因子。利用多元统计分析,建立各优势树种与蒸腾驱动因子的回归方程,如表3所示。结果表明,与树干液流速率显著相关的各因子都入选了方程,7月和11月马尾松、锥栗、木荷、广东润楠的R2值分别为:0.87、0.87、0.89、0.87和0.72、0.81、0.77、0.77,说明方程均能较好的揭示液流速率和蒸腾驱动因子的变化规律。

表2 树干液流与蒸腾驱动因子偏相关分析Table 2 Partial correlations between sap flow and transpiration driving factors during the dry and wet seasons
**P<0.01
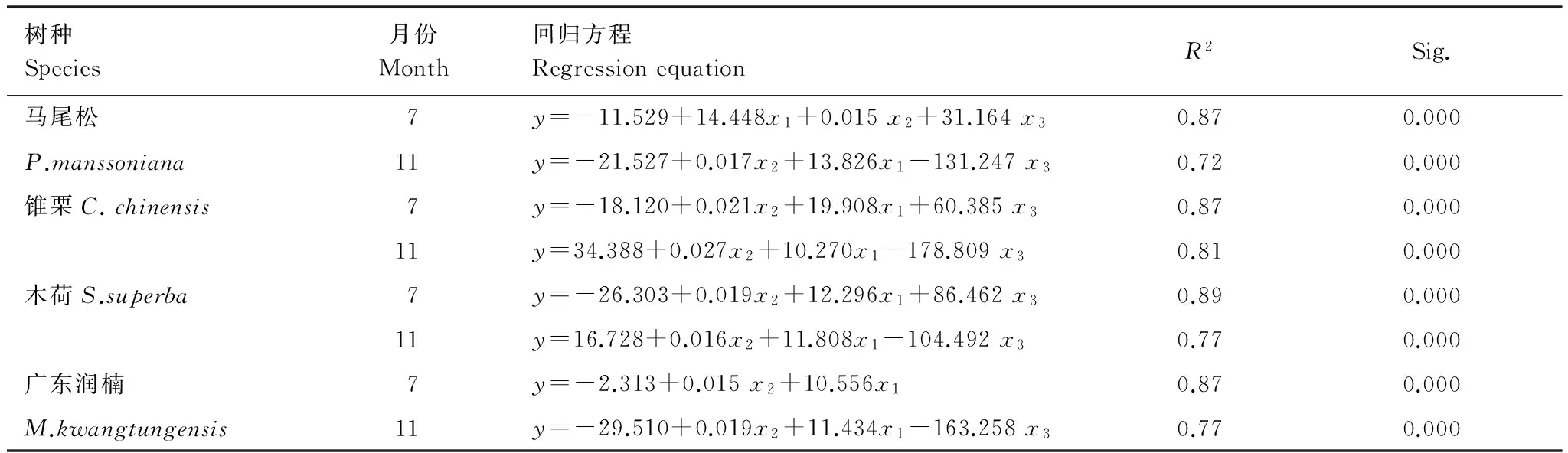
表3 干湿季树干液流速率(y)与蒸腾驱动因子(x)的回归方程Table 3 Multivariable regression models of sap flow velocity (y) and environmental factors (x) during the dry and wet seasons
2.4 整树蒸腾估算
将液流速率测定值扩展到整树蒸腾的尺度水平,马尾松、木荷、锥栗、广东润楠平均蒸腾速率Et(gH2O s-1株-1)由如下公式计算[21]:
Et=Js×As
(5)
式中,Js为瞬时液流密度(g H2O m- 2s-1),As为样树边材面积(m2)。夜间液流主要用于补充贮水部位的水分亏缺,因此不计入整树蒸腾量估算。结果表明,4种优势树种平均日蒸腾量湿季(7月)明显高于干季(11月),干湿季平均日蒸腾量均为木荷>锥栗>马尾松>广东润楠,然而平均边材面积则为木荷>马尾松>锥栗>广东润楠。马尾松、木荷、锥栗、广东润楠湿季平均日蒸腾量分别为29.52、39.29、30.40、9.41 kgH2O/d,干季分别为20.91、24.84、24.26、8.43 kgH2O/d。其中木荷湿季(7月)和干季(11月)日均蒸腾量最大值为广东润楠的2.9—4.2倍。

图3 干湿季优势树种平均日蒸腾量
单因素方差分析表明,除广东润楠,马尾松、木荷、锥栗干季(11月)和湿季(7月)平均日蒸腾量差异显著(图3)。树种日蒸腾量两两之间的比较,湿季除马尾松和锥栗差异不显著外,其他均差异显著;干季除木荷和锥栗差异不显著,其他均差异显著。
3 讨论与结论
马尾松、木荷、锥栗、广东润楠边材面积和胸径间均呈良好的幂函数关系,张宁南[23]、吴芳[24]、白云岗[25]等分别用二次函数、线性函数、幂函数很好地拟合了加勒比松、刺槐、胡杨等树种边材面积与胸径间的关系。由此可见,边材-胸径模型的表达形式在很大程度上取决于树种本身的生物学特性,树种不同,模型的表达形式可能不相同[26]。但不同地理区域以及海拔高度差异下,同一树种边材-胸径模型是否一致,需要深入探讨。
干湿季马尾松、木荷、锥栗、广东润楠树干液流日变化均呈现“昼高夜低”单峰曲线。这与黄德卫等[10]对鼎湖山海拔50 m针阔混交林样地同种优势树种的研究结果相似。然而本研究所处样地马尾松、木荷、锥栗、广东润楠湿季(7月)和干季(11月)平均液流速率分别为(10.07±1.47)、(10.07±1.51)、(12.20±1.73)、(8.35±1.28) gH2O m-2s-1;(7.22±1.33)、(6.41±1.15)、(9.78±1.64)、(7.37±1.34) gH2O m-2s-1,与黄德卫等[10]研究结果整体水平有差异,可能由如下原因导致。首先,不同立地条件下,相同树种的林木特征(如树龄、边材面积、胸径、树高等)区别较大;其次,海拔梯度造成的温度、湿度和土壤肥力等环境异质性极大影响植物的生长,导致植物在水分利用上产生一定的变化。
分析结果还表明同一环境条件下不同优势树种之间液流日变化规律一致,但液流速率差异较大,可能是因为树种之间的生物学结构的不同[27]。阔叶树广东润楠平均胸径小,边材导水面积小,随着树木高度的增加,水分运输路径增长,由于摩擦作用及水柱自身重力缘故,水分上行受阻[28],因此湿季(7月)阔叶树广东润楠平均液流速率、液流峰值及日液流量均低于其他优势树种。本研究还表明液流速率并不与胸径、边材面积成正相关关系。木荷边材面积较其他优势树种大,带动液流向上需要的蒸腾拉力也相应增加,因此在干季(11月)蒸腾拉力一定的情况下,液流速率相对较低。
马尾松、木荷、锥栗、广东润楠整树蒸腾有明显季节差异,湿季(7月)蒸腾活动明显比干季(11月) 旺盛。偏相关分析表明, PAR和VPD是冠层蒸腾的主要驱动因子,并能较好的拟合液流曲线。土壤含水量季节差异不明显,因此较适合作为长期观测及分析的因子。Wullschleger SD[29]等研究表明林分总蒸腾量主要由拥有最大边材面积的树种决定,朱丽薇[30]等也表明边材面积决定了树木蒸腾量的大小。本文研究表明树木蒸腾和针叶树与阔叶树生物学差异也相关,马尾松厚的针叶结构及由管胞运输水分,都限制了其蒸腾速率,因此边材面积相对小的锥栗蒸腾量明显大于马尾松[31]。干季(11月)土壤水分含量下降,植物普遍降低蒸腾量提高抗旱性[32],且随树种边材面积的增加,蒸腾减少的绝对量越大。
[1] 张小由, 康尔泗, 张智慧, 周茂先, 司建华. 沙枣树干液流的动态研究. 中国沙漠, 2006, 26(1): 146- 151.
[2] 张雷, 孙鹏森, 刘世荣. 树干液流对环境变化响应研究进展. 生态学报, 2009, 29(10): 5600- 5610.
[3] 孙龙, 王传宽, 孙慧珍, 张全智, 周晓峰. 生长季白桦树干液流密度的特征. 东北林业大学学报, 2007, 35(11): 13- 16.
[4] 马玲, 饶兴权, 赵平, 陆平, 蔡锡安, 曾小平. 马占相思整树蒸腾的日变化和季节变化特征. 北京林业大学学报, 2007, 29(1): 67- 73.
[5] 孙慧珍, 周晓峰, 康绍忠. 应用热技术研究树干液流进展. 应用生态学报, 2004, 15(6): 1074- 1078.
[6] Wullschleger S D, Meinzer F C, Vertessy R A. A review of whole-plant water use studies in tree. Tree Physiology, 1998, 18(8/9): 499- 512.
[7] 赵平, 饶兴权, 马玲, 蔡锡安, 曾小平. 马占相思树干液流密度和整树蒸腾的个体差异. 生态学报, 2006, 26(12): 4050- 4058.
[8] 梅婷婷, 王传宽, 赵平, 蔡锡安, 刘晓静, 张全智. 木荷树干液流的密度特征. 林业科学, 2010, 46(1): 40- 47.
[9] 黄玉清, 张中峰, 何成新, 赵平, 袁维园, 焦继飞, 尤业明. 岩溶区青冈栎整树蒸腾的季节变化. 应用生态学报, 2009, 20(2): 256- 264.
[10] 黄德卫, 张德强, 周国逸, 刘世忠, Dennis O, 李跃林. 鼎湖山针阔叶混交林优势种树干液流特征及其与环境因子的关系. 应用生态学报, 2012, 23(5): 1159- 1166.
[11] 池波, 蔡体久, 满秀玲, 李奕. 大兴安岭北部兴安落叶松树干液流规律及影响因子分析. 北京林业大学学报, 2013, 35(4): 21- 26.
[12] 曹恭祥, 熊伟, 王彦辉, 于澎涛, 高娃, 刘海龙, 陈亚峰. 宁夏六盘山华山松树干液流的动态研究. 内蒙古农业大学学报: 自然科学版, 2010, 31(2): 42- 47.
[13] 周存宇, 张德强, 王跃思, 周国逸, 刘世忠, 唐旭利. 鼎湖山针阔叶混交林地表温室气体排放的日变化. 生态学报, 2004, 24(8): 1738- 1741.
[14] 王春林, 周国逸, 唐旭利, 王旭, 周传艳, 于贵瑞, 唐力生, 孟泽. 鼎湖山针阔叶混交林生态系统呼吸及其影响因子. 生态学报, 2007, 27(7): 2659- 2668.
[15] 彭华贵, 杜彦君, 李炯, 欧阳学军, 黄忠良. 南岭大顶山与鼎湖山常绿阔叶林种群分布格局对比研究. 生态环境, 2006, 15(4): 770- 774.
[16] Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 1987, 3(4): 309- 320.
[17] Cˇermák J, Deml M, Penka M. A new method of sap flow rate determination in trees. Biologia Plantarum, 1973, 15(3): 171- 178.
[18] Campbell G S, Norman J M. An introduction to environmental biophysics. 2nd ed. NewYork: Springer-Verlag, 1998: 5- 9.
[19] 涂洁, 刘琪璟. 亚热带红壤区湿地松边材液流的尺度扩展及其对环境因子的响应. 水土保持学报, 2007, 21(2): 197- 200.
[20] 康绍忠, 刘晓明, 熊运章. 土壤-植物-大气连续体水分传输理论及其应用. 北京: 水利电力出版社, 1994: 51- 84.
[21] 赵平, 邹绿柳, 饶兴权, 马玲, 倪广艳, 曾小平, 蔡锡安. 成熟马占相思林的蒸腾耗水及年际变化.生态学报, 2011, 31(20): 6038- 6048.
[22] 林平, 李吉跃, 马达. 北京山区油松林蒸腾耗水特性研究. 北京林业大学学报, 2006, 28(S1): 47- 50.
[23] 张宁南, 徐大平, Morris J, 杨曾奖, 周光益. 雷州半岛加勒比松人工林旱季液流特征与耗水量研究. 林业科学研究, 2007, 20(5): 591- 597.
[24] 吴芳, 陈云明, 于占辉. 黄土高原半干旱区刺槐生长盛期树干液流动态. 植物生态学报, 2010, 34(4): 469- 476.
[25] 白云岗, 张江辉, 王新友, 李冰, 潘洪彬. 塔里木盆地胡杨单木与林分耗水量的尺度转换研究. 中国水利, 2008, (5): 24- 25.
[26] 涂洁, 刘琪璟, 李海涛, 林耀明. 江西千烟洲湿地松生长旺季树干液流动态及影响因素分析. 林业科学, 2008, 44(1): 46- 51.
[27] 刘明国, 唐敬超, 王玉涛, 丁士富. 辽西地区油松树干液流变化规律及影响因子研究. 沈阳农业大学学报, 2011, 42(2): 175- 179.
[28] 刘晓静, 赵平, 王权, 蔡锡安, 曾小平. 树高对马占相思整树水分利用的效应. 应用生态学报, 2009, 20(1): 13- 19.
[29] Wullschleger S D, Hanson P J, Todd D E. Transpiration from a multi-species deciduous forest as estimated by xylem sap flow techniques. Forest Ecology and Management, 2001, 143(1/3): 205- 213.
[30] 朱丽薇, 赵平, 蔡锡安, 曾小平, 邹绿柳, 王权. 荷木人工林蒸腾与冠层气孔导度特征及对环境因子的响应. 热带亚热带植物学报, 2010, 18(6): 599- 606.
[31] 孙双峰, 黄建辉, 林光辉, 韩兴国. 三峡库区岸边共存松栎树种水分利用策略比较. 植物生态学报, 2006, 30(1): 57- 63.
[32] 刘晓静, 赵平, 蔡锡安, 饶兴权, 曾小平. 不同径级马占相思整树蒸腾的湿、干季变化. 生态学报, 2009, 29(2): 619- 626.
Sap flow characteristics of four dominant tree species in a mixed conifer- broadleaf forest in Dinghushan
CHENG Jing1,2, OUYANG Xu1,2, HUANG Dewei1,3, LIU Shizhong1, ZHANG Deqiang1, LI Yuelin1,*
1SouthChinaBotanicalGarden,ChineseAcademyofSciences,Guangzhou510650,China2UniversityofChineseAcademyofSciences,Beijing100049,China3ExperimentalCenterofTropicalForestry,Pingxiang532600,China
The plant water use efficiency is not only crucial for plant to maintain a relatively high growth rate but also an important metric of water balance in forest ecosystem. Recent years, sap flow measurement has been widely used as a basis for assessing the water exchange between tree crown and ambient atmosphere for it can keep trees in natural condition without damaging the growth of trees. Many studies on trees′ sap flow in single species plantations have been carried out across China, but the studies in natural forests are still few. We measured sap flow of four dominant tree species (Pinusmanssoniana,Schimasuerba,CastanopsischinensisandMachiluskwangtungensis) in a mixed conifer-broadleaf natural forest in the Dinghushan Reserve of South China using the Granier′s thermal dissipation probe method. The sap flow was measured daily in July and November, 2010, which represented the wet and dry seasons in the region, respectively. The environmental factors such as photosynthetically active radiation (PAR), air temperature (T), relative air humidity (RH) and soil moisture were also recorded. The aims of this study were (1) to investigate the characteristics of sap flow of four dominant tree species, (2) to estimate their relationships with the driving factors of transpiration, (3) and to compare the transpiration rates of different tree species calculated from sap flux density combing with trees′ morphological characteristics. The results showed that sapwood area of the four dominant tree species had a significantly (P<0.05) positive relationship with diameter at breast height (DBH). The coefficient of determination R2of four trees species were 0.99、0.96、0.93、0.81 respectively. Sap flux density displayed an obvious pattern with higher values during daytime and also showed a significant seasonal variation with higher value in wet season. The average sap flow velocity ofP.manssoniana,S.superba,C.chinensisandM.kwangtungensiswere (10.07±1.47), (10.07±1.51), (12.20±1.73), (8.35±1.28) gH2O m-2s-1in wet season, and were (7.22±1.33), (6.41±1.15), (9.78±1.64), (7.37±1.34) gH2O m-2s-1in dry season. Contrast to previous study in Dinghushan Reserve of South China at an altitude of 50m, our research found that sap flow velocity displayed a significant variation along the gradient of altitude, which was induced by the differentiation of microenvironment and plant morphological characteristics. Partial correlation analysis indicated that photosynthetically active radiation (PAR) and vapor pressure deficiency (VPD) were the two main environmental factors determining the tree transpiration in both seasons. Transpiration of the four dominant tree species was much higher during the wet season than in the dry season. The average daily transpiration rates ofP.manssoniana,S.superba,C.chinensisandM.kwangtungensisduring the wet season were 29.52, 39.29, 30.40, 9.41 kgH2O/d, respectively, while those during the dry season the rates were 20.91, 24.84, 24.26 and 8.43 kgH2O/d, respectively. The order of the average daily transpiration was:S.superba>C.chinensis>P.manssoniana>M.kwangtungensisboth in wet and dry seasons. Such difference in daily transpiration among tree species was mainly determined by the sapwood area and biological characteristics.
Granier′s thermal dissipation probe; mixed conifer-broadleaf forest; sap wood area; sap flow; daily transpiration
国家自然科学基金项目(31170375); 中国科学院知识创新工程重要方向项目(KZCX2-YW-GJ01); 广东省自然科学基金项目(9151008901000130)
2013- 10- 20;
2014- 08- 28
10.5846/stxb201310202533
*通讯作者Corresponding author.E-mail: yuelin@scib.ac.cn
程静, 欧阳旭, 黄德卫, 刘世忠, 张德强, 李跃林.鼎湖山针阔叶混交林4种优势树种树干液流特征.生态学报,2015,35(12):4097- 4104.
Cheng J, Oyang X, Huang D W, Liu S Z, Zhang D Q, Li Y L.Sap flow characteristics of four dominant tree species in a mixed conifer-broadleaf forest in Dinghushan.Acta Ecologica Sinica,2015,35(12):4097- 4104.
猜你喜欢
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
小作家报·教研博览(2020年35期)2020-09-10
光谱学与光谱分析(2020年1期)2020-01-08
防护林科技(2019年12期)2019-04-14
快乐语文(2018年14期)2018-06-06
防护林科技(2017年4期)2017-04-27
中国科技纵横(2016年19期)2016-12-10
花卉(2016年14期)2016-07-31
东北林业大学学报(2015年6期)2015-04-03
