
黑土母质熟化过程微生物群落碳源代谢特征
2015-02-05 01:23张志明韩晓增
生态学报 2015年21期
张志明,韩晓增
中国科学院东北地理与农业生态研究所海伦农田生态系统国家野外科学观测研究站,哈尔滨 150081
黑土母质熟化过程微生物群落碳源代谢特征
张志明,韩晓增*
中国科学院东北地理与农业生态研究所海伦农田生态系统国家野外科学观测研究站,哈尔滨 150081
东北黑土区由于表土丧失,母质裸露等问题,严重威胁粮食生产和生态环境,通过揭示母质肥力形成过程中微生物群落的碳源利用特征可以让人们更好的理解肥力形成过程中相关微生物过程。以海伦农田生态系统国家野外科学观测研究站长期定位试验为材料,研究母质不同熟化过程下微生物群落碳源代谢特征,结果表明:苜蓿草熟化过程(PAfl)、农田化肥配施有机物熟化过程(PCS+F+OM)可培养微生物数量最多。自然熟化过程(PNat)、苜蓿草熟化过程、农田化肥农产品全循环熟化过程(PCS+F+BM)、农田化肥加有机肥配合型熟化过程可以将微生物活性提高到常规农田黑土的水平。不同熟化过程下微生物群落对单个碳源的利用不同。主成分分析表明自然熟化过程、农田化肥投入熟化过程(PCS+F)、农田化肥农产品全循环熟化过程、农田化肥加有机肥配合型熟化过程与常规农田黑土微生物代谢特征类似。研究表明不同母质熟化过程通过影响微生物群落结构导致对各个碳源的利用发生变化,最终改变了整体碳源代谢特征。
黑土母质;熟化过程;微生物;代谢特征
我国东北平原黑土区,主要分布在小兴安岭和松嫩平原腹地之间的过渡带[1],所以形成了大量坡耕地,这些坡耕地开垦后,由于侵蚀导致表土迁移流失,严重的地区母质裸露。再加上工程建设的取土和弃土,黑土表层被破坏,导致大量农田母质裸露[2]。在平原地区,对土壤掠夺性经营,导致表土退化形成劣质的土壤[3],即是母质裸露的表现。这些地区土壤的原始母质裸露后,丧失了土壤应有的生产植物的能力,同时对生态环境也带来了危害[4]。基于对退化严重土壤肥力恢复的需求,将2 m以下的黑土母质移至地表,分别模拟自然熟化过程、种植豆科牧草熟化过程;在模拟农田过程中,分别设置了无肥、化肥、化肥加有机肥和在施用化肥的条件下将所收获的农作物(秸秆和籽粒)全部还田,试图了解母质熟化过程中的各种肥力属性的变化特征。
土壤肥力的恢复与重建的引擎是土壤微生物主导下的生物化学过程[5],土壤微生物的碳代谢过程又是土壤生物化学过程非常重要的组成部分。1991年,Garland和Mills首次应用Biolog系统分析微生物群落的碳源代谢特征[6]。随后众多学者利用此方法研究在施肥[7-8]、耕作[9-10]、不同植被覆盖[11-12]和不同土壤类型[13]条件下土壤微生物群落碳源代谢特征,并以此来反映土壤肥力的变化。但是在土壤侵蚀严重、母质裸露地区,对土壤肥力恢复重建过程中微生物群落碳代谢过程的研究较少。本文通过研究模拟黑土侵蚀严重、母质裸露在地表后不同熟化过程下微生物群落碳源代谢的变化特征,探讨不同措施对母质肥力恢复的影响,为应对黑土耕地资源减少和开发新的耕地资源提供思路和理论依据。
1 材料与方法
1.1 试验地点
黑土母质熟化试验区设置于海伦农田生态系统国家野外科学观测研究站试验田内(47°27′N, 126°55′E),位于中国黑土带中部,海拔234 m,区域内土壤为典型中厚层黑土。试验区为温带大陆性季风气候,冬季寒冷干燥,夏季高温多雨,雨热同季,全年降水量500—600 mm,无霜期120—130 d,≥10 ℃有效积温2400—2500 ℃[14]。
1.2 试验设计
试验始于2004年6月。黑土母质是第四纪黄土状母质,母质出现在土壤剖面的155 cm以下,试验所用母质取自表层以下2.0—2.5 m深的有代表性的母质,移入地表混匀后,等量放入24个微区中。每个微区平面长1.4 m,宽1 m,深0.6 m,底部用200目双层尼龙网将试验母质和原土分开(原土性状与母质相近),不能生长根系但水分可以运移,四周均用钢筋混凝土铸成,可防水侧渗。试验设置6个处理:(1)母质自然熟化过程(PNat),没有任何人为措施,地上植物为自然生长,每年冬季枯死后自然还田;(2)母质苜蓿草熟化过程(PAlf),人工播种多年生苜蓿,不加任何其他措施,每年秋季地上生物量剪碎覆盖表面还田;(3)农田耕作无肥投入熟化过程(PCS-F);(4)农田耕作化肥投入熟化过程,仅施用NPK化肥(PCS+F);(5)农田耕作施用化肥农产品全循环熟化过程,NPK化肥+每年地上所有收获物(籽粒和秸秆)全量还田(PCS+F+BM);(6)农田耕作化肥加有机肥配合型熟化过程,NPK化肥+秸秆+炒熟豆粉(PCS+F+OM),模拟有机肥。每个处理4次重复,随机排列。(7)试验还采集距微区试验地100m处常规农田表层黑土(SCS+F)作为对照,具体试验设置见表1。处理代号含有CS代表大豆-玉米轮作种植方式;P代表供试土壤为母质;S代表供试土壤为常规农田表层耕作黑土。处理1—2免耕;处理3—7翻耕土层深度为17 cm。不同作物每年肥料用量见表1。

表1 不同作物施肥量Table 1 Different crop fertilizer rate
1.3 样品采集
样品于2012年春天播种前采集,系收获后玉米茬口,取样时保留1个区组不取样,取另外3个区组的小区土壤样品供测试研究,仅采集表层,免耕处理为0—10 cm,耕作处理根据翻耕深度采集0—17 cm土样,每个小区取样方法为每一层土壤全部取出混匀,多次四分法取样最终为1个样品。其中采集前将PNat和PAlf两个处理地表植物残体除去。样品取完后用封口袋封存,放入冰盒中带回实验室,在4℃冰箱中保存,以备分析。
1.4 试验方法
土壤微生物三大菌群的数量采用平板菌落计数法测定[15]。
Biolog EcoPlate接种和培养:称取相当于5 g烘干土重的鲜土加入到装有45 mL灭菌生理盐水(0.85%)的三角瓶中,超声1 min,然后在振荡器上振荡40 min混匀。土壤溶液稀释至10-3浓度梯度,向ECO板中每孔加入150 μL土壤稀释液, 28 ℃保湿避光培养。每隔24 h用Biolog 自动读数仪测量590 nm(颜色和浊度)和750 nm(浊度)波长下的数值[16]。
1.5 数据分析
按照Classen[16]方法计算:
(1)平均颜色变化率(AWCD)
AWCD=∑(C590-750)/n
(2)Shannon指数H(评估物种丰富度)
H=-∑piln(pi)
(3)Simpson指数(评估某些常见种的优势度)
D=1-∑(pi)2
(4)Mclntosh指数(评估群落物种均匀度)
式中,C是反应孔吸光值,n是碳源数。pi为第i孔相对吸光度值与整个平板相对吸光度值总和的比率,ni是第i孔的相对吸光值。
试验结果的方差分析及主成分分析采用SPSS19.0进行。
2 结果
2.1 可培养微生物数量变化特征
可培养微生物虽然只占土壤微生物总数的0.1%—1%[17],但它们的功能是可知的,在不同母质熟化过程中,可培养微生物的丰度和类群有明显不同的特征(表2)。母质自然熟化过程中基本检测不到放线菌,说明放线菌这个类群在从母质到具有肥力的土壤熟化过程中,响应速度较细菌和真菌慢得多;且放线菌在母质熟化过程中,对肥料相当敏感,在无肥料投入的3个处理中数量较少。在人工播入牧草免耕条件下,土壤可培养微生物总数相当于母质自然熟化过程的可培养微生物总数的2倍,证明了豆科植物苜蓿对土壤微生物有较强的促进作用。
在母质熟化的农田过程中,不同的施肥方式对土壤可培养微生物数量有不同的促进作用。无肥是可培养微生物数量的限制因子,与优化的施肥方式(处理6)相比,仅占其1/10。从施用化肥熟化过程与化肥加有机物配合施用熟化过程比较可以看出化肥在母质熟化初期对可培养微生物数量的促进作用更大。
2.2 不同黑土熟化过程土壤微生物群落功能多样性特征
2.2.1 土壤微生物群落平均颜色变化率的动态变化
母质不同熟化过程中微生物群落的平均颜色变化率(AWCD)是不同的(图1)。AWCD越高,微生物群落活性越大。苜蓿草通过2方面影响土壤微生物,一方面表层苜蓿根系密度大,对土壤切割使改变母质三相比;另一方面豆科植物激发母质根瘤菌生长,增加母质中养分含量,而且每年地上生物量自然还田也增加了养分。播种玉米和大豆(PCS-F)的母质中微生物活性最小,自然恢复草地(PNat)母质中的微生物活性居中。在农田熟化过程中,肥料对母质中微生物活性的影响是复杂的,化肥(PCS+F)是母质中微生物活性降低的直接控制因子,有机物料(PCS+F+BM;PCS+F+OM)提升母质中微生物活性可以达到农田表层黑土(SCS+F)的水平。

表2 不同处理对微生物数量的影响Table 2 Effect of different application on microorganism amounts
同列不同小写字母代表处理间差异显著(P<0.05);PNat: 母质自然熟化过程Parent-Natural;PAlf: 母质苜蓿草熟化过程Parent-Alfalfa;PCS-F: 农田耕作无肥投入熟化过程Parent Corn-Soybean-No Fertilizer;PCS+F: 农田耕作化肥投入熟化过程Parent Corn-Soybean-Fertilizer;PCS+F+BM: 农田耕作施用化肥农产品全循环熟化过程Parent Corn-Soybean+Fertilizer+Biomass cycle;PCS+F+OM: 农田耕作化肥加有机肥配合型熟化过程Parent Corn-Soybean+Fertilizer+Oganic Matter;SCS+F: 常规农田表层黑土Stander Soil Corn-Soybean+Fertilizer

图1 土壤微生物群落AWCD随培养时间的变化 Fig.1 AWCD changes with incubation time of different treatmentsPNat: 母质自然熟化过程Parent-Natural;PAlf: 母质苜蓿草熟化过程Parent-Alfalfa;PCS-F: 农田耕作无肥投入熟化过程Parent Corn-Soybean-No Fertilizer;PCS+F: 农田耕作化肥投入熟化过程Parent Corn-Soybean-Fertilizer;PCS+F+BM: 农田耕作施用化肥农产品全循环熟化过程Parent Corn-Soybean+Fertilizer+Biomass cycle;PCS+F+OM: 农田耕作化肥加有机肥配合型熟化过程Parent Corn-Soybean+Fertilizer+Oganic Matter;SCS+F: 常规农田表层黑土Stander Soil Corn-Soybean+Fertilizer
2.2.2 土壤微生物群落对各类碳源的利用特征
从图2可知,黑土母质熟化过程中,各处理母质中的微生物主要利用的碳源均为糖类和氨基酸类,其次是羧酸类和聚合物类。但是从表3中知道,母质自然熟化过程、母质苜蓿草熟化过程和常规农田土壤微生物群落利用率最高的5个碳源种类一致,都是氨基酸类和糖类。在有人为影响的农田熟化过程中,微生物群落利用最高的5个碳源与成熟的常规农田黑土不一致,没有肥料投入的熟化过程(PCS-F)中利用率最高的5个碳源没有糖类;投入化肥后(PCS+F)后,依然有羧酸和聚合物类,但增加了2个糖类碳源;施用秸秆(PCS+F+BM)和秸秆+豆粉(PCS+F+OM)后利用率最高的5个碳源中没有聚合物类,但依然有羧酸类碳源,而且在PCS+F+OM中有3个氨基酸类碳源。
2.2.3 主成分分析
不同熟化过程下土壤微生物群落对31种碳源利用的主成分分析(图3),本文共提取4个主成分,前2个主成分的方差贡献率分别是29.56%和20.80%,第3、4个主成分贡献率是18.85%和11.83%。所有处理在PC1轴上分成3类,母质苜蓿草熟化过程为一类;农田化肥熟化过程为一类;其余熟化过程为一类,而且这一类与常规农田黑土微生物群落碳源利用特征相似。对PC1产生相对较大影响的碳源是糖类和氨基酸类(表4)。

图2 不同处理下土壤微生物群落对Biolog板中6类碳源相对利用率 Fig.2 Relative use efficiency of six kinds of carbon sources in Biolog EcoPlate by soil microbe community of different treatments
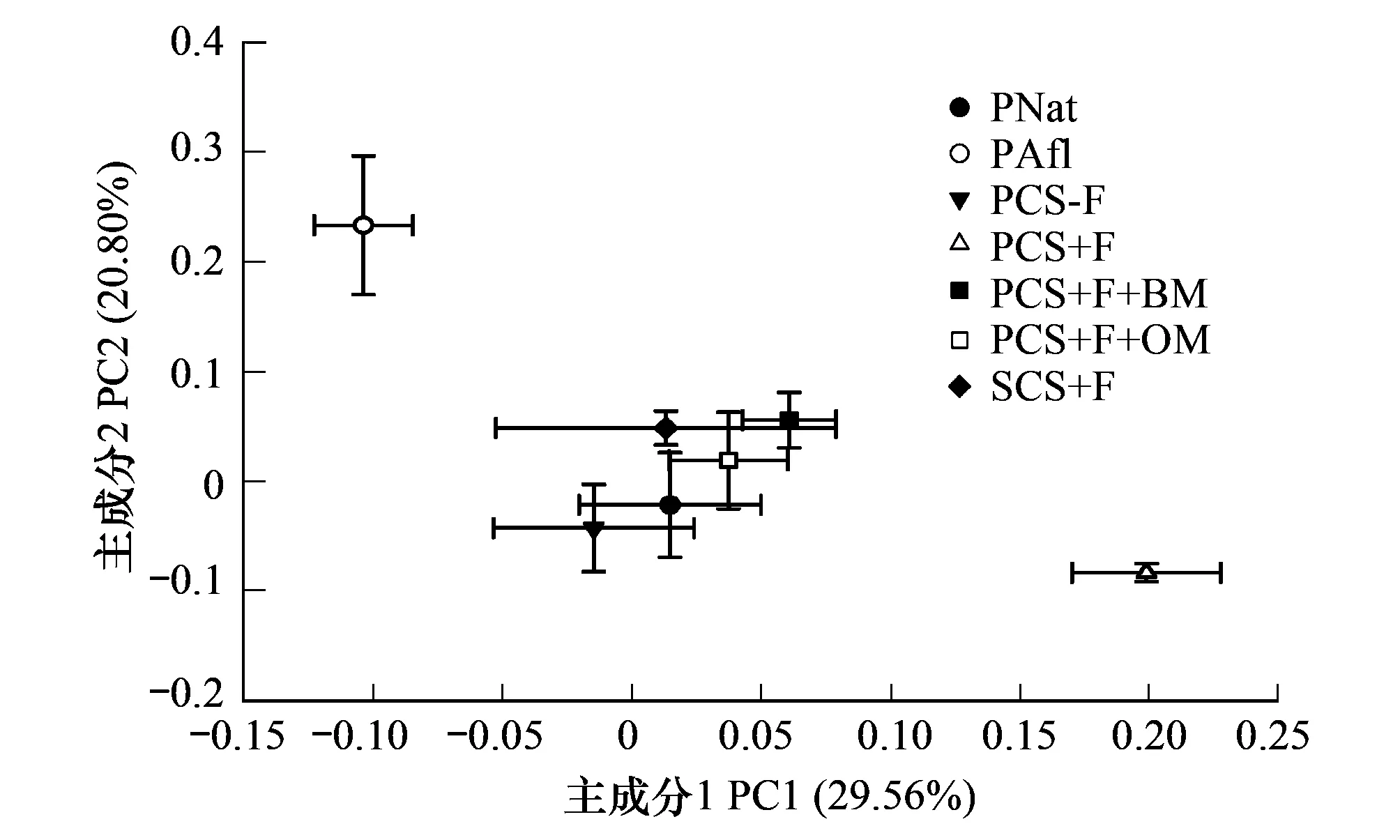
图3 不同熟化过程下的土壤微生物群落对31种碳源利用的主成分分析 Fig.3 Principal components analysis of carbon source utilization by soil microbial communities in soil maturation process

表3 各处理利用最高的5个碳源Table 3 The highest utilization rate of 5 carbon sources in all treatments

表4 各碳源与PC1和PC2的相关系数Table 4 Correlation coefficients between all substrates and PC1 and PC2
3 讨论
将黑土母质从地下2—2.5 m处相对封闭的环境移到地表相对开放的环境中,在不同的熟化过程中可培养微生物数量产生不同的变化。植被类型(PAlf,PCS-F)和肥料(PCS-F,PCS+F+OM)对母质中可培养微生物数量影响非常大。可培养放线菌对肥料非常敏感,在没有肥料投入的熟化过程中放线菌基本检测不到。本试验选取附近常规农田表层黑土作为对照,常规农田表层黑土的细菌∶放线菌∶真菌=364.54∶3.77∶1;农田化肥有机肥配合熟化过程的细菌∶放线菌∶真菌=371.79∶1.25∶1;农田化肥农产品全循环熟化过程的细菌∶放线菌∶真菌=390.7∶1.52∶1;农田化肥投入熟化过程的细菌∶放线菌∶真菌=368.26∶1.43∶1。农田无肥投入熟化过程的细菌∶真菌=119.27∶1;母质苜蓿草熟化过程的细菌∶真菌=1299.87∶1;母质自然熟化过程的细菌∶真菌=1438.26∶1,母质苜蓿草熟化过程的可培养微生物总数是自然熟化过程的2倍,但是各类群微生物数量比值相似,说明播种苜蓿比自然恢复对母质微生物数量的发展有更好的作用。各个处理与自然熟化过程比较说明农田化肥有机肥配合熟化过程、农田化肥农产品全循环熟化过程和农田化肥投入熟化过程的微生物数量性状发展逐渐接近常规农田黑土;母质苜蓿草熟化过程和母质自然熟化过程中细菌群落繁衍的极为迅速;农田无肥投入熟化过程各类微生物均发展很慢。
在本研究中可以看出不同植被类型对母质中微生物活性有很强的控制作用。AWCD越高微生物活性越高[18],代谢活动越旺盛,合成和分解的养分越多,各个养分元素循环的速度快,对母质肥力的形成有更强的促进作用。苜蓿对微生物群落的活性有很好的提高作用。杜天庆[19]、杨珍平[20]研究表明,苜蓿等豆科植物对黄土母质孔隙度、容重和团聚体有很好的改良作用,固氮作用可以提高土壤氮素营养及有机质含量,对母质有良好的改良效果。本试验结果看出,不投入有机物质种植农作物对微生物活性提高效果最差。而母质自然熟化过程前两年没有生物量产出,之后几年植被逐渐增加,经过8a地上地下产生的生物量和可培养微生物数量仍远小于母质苜蓿草熟化过程,但母质自然熟化过程的AWCD与母质苜蓿草熟化过程已经在同一水平。
肥料是微生物群落活性的关键影响因素。秸秆含有大量碳水化合物以及氮、磷、钾等营养元素,秸秆还田具有改善土壤结构,补充土壤养分,激发微生物活性等作用[21],且与无机肥料配施对改善土壤养分状况和维持土壤微生物活性有明显效果[8]。祁阳红壤实验站“生土熟化”长期定位试验的研究表明,有机无机肥配施和秸秆还田是快速提高母质肥力的措施[22],有机质含量与有机物投入量呈显著正相关[23]。农田熟化过程的地上植被完全相同,而化肥有机肥配合熟化过程和化肥农产品全循环熟化过程的AWCD显著高于无肥投入熟化过程,处理间仅有施肥上的不同,说明向母质中施入化肥加秸秆和豆粉等有机物质,通过增加了微生物利用的底物,从而提高了微生物群落的代谢活性。常规农田黑土的可培养微生物数量显著低于农田化肥有机肥配合熟化过程、农田化肥农产品全循环熟化过程,但是AWCD却是在同等水平;农田化肥投入熟化过程的可培养微生物数量显著高于常规农田黑土,但AWCD与常规农田黑土相比却低得多,说明种植作物仅施用化肥熟化方式可以刺激母质中微生物数量增加,但对微生物群落的活性的提高程度有限,而化肥与有机物配施熟化方式不仅能提高可培养微生物数量,而且能将其活性提高到常规农田黑土的水平。
母质在不同的熟化过程下各类碳源的利用情况也发生了变化。各个熟化过程中微生物群落主要利用糖类和氨基酸类碳源,其次是羧酸和聚合物类碳源,与常规农田黑土一致。常规农田黑土利用最高的5个碳源是氨基酸类(1个)和糖类(4个),母质自然熟化过程和母质苜蓿草熟化过程利用最高的5个碳源是糖类和氨基酸类。农田无肥投入熟化过程利用最高的5个碳源是氨基酸类、羧酸类和聚合物类;农田化肥投入熟化过程是氨基酸类、糖类、羧酸类和聚合物类;农田化肥农产品全循环熟化过程和农田化肥有机肥配合熟化过程利用最高的5个碳源是氨基酸类、糖类和羧酸类。农田熟化过程中,微生物对碳源的利用情况有很大不同,但处理之间只有施入物质的差异,所以投入不同是导致碳源利用差异的唯一原因。长期施用某类物质会使微生物利用的碳源趋于稳定,施入熟大豆粉进入土壤中,刺激了以蛋白质为碳源、氮源和能源的微生物的生长。而且有机物的施入减少了微生物对羧酸类和聚合物类碳源的利用率。不同熟化方式下微生物群落对31个碳源的利用表现出不同的规律,也说明不同熟化方式对母质肥力形成过程中对土壤碳库的影响不同。
主成分分析共提取了4个主成分,本文只对前两个主成分进行分析。分析结果表明不同熟化过程影响下的土壤微生物群落对碳源利用情况差异显著(图3)。整体碳源代谢特征的不同也间接体现了不同熟化措施对微生物群落结构的影响。除母质苜蓿草熟化过程和农田化肥投入熟化过程外,其他熟化过程的整体微生物碳源代谢特征与常规黑土农田相似,不同的熟化过程对母质中微生物在氨基酸类和糖类碳源利用能力上的影响是导致微生物群落碳源代谢特征变化的主要原因。母质苜蓿草熟化过程由于苜蓿的固氮作用激发了与氮素循环有关的微生物大量生长;农田化肥投入熟化过程连续8年只施用无机肥料会对母质pH值产生影响,虽然增加了可培养微生物数量,但也积累了毒害物质和抑制某些类微生物类群的发展,所以这两个熟化过程的碳源利用特征与其他熟化过程不同。
不同熟化过程使母质微生物对碳源利用能力、碳源利用率和群落功能表现上产生了较大差异,这些信息反映了母质微生物群落对不同熟化过程的响应程度,可以为土壤退化严重地区研究土壤肥力重建提供依据。
4 结论
母质由地下相对封闭环境到地表相对开放环境后,为使母质肥力快速形成采取了不同的熟化措施,得出以下结论:
自然熟化过程、苜蓿草熟化过程、农田耕作化肥投入熟化过程和农田化肥有机物配施熟化过程都可以显著提高黑土母质中可培养微生物数量,农田耕作化肥投入熟化过程和农田化肥有机物配施熟化的微生物数量性状发展逐渐接近成熟黑土;母质自然熟化过程、母质苜蓿草熟化过程中细菌群落繁衍的极为迅速;农田耕作无肥投入熟化过程中各类微生物均发展很慢。
植被和肥料对微生物活性有很强的影响,自然熟化过程、苜蓿草熟化过程、农田化肥有机物配施熟化过程显著提高母质中微生物的活性,达到成熟黑土的水平。主成分分析显示母质苜蓿草熟化过程和农田化肥投入熟化过程对母质微生物群落产生了不同的影响,所以整体碳源利用特征与常规农田黑土和其他熟化过程不同。
[1] 阎百兴, 杨育红, 刘兴土, 张树文, 刘宝元, 沈波, 王玉玺, 郑国相. 东北黑土区土壤侵蚀现状与演变趋势. 中国水土保持. 2008, (12): 26- 30.
[2] 王小兵, 吴元元, 邓玲. 东北黑土区黑土退化防治与保护研究. 资源与产业, 2008, 10(3): 81- 83.
[3] 于磊, 张柏. 中国黑土退化现状与防治对策. 干旱区资源与环境, 2004, 18(1): 99- 103.
[4] 张兴义, 隋跃宇, 宋春雨. 农田黑土退化过程. 土壤与作物, 2013, 2(1): 1- 6.
[5] 赵其国. 我国红壤的退化问题. 土壤, 1995, (6): 281- 285.
[6] Garland J L, Mills A L. Classification and characterization of heterotrophic microbial communities on the basisof patterns of community-level-sole-carbon-source-utilization. Applied and Environmental Microbiology. 1991, 57(8): 2351- 2359.
[7] Li F L, Liu M, Li Z P, Jiang C Y, Han F X, Che Y P. Changes in soil microbial biomass and functional diversity with a nitrogen gradient in soil columns. Applied Soil Ecology, 2013,64:1- 6.
[8] 罗希茜, 郝晓晖, 陈涛, 邓婵娟, 吴金水, 胡荣桂. 长期不同施肥对稻田土壤微生物群落功能多样性的影响. 生态学报, 2009, 29(2): 740- 748.
[9] Yan F, McBratney A B, Copeland L. Functional substrate biodiversity of cultivated and uncultivated A horizons of vertisols in NW New SouthWales. Geoderma, 2000, 96(4): 321- 343.
[10] 姬艳艳, 张贵龙, 张瑞, 刘玉升, 杨殿林, 王彩灵. 耕作方式对农田土壤微生物功能多样性的影响. 中国农学通报, 2013, 29(6): 117- 123.
[11] Yin R, Deng H, Wang H L, Zhang B. Vegetation type affects soil enzyme activities and microbial functional diversity following re-vegetation of a severely eroded red soil in sub-tropical China. Catena, 2014, 115: 96- 103.
[12] 刘明, 李忠佩, 张桃林. 不同利用方式下红壤微生物生物量及代谢功能多样性的变化. 土壤, 2009, 41(5): 744- 748.
[13] Sun B, Wang X Y, Wang F, Jiang Y, Zhang X X. Assessing the relative effects of geographic location and soil type on microbial communities associated with straw decomposition. Applied and Environmental Microbiology, 2013,79(11):3327- 3335.
[14] 李晓慧, 韩晓增, 王树起, 王守宇, 严君. 土壤微生物对不同作物根系活动的响应. 农业系统科学与综合研究, 2010, 26(2): 156- 159.
[15] 林先贵. 土壤微生物研究原理与方法. 高等教育出版社, 2010.
[16] Classen A T, Boyle S I, Haskins K E, Overby S T, Hart S C. Community-level physiologicalprofiles of bacteria and fungi plate type and incubation temperature influence on contrasting soils. FEMS Microbiology Ecology, 2003,44(3): 319- 328.
[17] Kirk J L, Beaudette L A, Hart M, Moutoglis P, Klironomos J N, Lee H, Trevors J T. Method of studying soil microbial diversity. Journal of Microbiological Methods, 2004, 58(2): 169- 188.
[18] Haack SK, Garchow H, Klug M J, Fomey L J. Analysis of factors affecting the accuracy, reproducibility, and interpretation of microbial community carbon source utilization patterns. Applied and Environmental Microbiology, 1995,61(4): 1458- 1468.
[19] 杜天庆, 苗果园, 郝建平, 杨锦忠, 崔福柱, 薛建福. 不同类型豆科植物对黄土母质生土的改土效果. 水土保持学报, 2011, 25(6): 76- 80, 86.
[20] 杨珍平, 郝教敏, 卜玉山, 高志强, 苗果园. 苜蓿根系生长对黄土母质生土的改良效应. 草地学报, 2012, 20(3): 489- 496.
[21] Debosza K,Peter H R,Pedersena A R.Temporal variations in microbial biomass C and cellulolytic enzyme activity in arable soils: effects of organic matter input.Applied Soil Ecology,1999,13 (3): 209- 218.
[22] 于寒青, 徐明岗, 吕家珑, 包耀贤, 孙楠, 高菊生. 长期施肥下红壤地区土壤熟化肥力评价. 应用生态学报, 2010, 21(7): 1772- 1778.
[23] 于寒青, 孙楠, 吕家珑, 高菊生, 徐明岗, 王伯仁. 红壤地区三种母质土壤熟化过程中有机质的变化特征. 植物营养与肥料学报, 2010, 16(1): 92- 98.
Microbial metabolic characteristics of carbon in the black soil parent material maturation process
ZHANG Zhiming, HAN Xiaozeng*
NationalObservationStationofHailunAgroecologySystem,NortheastInstituteofGeographyandAgroecology,ChineseAcademyofSciences,Harbin150081,China
The northeastern region of China represents the country′s largest commodity grain base. The Mollisols present in this northeastern region are very suitable for farming. However, the loss of top layer Mollisols and exposure of parent material have severely threatened the ecological environment and national food security in northeastern China. The soil microbial biochemistry process is the engine required for the reconstruction for soil fertilizers. Thus, it is important to characterize the pattern of carbon utilization by soil microbial communities during the parent maturation process to understand how microbial processes contribute to soil fertility. This study focuses on the carbon utilization pattern of soil microbial communities under different long-term maturing treatments at the national observation station of the Hailun Agroecology System. The results showed that Parent-Alfalfa (PAlf), Parent Corn-Soybean+Fertilizer+Biomass cycle (PCS+F+BM), and Parent Corn-Soybean+Fertilizer+Oganic Matter (PCS+F+OM) treatments contained the highest number of microorganisms. Parent-Natural (PNat), PAlf, PCS+F+BM, and PCS+F+OM treatments increased microbial activity to the level recorded under conventional farmland treatment. Different maturing process treatments produced different carbon utilization patterns in the microbial community. In all treatments, the highest utilization rate of the carbon source was carbohydrate. Principal component analysis indicated that the microbial metabolic characteristics under PNat, Parent-No Fertilizer (PCS-F), PCS+F+BM, and PCS+F+OM processes were similar to that those under Stander Soil Corn-Soybean+Fertilizer (SCS+F) treatment. We conclude that the change in microbial community structure under different maturing processes caused variations in the carbon utilization pattern, which ultimately influenced the metabolism characteristics of the overall carbon sources.
black soil parent material; maturing processes; microbial; metabolic characteristics
公益性行业(农业)科研专项经费资助(201203030, 201303126);国家自然科学基金资助(41371296)
2013- 12- 31;
日期:2015- 04- 14
10.5846/stxb201312313078
*通讯作者Corresponding author.E-mail: xzhan@neigaehrb.ac.cn
张志明,韩晓增.黑土母质熟化过程微生物群落碳源代谢特征.生态学报,2015,35(21):6957- 6964.
Zhang Z M, Han X Z.Microbial metabolic characteristics of carbon in the black soil parent material maturation process.Acta Ecologica Sinica,2015,35(21):6957- 6964.
猜你喜欢
中国化肥信息(2022年8期)2022-12-05
中国农业信息(2022年2期)2022-07-15
现代畜牧科技(2021年4期)2021-07-21
土壤学报(2019年6期)2020-01-15
农民致富之友(2019年33期)2019-12-20
浙江农业科学(2016年11期)2016-05-04
水土保持通报(2014年5期)2014-06-09
水土保持通报(2014年5期)2014-06-09
河北医科大学学报(2011年1期)2011-03-25
植物营养与肥料学报(2010年1期)2010-11-02
