
温带针阔混交林土壤碳氮气体通量的主控因子与耦合关系
2015-01-19 07:03党旭升程淑兰方华军于贵瑞韩士杰张军辉王永生徐敏杰李林森
生态学报 2015年19期
党旭升, 程淑兰, 方华军, 于贵瑞, 韩士杰, 张军辉, 王 淼, 王永生, 徐敏杰, 李林森, 王 磊
1 中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学, 北京 100049 3 中国科学院沈阳应用生态研究所, 沈阳 110016
温带针阔混交林土壤碳氮气体通量的主控因子与耦合关系
党旭升1,2, 程淑兰2, 方华军1,*, 于贵瑞1, 韩士杰3, 张军辉3, 王 淼3, 王永生1, 徐敏杰2, 李林森2, 王 磊1
1 中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学, 北京 100049 3 中国科学院沈阳应用生态研究所, 沈阳 110016
中高纬度森林地区由于气候条件变化剧烈,土壤温室气体排放量的估算存在很大的不确定性,并且不同碳氮气体通量的主控因子与耦合关系尚不明确。以长白山温带针阔混交林为研究对象,采用静态箱-气相色谱法连续4a(2005—2009年)测定土壤二氧化碳(CO2)、甲烷(CH4)和氧化亚氮(N2O)净交换通量以及温度、水分等相关环境因子。研究结果表明:温带针阔混交林土壤整体上表现为CO2和N2O的排放源和CH4的吸收汇。土壤CH4、CO2和N2O通量的年均值分别为-1.3 kg CH4hm-2a-1、15102.2 kg CO2hm-2a-1和6.13 kg N2O hm-2a-1。土壤CO2通量呈现明显的季节性规律,主要受土壤温度的影响,水分次之;土壤CH4通量的季节变化不明显,与土壤水分显著正相关;土壤N2O通量季节变化与土壤CO2通量相似,与土壤水分、温度显著正相关。土壤CO2通量和CH4通量不存在任何类型的耦合关系,与N2O通量也不存在耦合关系;土壤CH4和N2O通量之间表现为消长型耦合关系。这项研究显示温带针阔混交林土壤碳氮气体通量主要受环境因子驱动,不同气体通量产生与消耗之间存在复杂的耦合关系,下一步研究需要深入探讨环境变化对其耦合关系的影响以及内在的生物驱动机制。
土壤CO2通量; 土壤CH4通量; 土壤N2O通量; 主控因子; 耦合关系
气候变暖作为当前一个重要的环境问题,已备受关注。二氧化碳(CO2)、甲烷(CH4)和氧化亚氮(N2O)是导致全球气候变暖的3种最主要温室气体(GHGs)[1]。据统计,人类活动导致大气CO2、CH4和N2O浓度较工业革命前分别增加了35.4%、148%和18%,并以每年0.4%、0.8%和0.25%的速度递增,对温室效应的贡献分别为55%、19%和6%[2]。大气中GHGs浓度富集导致过去100年(1906—2005年)地表温度平均增加了0.74℃,预计在未来100年还将升高1.4—5.8℃[3]。虽然CH4和N2O浓度较小,但在百年尺度上,其辐射增温潜力分别是CO2的25和298倍,因此也必须给予足够的重视。大气中GHGs浓度的富集会改变陆地生态系统碳平衡和生态系统的可持续发展,因此研究陆地生态系统GHGs的吸收和排放通量及其影响因素,对于准确评价陆地生态系统对全球气候变化的贡献至关重要,同时也是制订GHGs减排措施的重要理论基础[4-5],在维持全球碳平衡方面具有重要的现实意义。
森林是陆地生态系统的主体,既是大气CO2、CH4、N2O等温室气体的源与汇[6],同时又是陆地最大的碳储存库和吸收汇[7]。因此,森林土壤碳库的微小变化都会对大气CO2、CH4、N2O浓度产生深远的影响[8],森林土壤碳氮气体交换通量对环境变化的响应是当前全球变化生态学的前沿领域之一。绿色植物通过光合作用合成有机物,植物枯死后凋落于土壤表面形成凋落层,凋落物和土壤有机碳被土壤微生物及动物利用,以CO2形式重新释放到大气中[9-11];同时植物根系的自养呼吸也会消耗有机碳释放CO2[9]。土壤CH4净交换通量是土壤剖面中CH4产生与氧化过程的平衡结果。在厌氧条件下,土壤腐生菌群落(如纤维分解菌、果胶分解菌和木质素分解菌)协同作用将有机质逐步降解成低分子有机酸,进而在甲烷产生菌作用下形成CH4[12-13],在好氧条件下,产生的CH4又会被甲烷氧化菌氧化为CO2。土壤N2O主要来源于硝化、反硝化和化学还原过程,其中硝化细菌和反硝化细菌作用下的生物源N2O占主导[14]。已有的研究结果表明,森林土壤CO2、CH4和N2O通量受多个因素的影响,包括温度、水分、森林类型、土壤有效碳氮含量、pH值以及人类干扰等[15-16]。在全球或区域尺度上,气候(降水和气温)是森林土壤碳氮气体通量的主要驱动因子,而在局域范围内,植被类型、土壤温度和土壤水分支配着森林土壤碳氮气体通量的季节与年际变异[17]。一般认为,水分充足时,温度是主要限制因子,而水分不足时,水分和温度共同影响森林土壤的温室气体排放[18]。此外,土壤CO2、CH4和N2O都是生物来源,在产生和消耗的过程中关键功能微生物群落之间存在复杂的交互作用,导致3种气体通量之间具有协同、随机和消长等耦合关系[16]。
温带森林主要分布在欧洲以及亚洲和北美洲的北部地区,占全球森林面积的18.3%,其碳储量约占森林总碳储量的10%[19]。我国温带森林占全国林地面积的31.4%[20],具有雨热同期以及对气候变化响应剧烈等特点,在区域森林碳氮循环和生态系统碳平衡研究中具有举足轻重的作用[21-22]。然而,由于缺乏长期的森林土壤温室气体观测数据,目前对东北地区温带森林土壤温室气体通量的数量、主控因子及其耦合关系尚未形成共识性的研究结论,导致区域尺度森林生态系统碳氮气体通量收支平衡计算、碳源汇效应评估以及对气候变化的响应与适应等方面存在很大的不确定性。基于上述研究的薄弱环节,本文以长白山温带针阔混交林为研究对象,基于多年的森林土壤温室气体连续监测数据,研究森林土壤碳氮气体净交换通量的季节变异特征、主要影响因素及其耦合关系,研究结果能够为完善现有的陆地生态系统碳氮耦合循环模型提供参数输入和模型验证,并为未来气候变化条件下温带针阔混交林温室气体排放控制以及降低区域森林生态系统碳汇评估的不确定性提供理论依据。
1 研究区域
研究地点位于中国科学院长白山森林生态系统研究站1号标准样地(42°24′ N,128°6′ E,海拔738 m),森林类型为阔叶红松林,林龄约200a。该区属温带大陆性山地气候,季节变化明显,5—9月为生长季。整个观测期间(2005—2009)年均温为3.6 ℃,年降雨量为600—800 mm (图1)。研究区地势平缓,平均坡度小于4°。阔叶红松林林分结构复杂,主要乔木为红松(Pinuskoraiensis)、紫椴(Tiliaamurensis)、蒙古栎(Quercusmongolica)、水曲柳(Fraxinusmandshurica)和色木槭(Acermono)。土壤为山地暗棕壤,母质为火山灰砂砾,土层厚度在70—100 cm;主要养分指标:有机碳11.45%, 全氮1.88%、全磷0.13%、全钾2.33%、C/N 6.09、速效氮933 mg/kg、速效磷9.96 mg/kg、速效钾381 mg/kg、pH 4.66[23]。
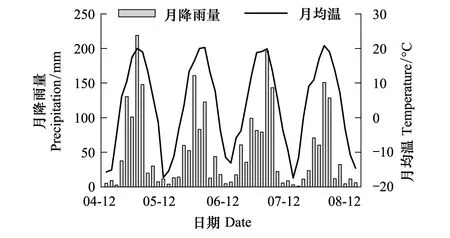
图1 2005年1月—2009年1月期间研究区气温和降雨量的季节和年际变异Fig.1 The seasonal and annual variability of air temperature and precipitation from 2004 to 2008
2 数据来源与研究方法
2.1 土壤温室气体通量监测
土壤GHGs净交换通量采用静态箱-气相色谱法测定。观测点设在通量塔附近200 m处,取样箱由底箱和盖箱组成, 其中盖箱为内径25 cm、高30 cm的不锈钢圆柱体;底箱高度为10 cm,其外径与盖箱的内径相同。采样频率至少每周1次,采样时间为9:00—11:00,代表土壤GHGs通量的日平均值。扣上盖箱,在30 min时段内每隔10min采集1次气样,存储于铝箔气袋中,运回实验室使用气相色谱仪(Agilent4890A,USA)测定CO2、CH4和N2O气体浓度。其中,CO2和CH4用氢离子化(FID)检测器测定,N2O用电子俘获(ECD)检测器测定。测定时气相色谱仪的主要参数:柱箱温度55℃,检测器FID和ECD的温度分别为250 ℃和330 ℃,载气(干空气及高纯H2)流量分别为300 mL/min和50 mL/min。通过气体浓度随时间的变化来计算单位面积的气体排放通量,线性相关系数(r2)大于0.9。计算公式如下:
(1)
式中,F为土壤CO2、N2O或CH4净交换通量(mg m-2h-1);M为气体的摩尔质量,V0为被测气体在标准状态下的摩尔体积,即22.41 L/mol;h,A和V分别是采样箱的高度(cm)、采样观测的土壤面积(cm2)和采样箱体积(cm3);P为观测点的大气压(kPa);T为箱内气体温度(oF);T0和P0分别是标准状态下被测气体的温度(273.15℉)和气压(101.325 kPa),dCt/dt为箱内碳氮气体浓度随时间变化的回归曲线斜率,正值表示排放,负值表示吸收。
2.2 辅助参数监测
利用水银温度计记录采样时段的开始和结束时刻气温,利用电子温度计(JM624,China)记录下箱内温度、地下5 cm温度。同时,在土壤非冻结期间,采用土壤水分仪(TDR200,USA)测定土壤10 cm土壤体积含水量,每个样方测定5点计算平均值做为某一时刻的土壤体积含水量。
2.3 数据统计分析
利用单因素方差分析评价月份对土壤碳氮气体通量以及温度、水分等环境因子的影响。利用线性和非线性回归方法拟合土壤碳氮气体通量与温度、水分之间的关系;利用一元回归分析方法探讨观测日和月两个时间尺度上土壤CO2、CH4和N2O通量之间的耦合关系。利用SPSS 16.0软件进行统计分析,利用Sigmaplot 10.0软件进行绘图。
3 结果与分析
3.1 土壤碳氮气体通量的季节变化与年际动态
基于2005年1月至2009年1月4年连续监测数据,研究发现长白山温带针阔混交林生态系统气温、地表温度和5cm土壤温度呈现单峰季节变化规律(图2,P<0.001),气温7月份最高(22.0oC),而地表和土壤温度则滞后一个月,峰值出现在8月,平均值分别为20.4oC和18.4℃(图2);气温最低值出现在12月份(-12.3℃),而且地表和5cm土壤温度最低值在1月份,均值分别为-7.5℃和-5.5℃(图2)。此外,不同年份之间3种温度的差异不明显(图2)。非冻结期间,土壤体积含水量最大值为5月份,平均为62.6%,此时土壤冻融作用强烈,温度低蒸发微弱;随后土壤含水量趋于下降,秋冬季节土壤含水量最低。
观测期间,温带针阔混交林土壤CH4排放与吸收交替出现,表现出强烈的时间异质性(图3,P<0.001)。总体而言,在2—3月份冬春交替季节,针阔混交林土壤表现出CH4排放,从4月份开始表现为吸收,直至翌年1月份,最大吸收值出现在10月份(图3)。土壤CH4月净交换通量变化范围为-0.093—0.007 mg CH4m-2h-1,年平均值为(-0.015±0.03) mg CH4m-2h-1(-1.3 kg CH4hm-2a-1)。土壤CO2排放呈现明显的单峰季节变化规律(P<0.001),最高值出现在8月份,与土壤温度变化格局一致(图3)。土壤CO2月排放通量变化范围为23.57—424.5 mg CO2m-2h-1,年平均值为(172.4±43.88) mg CO2m-2h-1(15102.2 kg CO2hm-2a-1)。土壤N2O通量也呈现显著的季节变化(P<0.001),随着温度的升高呈现增加的趋势,最大值也是出现在8月份,之后土壤N2O通量急剧下降(图3)。土壤N2O月排放通量变化范围为0.008—0.15 mg N2O m-2h-1,年平均值为(0.07±0.01) mg N2O m-2h-1(6.13 kg N2O hm-2a-1)。
3.2 土壤碳氮气体通量与温度、水分的关系
连续4a的观测数据显示,土壤CH4通量与5 cm土壤温度的相关性不显著,而与土壤含水量呈现极显著的正相关关系,土壤体积含水量能够解释土壤CH4变异的30%(图4)。土壤CO2通量与5 cm土壤温度显著正相关,两者的关系符合指数增长方程,土壤温度能够解释土壤CO2通量变异的68%(图4);此外,土壤CO2通量与土壤水分也显著正相关,但是水分只能解释土壤CO2通量变异的11%(图4)。值得关注的是,土壤N2O通量与土壤温度显著正相关,且两者之间也可用指数增长方程很好地拟合,土壤温度能够解释土壤N2O通量变异的16%(图4);土壤N2O通量与土壤水分也极显著正相关,土壤水分能够解释N2O通量变异的32%(图4)。上述研究结果表明,土壤CH4通量主要受水分驱动,温度影响较小。土壤CO2通量主要为土壤温度驱动,其次是水分;而土壤N2O通量主要受水分驱动,其次是温度。
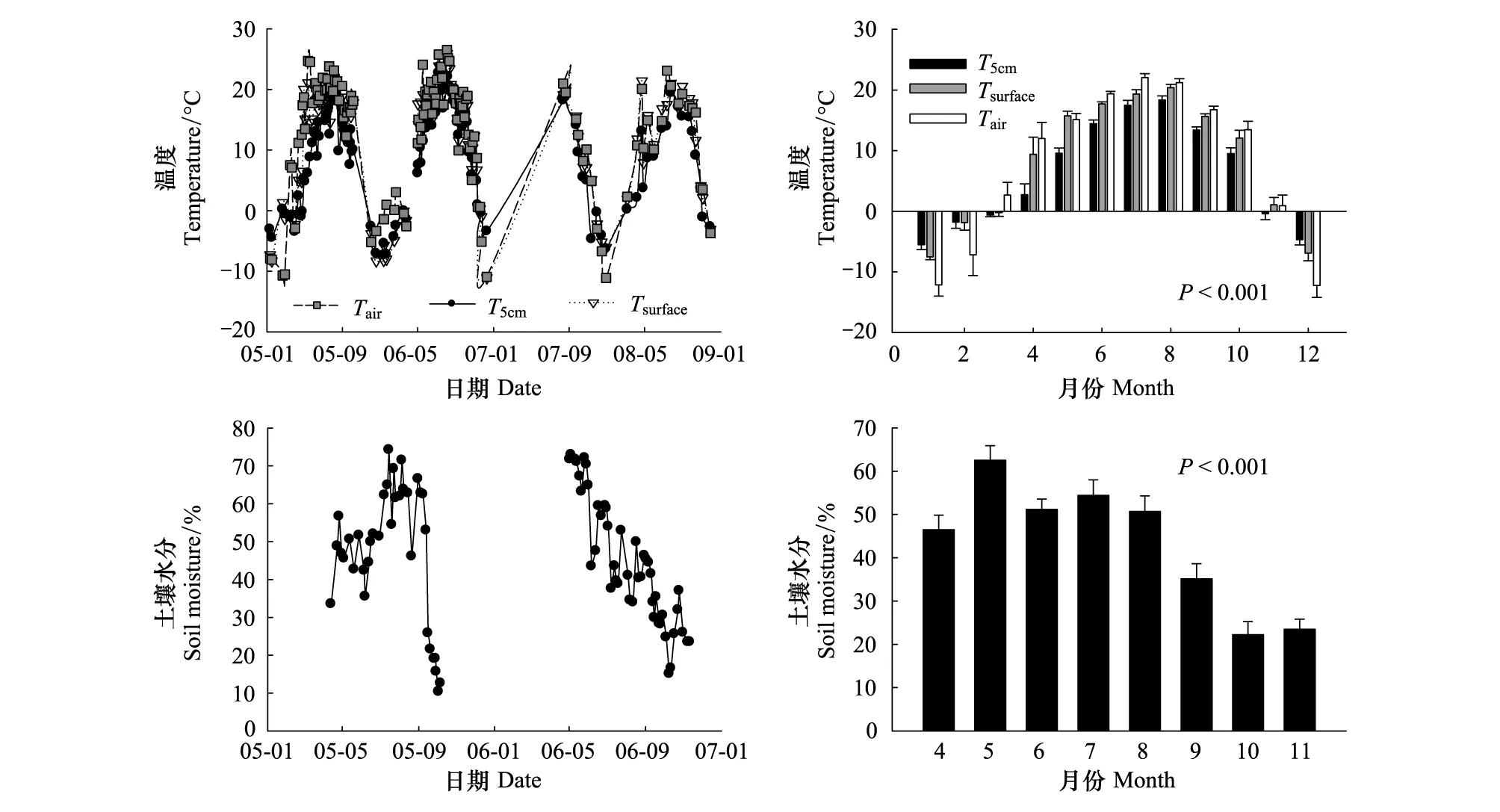
图2 气温、地表温度、5cm 土壤温度和10cm 土壤含水量的季节与年际变异特征Fig.2 The seasonal and annual variability of air temperature, surface temperature, soil temperature at the 5cm depth and soil moisture at the 10cm depth
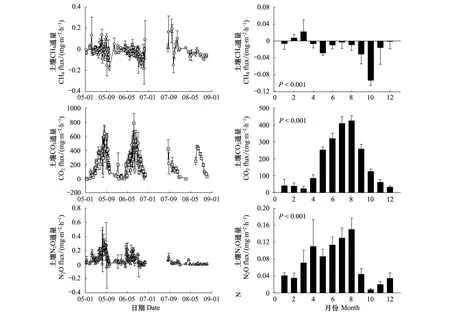
图3 土壤二氧化碳(CO2)、甲烷(CH4)和氧化亚氮(N2O)通量的季节与年际变异特征Fig.3 The seasonal and annual variability of soil carbon dioxide (CO2), methane (CH4), and nitrous oxide (N2O) fluxes
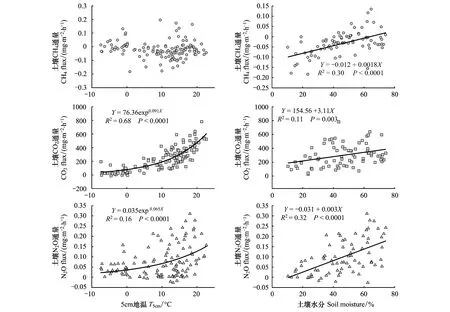
图4 土壤CO2, CH4和N2O通量与土壤温度、含水量之间的关系Fig.4 The relationships between soil CO2, CH4, N2O fluxes and soil temperature as well as soil moisture
3.3 土壤碳氮气体通量之间的耦合关系
基于观测日和月尺度的数据,研究发现土壤CO2通量与CH4通量之间无显著的相关关系,即两者表现为一种随机关系(图5)。在观测日尺度上,土壤CH4通量与土壤N2O通量之间正相关,但决定系数较小(图5),说明土壤CH4吸收与土壤N2O排放之间为弱消长型的耦合关系;在月尺度上,土壤CH4通量与N2O通量相关性不显著(图5)。在观测日和月尺度两个水平上,土壤CO2通量与N2O通量之间均呈现显著的正相关关系,两者对温度和水分有很强的同向响应(图5)。
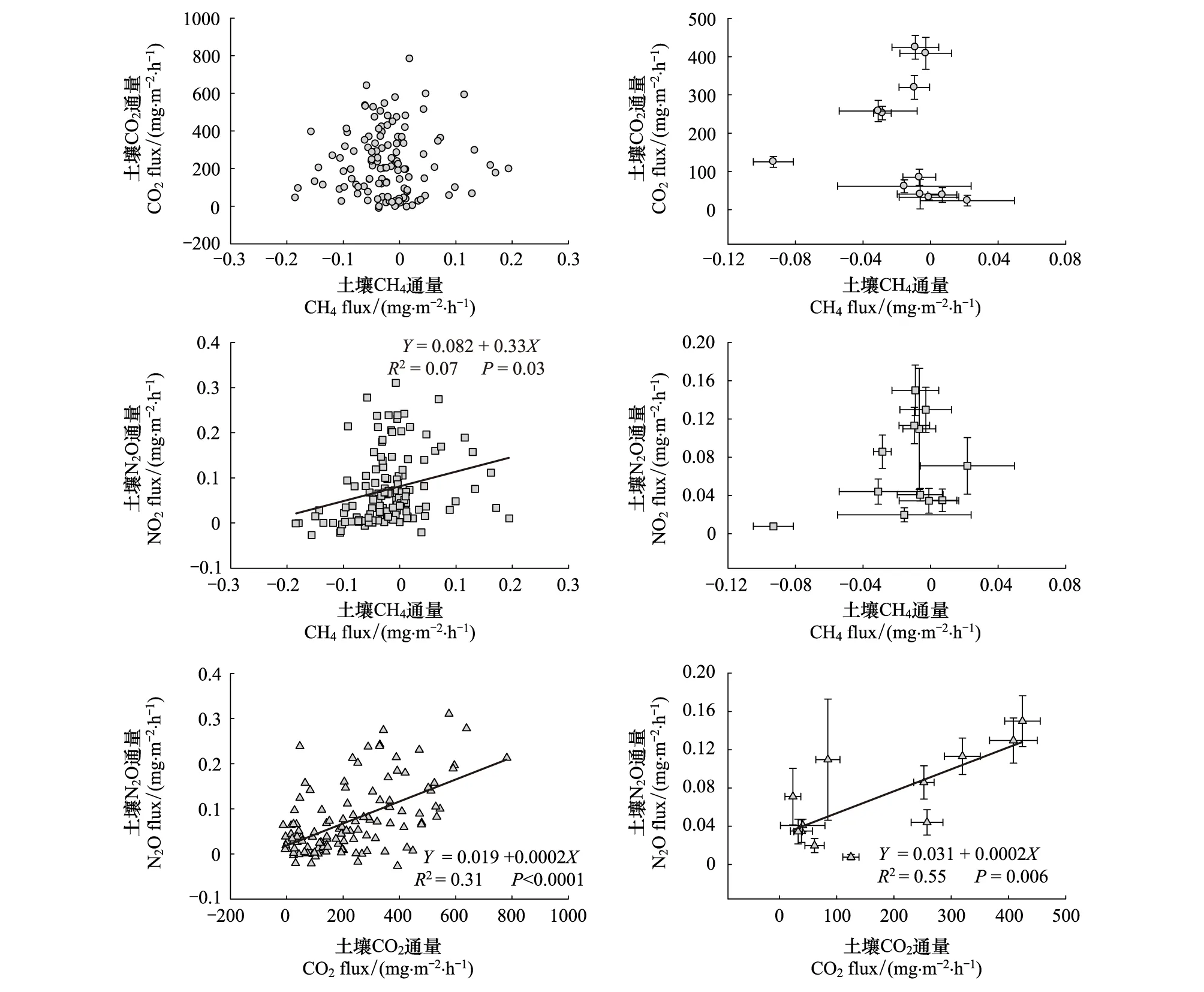
图5 土壤CO2, CH4和N2O通量之间的耦合关系Fig.5 The coupling relationships among soil CO2, CH4 and N2O fluxes
4 讨论
4.1 3种气体通量的季节变异特征和主控因子
土壤CO2排放通量呈现出夏高冬低的季节格局[18],与土壤温度的季节变化一致。土壤CO2排放通量除了化学氧化产生以外,主要是由植物根系自养呼吸和微生物异养呼吸产生。随着春季的到来,气温逐渐回升,土壤开始冻融,加上降雨频繁,植物根系代谢和土壤分解微生物群落活性增强,导致土壤自养和异养呼吸速率均增加;而进入秋季后,随着气温降低、降雨减少,植物代谢和微生物活性减弱,土壤呼吸速率降低[24-25]。另外,本研究表明温带针阔混交林土壤CO2通量主要受温度驱动,其次是土壤含水量,土壤CO2通量与温度的关系符合Arrhenius指数增长方程,这与许多室内培养及野外观测的结果相同[26-27]。在中高纬度的温带季风森林区,降水大于生态系统蒸散,土壤水分不是土壤CO2排放的主要限制性因子;由于温带针阔混交林土壤有机碳含量较高,活性组分比例较大,活化能较低,土壤CO2排放对温度反应敏感,所以土壤温度是支配土壤CO2释放的主要因子[28-29]。但是,在温带针阔混交林区,土壤温度只能解释土壤CO2排放通量变异的68%,低于一些样地尺度上的研究结论[30-32]。连续4a的监测结果表明,长白山温带阔叶混交林土壤CO2通量的变化范围为15.08—781.76 mg m-2h-1,与早期同地点的研究结果非常相近(14.4—720 mg m-2h-1)[33-35],明显高于北美温带森林土壤的CO2排放通量(10.0—234.0 mg m-2h-1)[36]和德国温带森林土壤CO2通量(10.8—540 mg m-2h-1)[37]。
温带针阔混交林生长季内土壤CH4平均吸收通量为15 μg m-2h-1,明显低于北京地区暖温带森林土壤CH4吸收通量(71 μg m-2h-1)[38],与不同气候带之间温度、水分、植被类型以及土壤类型等因素的差异有关。除了冬季冻结期外,温带针阔混交林土壤大部分时期表现为CH4吸收,这与大兴安岭寒温带针叶林的结果类似[39],体现了水分非饱和的自然土壤(如森林、草原等)都具有吸收大气CH4的能力,是大气CH4唯一的生物汇[1]。相似地,Groffman等[40]发现美国北部硬木林土壤结冻后有短暂的CH4释放现象,Sullivan 等[41]研究美国西南部黄松林地冬季CH4排放时未发现CH4有明显的吸收通量;相反Castro等[42]却报道美国马萨诸塞州红松林土壤在冬季始终表现为吸收CH4,这些不一致的研究结果体现了气候、森林类型、土壤属性的异质性。另外,本研究结果表明,温带针阔混交林土壤CH4通量与水分正相关,与温度相关性不显著(图4),其结果与以往的一些研究并不完全一致[43]。例如,Butterbach-Bahl等[44]研究表明,温带森林土壤CH4吸收通量会随着温度的升高而增加。普遍认为森林土壤CH4吸收通量会随湿度增加而降低[45-46],土壤CH4的吸收通量与温度、土壤含水量存在一定阈值关系[47],CH4净交换通量与土壤温度的关系符合二次方程,与土壤水分的关系成指数递减方程[48]。例如,Castro等[42]认为当土壤温度在-5—10 ℃时,温度为控制森林土壤CH4吸收的主导因子,当土壤温度在10℃以上时,土壤含水量则成为主导因子。在本研究中,2—3月份,土壤温度在-5—0℃之间,土壤冻结限制了大气O2向土壤剖面扩散,也限制了土壤CH4氧化菌群落的活性,同时厌氧条件下土壤剖面CH4累积,促进了土壤CH4产生菌群落活性,导致CH4氧化减弱排放增加[49]。而4月以后,随着温度升高,加上降水集中,土壤含水量较高,土壤CH4氧化吸收仍然不大,直到10月份,气候变得干燥降水减少,土壤含水量急剧下降,温度和水分达到最优配置,土壤CH4吸收强度最大。

4.2 3种气体通量的耦合关系
土壤CO2、CH4、N2O的产生与消耗过程之间存在复杂的交互作用,并受外界环境因子的影响。三者之间的耦合作用是以土壤微生物功能群为媒介,通过一系列的氧化还原反应完成电子传递、能量流动和物质转化,在不同生态系统、不同外界驱动力下表现出形式各异的耦合关系。土壤CO2、CH4和N2O通量的耦合关系常以他们之间的回归曲线的斜率来表征:协同关系(斜率大于零),消长关系(斜率小于零)或随机关系(斜率等于零)[16]。耦合关系不同于一般的相关关系,前者需要满足一定的条件,如存在交互作用的媒介(如甲烷氧化菌vs 氨氧化菌)、相同的底物(如有效碳氮)、相似的驱动过程(氧化还原反应、电子得失)等[16]。
通过对3种主要温室气体通量的研究分析,发现土壤CO2通量和CH4通量之间无显著的相关关系,机理上并不存在任何类型的耦合关系,其原因如下:首先,土壤CO2来源于植物根系的自养呼吸和微生物的异养呼吸,而土壤CH4通量取决于甲烷产生菌产生CH4和甲烷氧化菌氧化CH4两个过程的平衡,微生物使用的碳源基质存在很大的差异;其次,CO2通量和CH4通量的主要驱动因子不同,CO2通量主要受温度驱动,其次是水分,而CH4通量主要受水分驱动,与温度关系不显著,水热因子对不同微生物群落活性、对底物的利用程度的影响不同,导致CO2通量和CH4通量在观测日、月两个时间尺度上均表现为随机关系。土壤CH4和N2O通量的变化表现为消长型的耦合关系,其潜在的机理为:首先,土壤CH4和N2O产生的主要控制因子均为水分,即土壤通气性控制着两种气体在土壤剖面扩散,氧化还原反应控制着土壤CH4的产生与氧化以及土壤硝化和反硝化作用。在长白山阔叶红松林不同地点的土壤N2O排放和CH4吸收的原位观测中发现,N2O排放和CH4吸收间存在着非线性负相关关系,这种消长关系与土壤含水量的变化有关[67]。其次,从微生物生理和生态学角度上来看,土壤N2O是在氨氧化菌、硝化细菌和反硝化细菌作用下产生,CH4吸收是在甲烷氧化菌驱动下完成的生物化学过程;由于CH4和NH3分子量相近,土壤甲烷氧化菌和氨氧化菌均能同时氧化CH4和NH3,竞争利用相同的底物如O2、CH4和NH3;其次,甲烷氧化菌和氨氧化菌都具有非常复杂的胞质内膜,在氧化CH4和NH3的过程中会竞争利用功能相似的单氧酶,尤其是甲烷单氧酶(MMO)[68-69]。然而,也有少数研究显示,N2O排放速率和CH4吸收速率间呈显著的正相关关系[70],与生态系统的氮饱和状态有关。土壤CO2和N2O通量对温度和水分有很强的同向响应,归因于两者的驱动因子相同,均为温度和水分共同驱动,但从机理上来说,二者并不存在耦合。本研究只是探讨了土壤CO2, CH4和N2O通量的水热驱动作用,其它环境因子和关键功能微生物群落动态对3种通量耦合关系的影响有待更为深入的研究。
5 结论
(1)土壤CO2和N2O交换通量表现为明显的单峰季节变化,而土壤CH4排放与吸收交替出现,表现出强烈的时间异质性。土壤CO2和N2O主要受土壤温度和水分影响,而土壤CH4主要受土壤水分驱动。
(2)土壤CO2通量和CH4通量之间表现为随机关系,可能与两者的底物利用不同有关;CH4吸收和N2O排放通量表现为消长关系,归因于水分控制以及氨氧化菌与甲烷氧化菌之间的竞争作用;而CO2和N2O通量之间具有很强的协同关系,与它们对温度和水分的同向响应有关。
[1] Garcia-Gonzalo J, Peltola H, Briceno-Elizondo E, Kellomaki S. Changed thinning regimes may increase carbon stock under climate change: A case study from a Finnish boreal forest. Climatic Change, 2007, 81(3/4): 431-454.
[2] Lelieveld J, Crutzen P J. Indirect chemical effects of methane on climate warming. Nature, 1992, 355(6358): 339-342.
[3] Solomon S. Climate Change 2007-The Physical Science Basis: Working group I Contribution to the Fourth Assessment Report of the IPCC. Cambridge: Cambridge University Press, 2007.
[4] Sun D M, Zhang L, Xue M S. The online measurements and estimation of nutrient solution in greenhouse agriculture // Proceedings of the 3rd World Congress on Intelligent Control and Automation. Hefei: IEEE, 2000: 2155-2157.
[5] Butterbach-Bahl K, Rothe A, Papen H. Effect of tree distance on N2O and CH4-fluxes from soils in temperate forest ecosystems. Plant and Soil, 2002, 240(1): 91-103.
[6] Bowden R D, Davidson E, Savage K, Arabia C, Steudler P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. Forest Ecology and Management, 2004, 196(1): 43-56.
[7] 刘世荣, 王晖, 栾军伟. 中国森林土壤碳储量与土壤碳过程研究进展. 生态学报, 2011, 31(19): 5437-5448.
[8] Dixon R K, Brown S, Houghton R A, Solomon A M, Trexier M C, Wisniewski J. Carbon pools and flux of global forest ecosystems. Science, 1994, 263(5144): 185-190.
[9] 齐玉春, 董云社, 耿元波, 杨小红, 耿会立. 我国草地生态系统碳循环研究进展. 地理科学进展, 2003, 22(4): 342-352.
[10] 谢军飞, 李玉娥. 农田土壤温室气体排放机理与影响因素研究进展. 中国农业气象, 2002, 23(4): 48-53.
[11] 李长生, 肖向明, Frolking S, Moore B Ⅲ, Salas W, 邱建军, 张宇, 庄亚辉, 王效科, 戴昭华, 刘纪远, 秦小光, 廖柏寒, Sass R. 中国农田的温室气体排放. 第四纪研究, 2003, 23(5): 493-503.
[12] 王跃思, 薛敏, 黄耀, 刘广仁, 王明星, 纪宝明. 内蒙古天然与放牧草原温室气体排放研究. 应用生态学报, 2003, 14(3): 372-376.
[13] 冯虎元, 程国栋, 安黎哲. 微生物介导的土壤甲烷循环及全球变化研究. 冰川冻土, 2004, 26(4): 411-419.
[14] 李海防, 夏汉平, 熊燕梅, 张杏锋. 土壤温室气体产生与排放影响因素研究进展. 生态环境, 2007, 16(6): 1781-1788.
[15] 程淑兰, 方华军, 于贵瑞, 徐敏杰, 张裴雷, 郑娇娇, 高文龙, 王永生. 森林土壤甲烷吸收的主控因子及其对增氮的响应研究进展. 生态学报, 2012, 32(15): 4914-4923.
[16] 方华军, 程淑兰, 于贵瑞, 王永生, 徐敏杰, 党旭升, 李林森, 王磊. 大气氮沉降对森林土壤甲烷吸收和氧化亚氮排放的影响及其微生物学机制. 生态学报, 2014, 34(17), doi: 10. 5846/stxb201310262582.
[17] Smith K A, Ball T, Conen F, Dobbie K E, Massheder J, Rey A. Exchange of greenhouse gases between soil and atmosphere: interactions of soil physical factors and biological processes. European Journal of Soil Science, 2003, 54(4): 779-791.
[18] 孙向阳, 乔杰, 谭笑. 温带森林土壤中的CO2排放通量. 东北林业大学学报, 2001, 29(1): 34-39.
[19] Lal R. Forest soils and carbon sequestration. Forest Ecology and Management, 2005, 220(1/3): 242-258.
[20] 中华人民共和国林业部. 全国森林资源统计(1989-1993). 北京: 国家林业部, 1994.
[21] 牟艳玲, 赵文龙, 陈亚雄, 贾超杰, 孙国钧. 中国北方森林潜在分布及其对气候变化响应的模拟. 兰州大学学报: 自然科学版, 2010, 46(S1): 25-32.
[22] 栾兆平. 气候变化与中国北方森林恢复和经营. 内蒙古林业调查设计, 2007, 30(5): 47-49, 74-74.
[23] Cheng S L, Fang H J, Yu G R, Zhu T H, Zheng J J. Foliar and soil15N natural abundances provide field evidence on nitrogen dynamics in temperate and boreal forest ecosystems. Plant and Soil, 2010, 337(1/2): 285-297.
[24] 张慧东, 周梅, 赵鹏武, 包青春, 海龙. 寒温带兴安落叶松林土壤呼吸特征. 林业科学, 2008, 44(9): 142-145.
[25] 周丽艳, 贾丙瑞, 周广胜, 曾伟, 王宇. 中国北方针叶林生长季碳交换及其调控机制. 应用生态学报, 2010, 21(10): 2449-2456.
[26] 周海霞, 张彦东, 孙海龙, 吴世义. 东北温带次生林与落叶松人工林的土壤呼吸. 应用生态学报, 2007, 18(12): 2668-2674.
[27] 陈宝玉, 刘世荣, 葛剑平, 王辉, 常建国, 孙甜甜, 马姜明, 施恭暕. 川西亚高山针叶林土壤呼吸速率与不同土层温度的关系. 应用生态学报, 2007, 18(6): 1219-1224.
[28] Crill P M. Seasonal patterns of methane uptake and carbon dioxide release by a temperate woodland soil. Global Biogeochemical Cycles, 1991, 5(4): 319-334.
[29] Davidson E A, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology, 1998, 4(2): 217-227.
[30] 常建国, 刘世荣, 史作民, 康冰, 陈宝玉. 锐齿栎林土壤呼吸对土壤水热变化的响应. 林业科学, 2006, 42(12): 21-27.
[31] 陈全胜, 李凌浩, 韩兴国, 阎志丹, 王艳芬, 袁志友. 水热条件对锡林河流域典型草原退化群落土壤呼吸的影响. 植物生态学报, 2003, 27(2): 202-209.
[32] 常宗强, 史作民, 冯起, 苏永红. 黑河流域山区牧坡草地土壤呼吸的时间变化及水热因子影响. 应用生态学报, 2005, 16(9): 1603-1606.
[33] Vogt K A, Edmonds R L, Antos G C, Vogt D J. Relationships between CO2evolution, ATP concentrations and decomposition in four forest ecosystems in western Washington. Oikos, 1980, 35: 72-79.
[34] Robert E S, Keith V C. Relationships between CO2evolution from soil, substrate-temperature, and substrate moisture in four mature forest types in interior Alaska. Canadian Journal of Forest Research, 1985, 15(1): 97-106.
[35] Pajari B. Soil respiration in a poor upland site of scots pine stand subjected to elevated temperatures and atmospheric carbon concentration. Plant and Soil, 1995, 168-169(1): 563-570.
[36] Singh J S, Gupta S R. Plant decomposition and soil respiration in terrestrial ecosystems. The botanical Review, 1977, 43(4): 449-528.
[37] 董云社, 彭公炳, 李俊. 温带森林土壤排放CO2、CH4、N2O时空特征. 地理学报, 1996, 51(S1): 120-128.
[38] 杜睿, 黄建辉, 万小伟, 贾月慧. 北京地区暖温带森林土壤温室气体排放规律. 环境科学, 2004, 25(2): 12-16.
[39] Fang H J, Yu G R, Cheng S L, Zhu T H, Wang Y S, Yan J H, Wang M, Cao M, Zhou M. Effects of multiple environmental factors on CO2emission and CH4uptake from old-growth forest soils. Biogeosciences, 2010, 7(1): 395-407.
[40] Groffman P M, Hardy J P, Driscoll C T, Fahey T J. Snow depth, soil freezing, and fluxes of carbon dioxide, nitrous oxide and methane in a northern hardwood forest. Global Change Biology, 2006, 12(9): 1748-1760.
[41] Sullivan B W, Kolb T E, Hart S C, Kaye J P, Dore S, Montes-Helu M. Thinning reduces soil carbon dioxide but not methane flux from southwestern USA ponderosa pine forests. Forest Ecology and Management, 2008, 255(12): 4047-4055.
[42] Castro M S, Steudler P A, Melillo J M, Aber J D, Bowden R D. Factors controlling atmospheric methane consumption by temperate forest soils. Global Biogeochemical Cycles, 1995, 9(1): 1-10.
[43] 刘实, 王传宽, 许飞.4种温带森林非生长季土壤二氧化碳、甲烷和氧化亚氮通量. 生态学报, 2010, 30(15): 4075-4084.
[44] Butterbach-Bahl K, Gasche R, Huber C, Kreutzer K, Papen H. Impact of N-input by wet deposition on N-trace gas fluxes and CH4-oxidation in spruce forest ecosystems of the temperate zone in Europe. Atmospheric Environment, 1998, 32(3): 559-564.
[45] Castro M S, Gholz H L, Clark K L, Steudler P A. Effects of forest harvesting on soil methane fluxes in Florida slash pine plantations. Canadian Journal of Forest Research, 2000, 30(10): 1534-1542.
[46] Verchot L V, Davidson E A, Cattanio J H, Ackerman I L. Land-use change and biogeochemical controls of methane fluxes in soils of eastern Amazonia. Ecosystems, 2000, 3(1): 41-56.
[47] Heyer J. Results of enrichment experiments of methane-assimilating organisms from an ecological point of view. Microbial growth on C1 compounds. USSR Academy of Sciences, 1977: 19-21.
[48] Xu M J, Cheng S L, Fang H J, Yu G R, Gao W L, Wang Y S, Dang X S, Li L S. Low-Level nitrogen addition promotes net methane uptake in a boreal forest across the great Xing′an Mountain region, China. Forest Science, 2014, 60(2), doi: 10.5849/forsci.13-075.
[49] 丁维新, 蔡祖聪. 温度对土壤氧化大气CH4的影响. 生态学杂志, 2003, 22(3): 54-58.
[50] 傅民杰, 王传宽, 王颖, 刘实, 丁爽. 气候暖化对解冻期不同纬度兴安落叶松林土壤氧化亚氮释放的影响. 应用生态学报, 2009, 20(7): 1635-1642.
[51] 肖冬梅, 王淼, 姬兰柱, 韩士杰, 王跃思. 长白山阔叶红松林土壤N2O排放通量的变化特征. 生态学杂志, 2004, 23(5): 46-52.
[52] 胡承彪, 韦源连, 梁宏温, 朱宏光. 两种森林凋落物分解及其土壤效应的研究. 广西农业大学学报, 1992, 11(4): 47-52.
[53] 王鹤松, 张劲松, 孟平, 高峻, 贾长荣, 任迎丰. 侧柏人工林地土壤呼吸及其影响因子的研究. 土壤通报, 2009, 40(5): 1031-1035.
[54] 齐玉春, 董云社. 土壤氧化亚氮产生、排放及其影响因素. 地理学报, 1999, 54(6): 534-542.
[55] 谢军飞, 李玉娥. 土壤温度对北京旱地农田N2O排放的影响. 中国农业气象, 2005, 26(1): 7-10.
[56] Teepe R, Vor A, Beese F, Ludwig B. Emissions of N2O from soils during cycles of freezing and thawing and the effects of soil water, texture and duration of freezing. European Journal of Soil Science, 2004, 55(2): 357-365.
[57] Mummey D L, Smith J L, Bolton H J. Nitrous oxide flux from a shrub-steppe ecosystem: sources and regulation. Soil Biology & Biochemistry, 1994, 26(2): 279-286.
[58] Teepe R, Brumme R, Beese F. Nitrous oxide emissions from frozen soils under agricultural, fallow and forest land. Soil Biology & Biochemistry, 2000, 32(11-12): 1807-1810.
[59] 孙向阳, 徐化成. 北京低山区两种人工林土壤中N2O排放通量的研究. 林业科学, 2001, 37(5): 57-63.
[60] Riley R H, Vitousek P M. Nutrient dynamics and nitrogen trace gas flux during ecosystem development in montane rain forest. Ecology, 1995, 76(1): 292-304.
[61] Kiese R, Butterbach-Bahl K. N2O and CO2emissions from three different tropical forest sites in the wet tropics of Queensland, Australia. Soil Biology & Biochemistry, 2002, 34(7): 975-987.
[62] Singurindy O, Molodovskaya M, Richards B K, Steenhuis T S. Nitrous oxide emission at low temperatures from manure-amended soils under corn (ZeamaysL.). Agriculture Ecosystems & Environment, 2009, 132(1): 74-81.
[63] Papen H, Butterbach-Bahl K. A 3-year continuous record of nitrogen trace gas fluxes from untreated and limed soil of a N-saturated spruce and beech forest ecosystem in Germany: 1. N2O emissions. Journal of Geophysical Research-Atmospheres (1984-2012), 1999, 104(D15): 18487-18503.
[64] Muller C, Martin M, Stevens R J, Laughlin R J, Kammann C, Ottow J C G, Jager H J. Processes leading to N2O emissions in grassland soil during freezing and thawing. Soil Biology & Biochemistry, 2002, 34(9): 1325-1331.
[65] Teepe R, Brumme R, Beese F. Nitrous oxide emissions from soil during freezing and thawing periods. Soil Biology & Biochemistry, 2001, 33(9): 1269-1275.
[66] Oquist M G, Nilsson M, Sorensson F, Kasimir-Klemedtsson A, Persson T, Weslien P, Klemedtsson L. Nitrous oxide production in a forest soil at low temperatures-processes and environmental controls. FEMS Microbiology Ecology, 2004, 49(3): 371-378.
[67] Mosier A, Schimel D, Valentine D, Bronson K, Parton W. Methane and Nitrous-Oxide fluxes in native, fertilized and cultivated grasslands. Nature, 1991, 350(6316): 330-332.
[69] Sitaula B K, Bakken L R. Nitrous oxide release from spruce forest soil: Relationships with nitrification, methane uptake, temperature, moisture and fertilization. Soil Biology & Biochemistry, 1993, 25(10): 1415-1421.
[70] 张秀君, 徐慧, 陈冠雄. 影响森林土壤N2O排放和CH4吸收的主要因素. 环境科学, 2002, 23(5): 8-12.
The controlling factors and coupling of soil CO2, CH4and N2O fluxes in a temperate needle-broadleaved mixed forest
DANG Xusheng1,2, CHENG Shulan2, FANG Huajun1,*, YU Guirui1, HAN Shijie3, ZHANG Junhui3, WANG Miao3, WANG Yongsheng1, XU Minjie2, LI Linsen2, WANG Lei1
1KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicalSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China2UniversityofChineseAcademyofSciences,Beijing100049,China3ShenyangInstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China
Carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O) are three major greenhouse gases, accounting for 80% of global warming. Forest ecosystems comprise the largest carbon stocks in terrestrial ecosystems, and function as important sinks and sources of atmospheric CO2, CH4and N2O. Complicated interactions occur during the generation and absorption of soil CO2, CH4and N2O, including synergies, tradeoffs, and randomness. High-latitude forests are experiencing the effects of significant global change (e.g., warming, changed precipitation, and increased nitrogen deposition), leading to great uncertainty in estimates of soil greenhouse gas fluxes. Furthermore, the factors controlling the coupling of soil CO2, CH4and N2O fluxes remain unclear. This study was conducted in the temperate needle-broadleaved mixed forest of Changbai Mountain, Northeast China. The net exchange fluxes of soil CO2, CH4and N2O, as well as soil temperature and soil moisture, were measured over four years (2005-2009) using static chamber and gas chromatograph techniques. The results showed that temperate needle-broadleaved mixed forest soils behaved as a source of atmospheric CO2and N2O but a sink of atmospheric CH4over the course of the study. The average soil CH4, CO2and N2O fluxes were estimated at -1.30 kg CH4hm-2a-1, 15102.2 kg CO2hm-2a-1, and 6.13 kg N2O hm-2a-1, respectively. In addition, soil CO2flux exhibited significant seasonality, and was mainly affected by soil temperature, followed by soil moisture. Seasonal variation in soil CH4flux was less significant than that of soil CO2and N2O fluxes; moreover, it was positively correlated with soil moisture. When soil temperatures were within a threshold range, soil moisture determined CH4production and oxidation in soil profiles by regulating CH4and O2diffusion as well as methanotrophic community activity. Similar to soil CO2flux, soil N2O flux was significantly correlated with soil moisture and soil temperature. Furthermore, there were no significant relationships between soil CO2flux and soil CH4flux, or between soil CO2flux and soil N2O flux, which exhibited a random relationship. However, a significant negative relationship between soil CH4uptake and N2O emission was found, indicating a tradeoff between them. The random relationship between soil CO2and CH4fluxes was attributed to their different pathways and substrate utilization. The trade-off relationship between soil CH4and N2O fluxes was related to moisture control and competition for mono-oxygenase (MMO) between ammonia-oxidizers and methanotrophic communities. The weak synergy between soil CO2and N2O fluxes reflected that they were driven by same environmental factors, such as soil temperature and moisture, and that no microbial mechanisms drove their production or consumption.These results suggest that soil carbon and nitrogen gas fluxes are mainly driven by environmental factors and substrate availability, and their complicated couplings are related to the activity and functional composition of microbial communities. It is necessary to further explore the effects of environmental change on the coupling of soil CO2, CH4and N2O fluxes as well as the microbial mechanisms underlying these using molecular biology and metagenomic analyses.
soil CO2flux; soil CH4flux; soil N2O flux; controlling factors; coupling
国家自然科学基金(31290221, 41471212, 31470558, 31290222, 31130009); 国家重点基础研究发展计划项目(2012CB417103); 中国科学院地理科学与资源研究所“秉维”优秀青年人才基金(2011RC202); 中国科学院战略性先导科技专项(XDA05050600)
2014-03-22; < class="emphasis_bold">网络出版日期:
日期:2014-12-04
10.5846/stxb201403220513
*通讯作者Corresponding author.E-mail: fanghj@igsnrr.ac.cn
党旭升, 程淑兰, 方华军, 于贵瑞, 韩士杰, 张军辉, 王淼, 王永生, 徐敏杰, 李林森, 王磊.温带针阔混交林土壤碳氮气体通量的主控因子与耦合关系.生态学报,2015,35(19):6530-6540.
Dang X S, Cheng S L, Fang H J, Yu G R, Han S J, Zhang J H, Wang M, Wang Y S, Xu M J, Li L S, Wang L.The controlling factors and coupling of soil CO2, CH4and N2O fluxes in a temperate needle-broadleaved mixed forest.Acta Ecologica Sinica,2015,35(19):6530-6540.
猜你喜欢
疯狂英语·新悦读(2022年7期)2022-11-23
辽宁林业科技(2017年4期)2017-06-22
浙江大学学报(工学版)(2016年2期)2016-06-05
中国环境科学(2016年4期)2016-05-27
安徽农业科学(2015年30期)2015-12-25
大气科学(2015年5期)2015-12-04
中国粮油学报(2015年5期)2015-02-06
亚热带资源与环境学报(2015年4期)2015-01-22
东北师大学报(自然科学版)(2014年1期)2014-02-27
亚热带资源与环境学报(2014年3期)2014-01-23
