
不同有机类肥料对小麦和水稻根际土壤线虫的影响
2015-01-19 07:06叶成龙陈小云沈其荣李辉信
生态学报 2015年19期
刘 婷, 叶成龙, 李 勇, 陈小云, 冉 炜, 沈其荣, 胡 锋, 李辉信,*
1 南京农业大学资源与环境科学学院, 南京 210095 2 江苏省有机固体废弃物资源化协同创新中心, 南京 210095 3 江苏省金坛市土壤肥料技术指导站, 金坛 213200
不同有机类肥料对小麦和水稻根际土壤线虫的影响
刘 婷1,2, 叶成龙1, 李 勇3, 陈小云1, 冉 炜1, 沈其荣1, 胡 锋1, 李辉信1,2,*
1 南京农业大学资源与环境科学学院, 南京 210095 2 江苏省有机固体废弃物资源化协同创新中心, 南京 210095 3 江苏省金坛市土壤肥料技术指导站, 金坛 213200
国内外对土壤线虫的研究绝大部分都是基于旱地的非根际土壤,对水田土壤,尤其是水旱轮作下根际土壤线虫对有机肥的响应研究非常地有限。因此,探讨在稻麦轮作体系中以猪粪堆肥和秸秆为原料的不同有机类肥料,对水稻和小麦根际土壤线虫的影响。田间试验处理为:CK(不施肥)、F(100%化肥)、PF(猪粪堆肥+50%化肥)、SF(秸秆全量还田+100%化肥)、PSF(猪粪堆肥+秸秆全量还田+50%化肥)、PMF(猪粪商品有机无机复合肥)。结果表明:小麦根际主要线虫优势属为头叶属(Cephalobus)、拟丽突属(Acrobeloides)和丝尾垫刃属(Filenchus);水稻根际则为杆咽属(Rhabdolaimus)、潜根属(Hirschmanniella)和丝尾垫刃属(Filenchus)。小麦根际土壤线虫总数显著高于水稻根际;SF和PSF对两季土壤线虫总数的提高显著高于PF和PMF,F对线虫总数没有显著影响。线虫营养类群表现为小麦根际食真菌线虫相对丰度显著高于水稻根际,植食性线虫和杂食/捕食性线虫丰度显著低于水稻根际。有机肥对稻季线虫群落的影响远大于麦季,秸秆配施有机肥显著提高麦季瓦斯乐斯卡指数(WI);施肥均显著提高稻季的多样性指数(H′),降低优势度指数(λ)。土壤线虫对肥料的响应取决于作物及肥料的质量和养分含量;在稻麦轮作系统下,同时添加秸秆和猪粪对提高土壤线虫数量和食微线虫丰度更有益。
根际; 有机无机配施; 稻麦轮作体系
大量实践表明,化肥虽能快速提高作物产量,但长期的化肥施用会对土壤结构、土壤肥力和生物多样性等产生严重影响[1-3]。有机肥有助于改善土壤结构,提高土壤肥力和促进微生物活性[4-5]。因此,有机肥配施化肥不仅能够满足植物养分的需求,还可以逐步提升耕地质量,培肥地力。
作为土壤生物区系的重要组成部分之一,线虫因具有身体透明易于鉴别、结构与功能对应关系好等特点,而越来越多地被作为土壤指示生物来加以应用,尤其是用于评价生态系统的土壤健康水平、生态系统演替或受干扰的程度[6]。因此,线虫可以作为一种具有代表性的土壤动物来研究不同施肥措施对土壤食物网产生的影响。Yeates等[7]将线虫划分为8个营养类群,其中,食细菌线虫、食真菌线虫、植食性线虫和杂食/捕食性线虫是农田生态系统中的主要营养类群[8]。
国内外绝大部分研究施肥对线虫群落结构的影响都集中于旱地土壤,我国也主要以研究北方旱旱轮作体系为主[1,9-11]。其中,大部分的实验表明有机肥的施用能够增加土壤线虫总数[1,9,11-12],提高土壤食细菌线虫、食真菌线虫[1,9,13-14]和杂食捕食性线虫的数量[1,13,15],降低植食性线虫的数量[1,13,15],从而对土壤微生物区系健康环境的构建有促进作用。然而,在本研究中,由于水田和旱田在根系、水分、pH等诸多方面存在明显差异,轮作又导致土壤系统季节间相互作用、相互影响,使得稻麦轮作成为一个独特的农田生态系统[16]。因此,在这种条件下有机肥的施用会对麦季和稻季的土壤线虫群落产生怎样的影响尚不明确。同时,由于根际是一个微生物密集的区域,是土壤、植物根系、微生物和土壤动物交换的活跃界面[17],研究根际的线虫群落结构更有助于反映植物生长微域的健康水平。因此,本文研究的目的是探讨稻麦轮作体系下麦季和稻季的根际土壤线虫群落结构的差异,及研究以秸秆和猪粪与不同量的化肥配施对线虫群落结构的影响。本研究有助于比较不同种类的有机肥及其与不同量的无机肥配施,对稻季和麦季的土壤肥力及线虫群落结构的调控差异,从而为有效选择和利用有机类肥料调控和构建一个健康的生物区系,提升水旱轮作下的耕地质量提供相关参考。
1 材料与方法
1.1 试验地概况
试验点位于江苏省金坛市指前镇建春村(N 31°39′41.8″,E 119°28′23.5″),海拔高度10m,属亚热带湿润季风性气候,年均气温15.5℃,年均湿度78%,年降雨量1084.7mm。
田间试验始于2010年11月小麦季,为夏水稻—冬小麦的水旱轮作体系。采样时间均为小麦和水稻种植两年后的收获期。实验小区共24个(6个处理×4次重复),每小区40m2(8m×5m),随机区组排列,小区间水泥埂隔离,防止串水串肥。供试土壤为脱潜型水稻土(乌栅土);供试水稻品种为武运粳23;供试小麦品种为扬辐麦4号。
试验设计具体施肥措施:① CK,完全不施肥;② F,纯NPK (100% 当地用量,下同);③ PF +50%NPK;④ SF,秸秆全量还田+100%NPK;⑤ PSF,猪粪堆肥 (6 t/hm2)+秸秆全量还田+50%NPK;⑥ PMF,猪粪商品有机无机复合肥 (3.6 t/hm2)。其中,纯NPK (尿素、过磷酸钙、硫酸钾) 肥料用量为:N 300 kghm2,P2O5120 kghm2,K2O 100 kghm2;猪粪堆肥养分含量:有机质45.4%,N 2.3%,P2O52.9%,K2O 1.2%,含水量29.1%;猪粪商品有机无机复合肥养分含量:有机质16.1%,N 12.2%,P2O54.1%,K2O 4.1%,含水量19.3%。
1.2 研究方法
采样分别在小麦(2012年6月)和水稻(2012年11月)收割时进行。小麦根际土壤用抖根法[18]采集;本实验中将土钻贴根采集的土壤近似认为水稻根际土,采样取土深度为0—10 cm,每小区随机选取10点采样。将采好的土样迅速装入自封袋编号,带回实验室作简单处理后置于4℃冰箱保存。
称取土壤20g,采用浅盘法分离土壤中的线虫[19],解剖镜下计算数量,然后随机抽取150条左右在光学显微镜下鉴定到属。线虫鉴定参考《中国土壤动物检索图鉴》[20]及Bongers主编的《DE NEMATODEN VAN NEDERLAND》[21]。将线虫分为4个营养类群:食细菌线虫(Bacterivores)、食真菌线虫(Fungivores)、植食性线虫(Plant-parasites)和杂食/捕食性线虫(Omnivores/ predators)[8]。
采用多种生态指数对土壤线虫的多样性和群落结构进行评价,具体计算方法如下:
(1)自由生活线虫成熟指数(MI)[22]
MI=∑c-pi·pi(仅包括自由生活线虫)
式中,pi为第i个分类单元中个体占线虫总个体数量的比例;
(2)植食性线虫成熟指数(PPI)[22]
PPI=∑c-pi·pi(仅包括植物寄生线虫)
(3)瓦斯乐斯卡指数(WI)[23]
WI=(BF+FF)/PP
式中,BF为食细菌线虫的数量;FF为食真菌线虫的数量;PP为植食性线虫的数量;
(4)线虫通路比值(NCR)[24]
NCR=BF/(BF+FF)
(5)香农多样性指数(H′)[23]
H′=-∑pi(lnpi)
(6)优势度指数(λ)[25]
式中,c-pi为赋予某一种类线虫的cp(colonizer-persister)值,cp值是Bongers[23]根据线虫不同的生活史策略,将陆地和淡水生活的线虫划分为r-对策者(r-strategies,世代时间短,产卵量大,耐环境压力,cp值小)向k-对策者(k-strategies,世代时间长,产卵量小,对环境压力敏感,cp值大)过渡的5个类群。
1.3 统计分析
土壤线虫数量折算成每100g干土含有线虫的条数;丰度以不同属或营养类群的线虫数量占线虫总数的百分比(%)来表示;采用SPSS 18.0软件进行LSD检验和单因子、双因子方差分析。
2 结果
2.1 土壤性质
方差分析表明,麦季的pH、土壤全氮、有机质和微生物氮均显著高于稻季,土壤含水量和土壤呼吸则显著低于稻季(P<0.01)(表1,表2);作物和施肥的交互作用对pH有显著影响,对其他土壤性质影响不显著(P<0.05)(表2)。配施有机肥对稻季含水量和pH没有显著影响,却显著降低麦季土壤pH(P<0.05)(表1)。与CK相比,配施有机肥SF和PSF显著提高麦季和稻季的土壤全氮、有机质和微生物碳(P<0.05);而F、PMF与CK之间没有显著差异(P<0.05)(表1)。
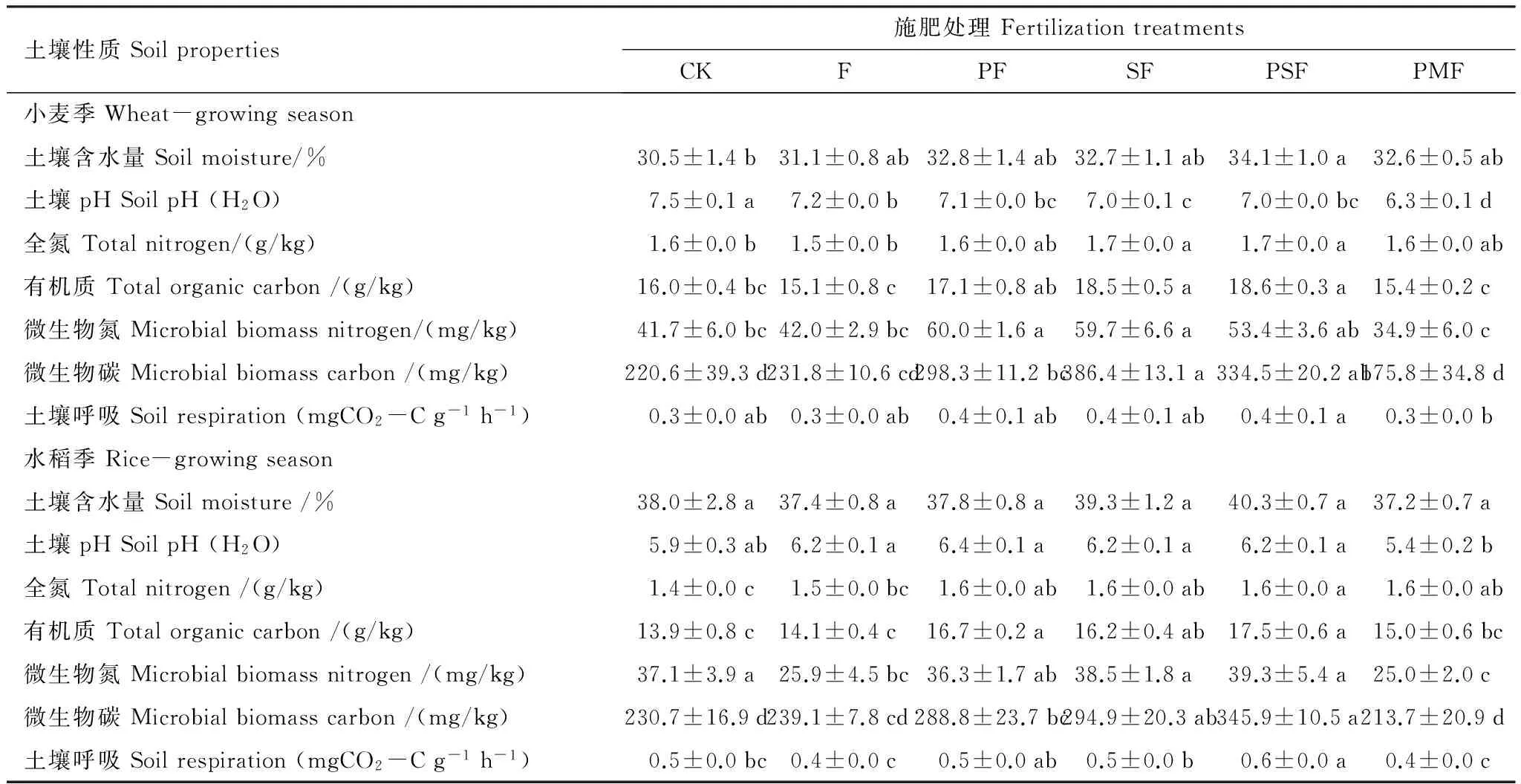
表1 不同施肥处理下小麦和水稻根际土壤性质
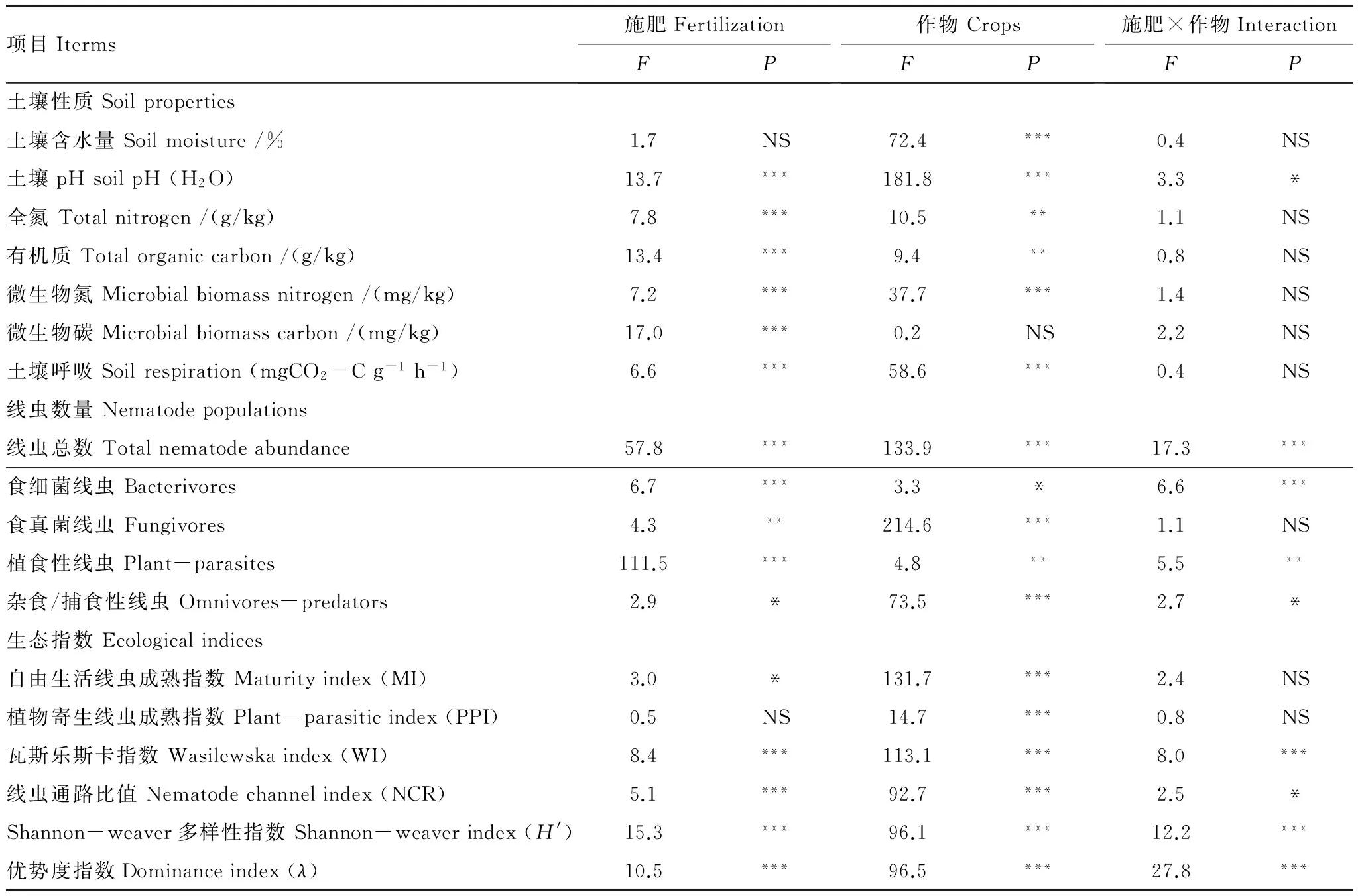
表2 土壤性质、线虫数量及生态指数方差分析
2.2 土壤线虫总数
方差分析表明,麦季线虫总数显著高于稻季(图1,表2);作物和施肥的交互作用对线虫总数的影响显著(P<0.001)(表2)。小麦根际每100g干土线虫数量范围在1307—4228条;水稻根际每100g干土线虫数量范围在1065—2004条。配施有机肥对麦季线虫总数的增加较稻季显著,两季均以PSF对线虫总数贡献最大,其次为SF,二者均和CK达到显著差异(P<0.01);F、PF和CK之间均没有显著差异(P<0.05);PMF能显著提高小麦根际线虫总数,但对水稻根际没有显著性影响(P<0.05)(图1)。
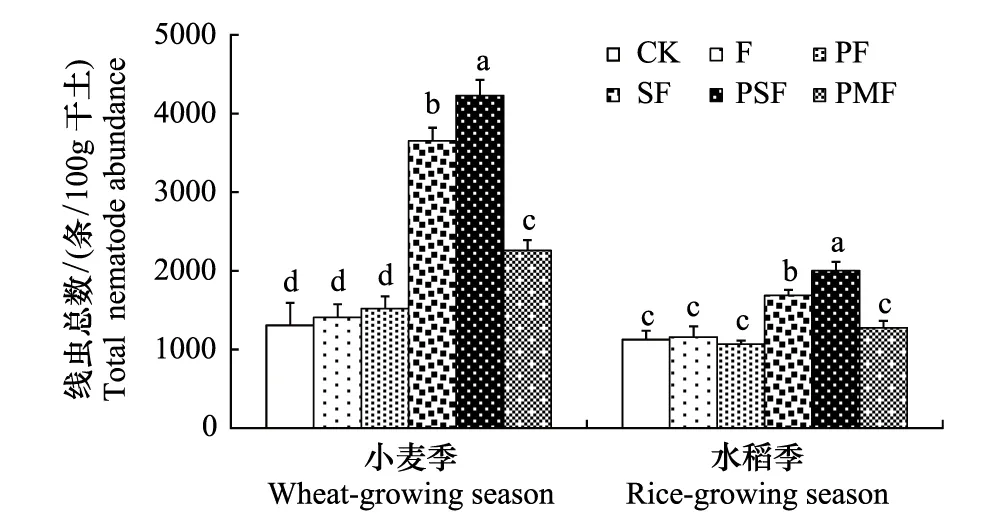
图1 不同施肥处理对小麦和水稻根际土壤线虫总数的影响 Fig.1 Effects of different fertilization treatments on total nematode abundance in the rhizosphere of wheat and rice 图中数值为平均值+标准误;不同英文字母表示同一采样时期不同处理间差异显著(P<0.05); CK:不施肥;F:100%化肥;PF:猪粪堆肥 + 50%化肥;SF:秸秆全量还田 + 100%化肥;PSF:猪粪堆肥 + 秸秆全量还田 + 50%化肥;PMF:猪粪商品有机无机复合肥. CK: no fertilizer, F: 100% chemical fertilizer, PF: pig manure compost + 50% chemical fertilizer, SF: straw return + 100% chemical fertilizer, PSF: pig manure + straw return + 50% chemical fertilizer, PMF: commercial fertilizer of pig manure mixed with chemical fertilizer
2.3 土壤线虫群落
本研究共鉴定出小麦根际土壤线虫32属,其中,食细菌线虫13属,食真菌线虫6属,植食性线虫4属,杂食/捕食性线虫9属;水稻根际土壤线虫31属,其中,食细菌线虫15属,食真菌线虫4属,植食性线虫2属,杂食/捕食性线虫10属(表3)。水稻根际各处理的线虫类群比小麦根际丰富(麦季16—24属;稻季20—26属)(表3)。两季均以PSF的线虫属最多(麦季24属;稻季26属),CK最少(麦季16属;稻季20属)(表3)。
小麦根际主要线虫优势属(个体数占土壤线虫群落个体总数10%以上)为头叶属(Cephalobus)、拟丽突属(Acrobeloides)和丝尾垫刃属(Filenchus);水稻根际主要线虫优势属为杆咽属(Rhabdolaimus)、潜根属(Hirschmanniella)和丝尾垫刃属(Filenchus)(表3)。其中,丝尾垫刃属(Filenchus)虽为两季共同的线虫优势属,但在小麦根际的丰度(平均28.8%)高于水稻根际(平均13.2%)。杆咽属(Rhabdolaimus)在水稻根际不施肥处理中的丰度(57.0%)远高于单施化肥(10.3%)和配施有机肥(平均9.9%)。潜根属(Hirschmanniella)在水稻根际的所有施肥处理(尤其是单施化肥和猪粪商品有机无机复合肥)(平均15.4%)中均高于不施肥处理(8.1%)。
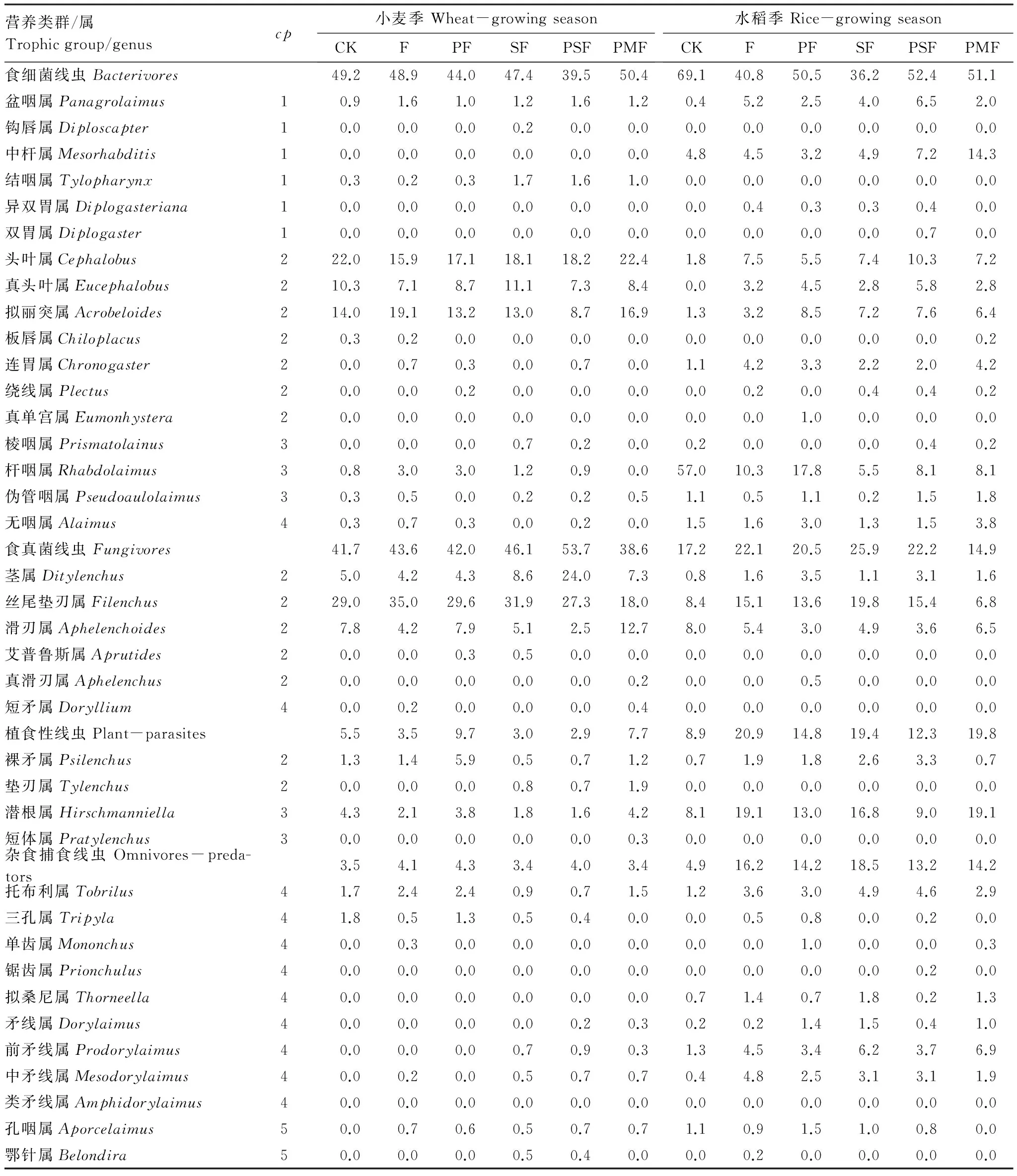
表3 小麦和水稻根际土壤线虫群落组成及其相对丰度
方差分析表明,土壤线虫各营养类群的相对丰度在施肥处理之间和两种作物之间均存在显著差异(P<0.05);作物和施肥的交互作用对食细菌线虫、植食性线虫和杂食/捕食性线虫有显著影响,而对食真菌线虫影响不显著(表2)(P<0.05)。总体来看,麦季与稻季的线虫营养类群结构存在较大差异。小麦根际所有处理的食真菌线虫丰度(麦季平均44.3%,稻季平均22.4%)高于水稻根际,植食性线虫(麦季平均5.4%,稻季平均16.0%)和杂食/捕食性线虫丰度(麦季平均3.8%,稻季平均13.5%)低于水稻根际(表3)。施肥对小麦季的线虫营养类群结构影响较小,对水稻季的影响较大(图2)。麦季中,PSF能显著提高食真菌线虫的丰度,PF显著提高植食性线虫丰度(P<0.05);食细菌线虫和杂食/捕食性线虫丰度各处理间没有显著性差异(P<0.05)(图2)。稻季中,施肥处理均显著降低食细菌线虫丰度,显著提高杂食/捕食性线虫丰度(P<0.05);除PSF外,均显著提高植食性线虫丰度(P<0.05)(图2)。
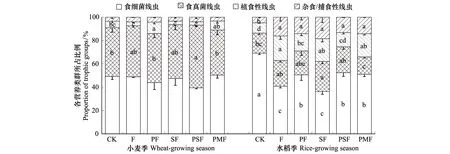
图2 不同处理对小麦和水稻根际土壤线虫营养类群比例的影响 Fig.2 Effects of different fertilization treatments on the relative abundance of nematode trophic groups in the rhizosphere of wheat and rice图中数值为平均值-标准误;不同英文字母表示同一采样时期相同营养类群比例在不同处理间差异显著(P<0.05),未标注字母即各处理间差异不显著(P>0.05)
2.4 土壤线虫生态指数
方差分析表明,水稻根际的自由生活线虫成熟指数(MI)、植食性线虫成熟指数(PPI)、线虫通路比值(NCR)和香农多样性指数(H′)均显著高于小麦根际,瓦斯乐斯卡指数(WI)和优势度指数(λ)显著低于小麦根际(P<0.001)(表2,表4);施肥与作物的交互作用对WI、NCR、H′和λ影响显著,对PPI和MI的影响不显著(P<0.05)(表2)。麦季的MI、PPI、NCR和H′在各处理间没有显著差异,SF和PSF的WI和λ值显著高于CK(P<0.05);稻季各施肥处理均显著降低WI和λ,显著提高H′,对PPI影响不显著(P<0.05)(表4)。
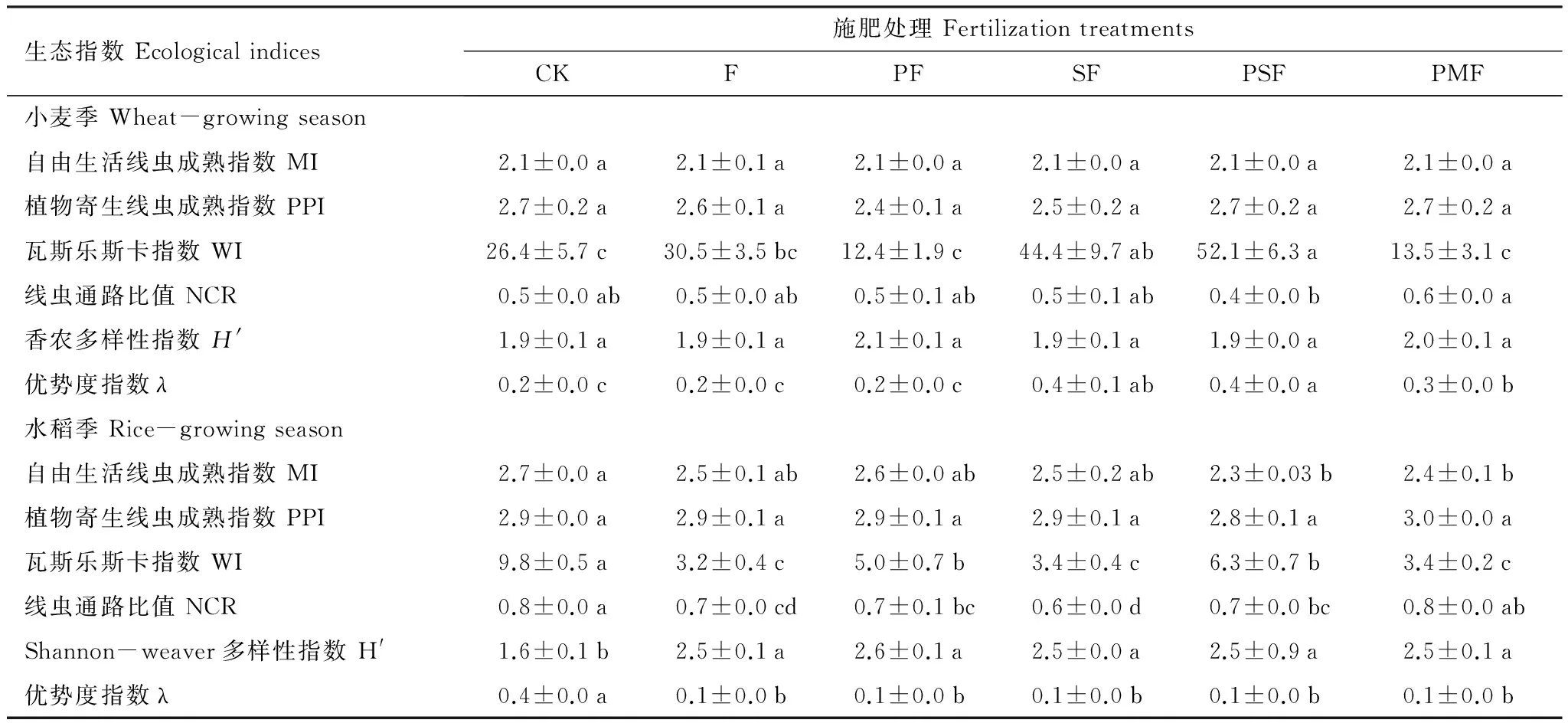
表4 小麦季和水稻季的土壤线虫生态指数
3 讨论
线虫作为土壤健康的指示生物,已被广泛应用于评价各种生态系统受干扰的情况[23]。作为农田生态系统的两项基本管理措施,施肥和轮作的人为干扰对本研究的土壤性质和线虫区系产生了影响,其交互作用对土壤性质影响较小,对线虫群落结构影响显著。有机肥的施用被证明有助于改善土壤结构,提高土壤生物多样性和活性,从而提升耕地质量[5]。在本实验中,配施秸秆的有机肥较单施化肥和不施肥均显著提高了土壤肥力、微生物活性和土壤线虫数量。然而麦季和稻季的土壤线虫群落结构有着显著差异,有机肥对两季土壤线虫的影响也存在较大差异。
3.1 麦季和稻季的线虫群落结构
整体而言,麦季线虫总数显著高于稻季,可能是因为麦季的水分含量较稻季少,通气性得到改善,有利于有机质的分解和矿化[16],使得其土壤肥力较水稻季高;同时,有机质的分解使得微生物的食物来源得到增加,进而为线虫的繁殖提供了更多的食物来源。Okada等[26]的研究也表明,水田种植的水稻土壤含氧量较少,对线虫的生长不利,而旱地种植的水稻由于土壤透气性较好,有益于线虫的生长。再者,采样方法的不同也可能导致这一结果的发生,由于对含水量较高的水稻根际土壤还没有较好的采样方法,在本实验中,将土钻贴根采集的土壤近似认为水稻根际土,而这种方法不如麦季抖根法采集的根际土壤接近根表,由于根系分泌物的存在,根表周围有大量的微生物聚集[17],土壤线虫数量自然也更多。
稻季植食性和杂食/捕食性线虫的数量显著高于麦季。Bert等[27]和Okada等[26]研究发现属于植物寄生线虫的潜根属(Hirschmanniella)线虫偏好取食水稻和莲花等淹水条件下的植物根系,并认为潜根属(Hirschmanniella)是稻田土壤中的特征属。在本实验中,稻季植食性线虫潜根属(Hirschmanniella)的丰度和植物寄生性线虫成熟指数(PPI)显著高于麦季,说明稻麦轮作比水稻连作更有助于降低土壤中有害的植食性线虫。与本文研究结果相似,Liu等[28]的研究也发现水稻土壤中矛线科Dorylaimidae的数量很丰富,他还证实长期种植水稻会使杂食/捕食性线虫丰度提高。在对很多稻田的线虫鉴定中均发现杂食/捕食性线虫数量很丰富,这可能和水稻的根系特征、稻田淹水等土壤环境有关,再者,杂食/捕食性线虫也有很多取食藻类,如托布利属(Tobrilus),这和水田中数量丰富的藻类一致[27](在采样过程中肉眼可见地表覆盖了一层绿色的藻类)。
3.2 有机肥对土壤线虫群落结构的影响
关于有机肥能够增加土壤线虫总数的结论已经得到许多研究的证明[1,9,11-12]。本实验中,对线虫总数的提高表现为秸秆+猪粪>秸秆>猪粪>单施化肥。线虫总数与有机肥的施用量、有机肥的种类及肥料的养分含量有关,从有机肥的施用量来看,秸秆配施猪粪的有机肥施用量大于秸秆或猪粪;从有机肥种类和养分含量上来看,秸秆的有机质含量(78.6%—82.6%)远高于猪粪堆肥(16.1%—45.4%)和化肥。Elfstrand[29]等的研究结果也表明土壤微生物对有机肥的响应取决于有机肥的养分含量和质量。再者,从本文土壤肥力和微生物性质的数据也可以得出线虫总数与土壤有机质和微生物碳氮含量成正相关。
许多研究表明,有机肥能够增加土壤食细菌线虫[1,9,13]、食真菌线虫[13-14]和杂食/捕食性线虫的数量[1,13,15],降低植食性线虫的数量[1,13,15]。本实验中,配施有机肥均显著降低稻季食细菌线虫丰度和瓦斯乐斯卡指数(WI);秸秆配施有机肥对麦季食细菌线虫丰度没有显著影响,却增加了食真菌线虫丰度和WI。这是因为施肥明显抑制了稻季的属于食细菌线虫的杆咽属(Rhabdolaimus)线虫的繁殖,使得其数量大量减少,杆咽属(Rhabdolaimus)拥有较高的cp值,为k-对策者,对施肥这种外界干扰较敏感[25]。与本研究结果不同的是我们对曲周小麦玉米轮作体系下线虫群落结构的调查中发现,配施有机肥均能够提高麦地土壤食细菌线虫数量和WI[30],而在本研究中,可能由于水旱轮作导致麦季的土壤和微生物环境仍受稻季的影响,使得麦季的食细菌线虫丰度并没有随着配施有机肥的施用得到增加。有机肥对麦季植食性线虫和杂食/捕食性线虫的数量影响较小,对稻季影响显著,这是因为麦季的根系特征和诸如土壤水分、pH、和肥力等土壤环境异于稻季,如上所述,水稻根系和稻田淹水的土壤环境对潜根属(Hirschmanniella)和矛线科Dorylaimidae线虫的生长有益[31],配施有机肥明显地促进了水稻根系的生长,使得潜根属(Hirschmanniella)和矛线科Dorylaimidae线虫的食物来源得到增加[32-33]。
3.3 有机肥对线虫多样性的影响
本研究中,多样性指数(H′)和优势度指数(λ)比成熟指数(MI,PPI)能更有效地指示生态系统功能的变化。施肥对H′的影响因作物而异,Hu等[1]对我国玉米小麦轮作系统下玉米季土壤线虫群落的研究发现,有机肥的施用对线虫多样性没有显著影响;Bulluck等[34]对番茄地土壤线虫的研究表明,有机肥和化肥显著降低线虫多样性。本实验中的单施化肥和配施有机肥对麦季线虫多样性没有显著影响,但却显著提高稻季线虫多样性。这是由于施肥使得稻季的杆咽属(Rhabdolaimus)线虫数量急剧减少,λ显著降低,从而H′升高,这种变化可能对线虫群落稳定性的提高,甚至其在生态系统功能中的作用有积极的影响。
综上所述,配施有机肥对土壤肥力的提高在施肥的第3年(2010—2012年)已经较为明显;配施有机肥对麦季线虫群落的影响较小,对稻季线虫群落的改变较大。从线虫总数来看,两季均表现为配施秸秆的有机肥显著高于其他施肥处理。从线虫群落来看,配施秸秆的有机肥对麦季食微线虫丰度的提高比其他施肥处理好;所有施肥处理对稻季土壤线虫的多样性均显著增加。总的来看,在本实验稻麦轮作系统下,同时添加秸秆和猪粪对提高土壤线虫数量和食微线虫丰度更有益。
[1] Hu C, Qi Y C. Effect of compost and chemical fertilizer on soil nematode community in a Chinese maize field. European Journal of Soil Biology, 2010, 46(3/4): 230-236.
[2] Yardirn E N, Edwards C A. The effects of chemical pest, disease and weed management practices on the trophic structure of nematode populations in tomato agroecosystems. Applied Soil Ecology, 1998, 7(2): 137-147.
[3] Yeates G W, Bongers T. Nematode diversity in agroecosystems. Agriculture, Ecosystems & Environment, 1999, 74(1/3): 113-135.
[4] Neher D A. Role of nematodes in soil health and their use as indicators. Journal of Nematology, 2001, 33(4): 161-168.
[5] Mäder P, Fliessbach A, Dubois D, Gunst L, Fried P, Niqqli N. Soil fertility and biodiversity in organic farming. Science, 2002, 296 (5573): 1694-1697.
[6] 傅声雷. 土壤生物多样性的研究概况与发展趋势. 生物多样性, 2007, 15(2): 109-115.
[7] Yeates G W, Bongers T, De Goede R G M, Freckman D W, Georgieva S S. Feeding habits in soil nematode families and genera—an outline for soil ecologists. Journal of Nematology, 1993, 25(3): 315-331.
[8] 李琪, 梁文举, 姜勇. 农田土壤线虫多样性研究现状及展望. 生物多样性, 2007, 15(2): 134-141.
[9] Liang W J, Lou Y L, Li Q, Zhong S, Zhang X K, Wang J K. Nematode faunal response to long-term application of nitrogen fertilizer and organic manure in Northeast China. Soil Biology and Biochemistry, 2009, 41(5): 883-890.
[10] 胡诚, 曹志平, 齐迎春, 胡菊, 李双来. 土壤线虫群落对施用EM生物有机肥的响应. 生态学报, 2010, 30(18): 5012-5021.
[11] 刘艳军, 张喜林, 高中超, 周宝库. 长期施肥对哈尔滨黑土土壤线虫群落的影响. 土壤通报, 2011, 42(5): 1112-1115.
[12] 江春, 黄菁华, 李修强, 李辉信, 孙波. 长期施用有机肥对红壤旱地土壤线虫群落的影响. 土壤学报, 2010, 48(6): 1235-1241.
[13] Nahar M S, Grewal P S, Miller S A, Stinner D, Stinner B R, Kleinhenz M D, Wszelaki A, Doohan D. Differential effects of raw and composted manure on nematode community, and its indicative value for soil microbial, physical and chemical properties. Applied Soil Ecology, 2006, 34(2/3): 140-151.
[14] Villenave C, Saj S, Pablo A L, Sall S, Djigal D, Chotte J L, Bonzi M. Influence of long-term organic and mineral fertilization on soil nematofauna when growing Sorghum bicolor in Burkina Faso. Biology and Fertility of Soils, 2010, 46(7): 659-670.
[15] Akhtar M, Mahmood I. Control of plant-parasitic nematodes with organic and inorganic amendments in agricultural soil. Applied Soil Ecology, 1996, 4(3): 243-247.
[16] 范明生, 江荣风, 张福锁, 吕世华, 刘学军. 水旱轮作系统作物养分管理策略. 应用生态学报, 2008, 19(2): 424-432.
[17] Walker T S, Bais H P, Grotewold E, Vivanco J M. Root exudation and rhizosphere biology. Plant Physiology, 2003, 132(1): 44-51.
[18] Wang X P, Zabowski D. Nutrient composition of Douglas-fir rhizosphere and bulk soil solutions. Plant and Soil, 1998, 200(1): 13-20.
[19] 毛小芳, 李辉信, 陈小云, 胡锋. 土壤线虫三种分离方法效率比较. 生态学杂志, 2004, 23(3): 149-151.
[20] 尹文英, 胡圣豪, 沈韫芬. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998: 51-89.
[21] Bongers T. The Nematodes of Netherlands. Utrecht: Foundation Publisher of Royal Dutch Natural History Society, 1994.
[22] Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 1990, 83(1): 14-19.
[23] 李玉娟, 吴纪华, 陈慧丽, 陈家宽. 线虫作为土壤健康指示生物的方法及应用. 应用生态学报, 2005, 16(8): 1541-1546.
[24] Yeates G W. Nematodes as soil indicators: functional and biodiversity aspects. Biology and Fertility of Soils, 2003, 37(4): 199-210.
[25] Yeates G W. Variation in soil nematode diversity under pasture with soil and year. Soil Biology and Biochemistry, 1984, 16(2): 95-102.
[26] Okada H, Niwa S, Takemoto S, Komatsuzaki M, Hiroki M. How different or similar are nematode communites bewteen a paddy and an upland rice fields across a flooding-drainage cycle. Soil Biology and Biochemistry, 2011, 43(10): 2142-2151.
[27] Bert W, Borgonie G. Order Rhabditida: suborder Tylenchia // Eyualem A, Andrássy I, Traunspurger W. Freshwater Nematodes: Ecology and Taxonomy. Oxfordshire: CABI Publishing, 2006: 648-695.
[28] Liu M Q, Chen X Y, Qin J T, Wang D, Griffiths B, Hu F. A sequential extraction procedure reveals that water management affects soil nematode communities in paddy fields. Applied Soil Ecology, 2008, 40(2): 250-259.
[29] Elfstrand S, Hedlund K, Mårtensson A. Soil enzyme activities, microbial community composition and function after 47 years of continuous green manuring. Applied Soil Ecology, 2007, 35(3): 610-621.
[30] 叶成龙, 刘婷, 张运龙, 张俊伶, 沈其荣, 李辉信. 麦地土壤线虫群落结构对有机肥和秸秆还田的响应.土壤学报, 2013, 50(5): 998-1006.
[31] 刘婷, 叶成龙, 陈小云, 冉炜, 沈其荣, 胡锋, 李辉信. 不同有机肥源及其与化肥配施对稻田土壤线虫群落结构的影响. 应用生态学报, 2013, 24(12): 3508-3516.
[32] Wang K H, McSorley R, Marshall A, Gallaher R N. Influence of organicCrotalariajunceahay and ammonium nitrate fertilizers on soil nematode communities. Applied Soil Ecology, 2006, 31(3): 186-198.
[33] Ferris H, Bongers T, De Goede R G M. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Applied Soil Ecology, 2001, 18(1): 13-29.
[34] Bulluck III L R, Barker K R, Ristaino J B. Influences of organic and synthetic soil fertility amendments on nematode trophic groups and community dynamics under tomatoes. Applied Soil Ecology, 2002, 21(3): 233-250.
Effects of different organic fertilizers on soil nematodes in the rhizosphere of rice and wheat cropland
LIU Ting1,2, YE Chenglong1, LI Yong3, CHEN Xiaoyun1, RAN Wei1, SHEN Qirong1, HU Feng1, LI Huixin1,2,*
1CollegeofResourcesandEnvironmentalSciences,NanjingAgriculturalUniversity,Nanjing210095,China2JiangsuCollaborativeInnovationCenterforSolidOrganicWasteResourceUtilization,Nanjing210095,China3SoilandFertilizerTechnicalguidanceStationofJintanCity,Jintan213200,China
With rapid global nutrient enrichment, the responses of soil biota to nutrient addition have gained much attention. There are numerous studies addressing the responses of soil nematodes and microorganisms to chemical or organic fertilization. Moreover, existing studies of nematode communities mostly focused on uplands as well as on bulk soil. As yet, the responses of soil nematodes in the rhizosphere to both chemical and organic fertilization remain unclear. This is of special significance for the widespread rice-wheat rotation agro-ecosystem in China, where chemical fertilization is commonly used and organic fertilization is promoted, e.g. straw return. The aim of our study was to assess the effects of both chemical and organic fertilization on soil nematodes in the rhizosphere of rice-wheat rotation agro-ecosystems in eastern China. We applied fertilization treatments (CK: no fertilizer, F: 100% chemical fertilizer, PF: pig manure compost + 50% chemical fertilizer, SF: straw return + 100% chemical fertilizer, PSF: pig manure + straw return + 50% chemical fertilizer, PMF: commercial fertilizer of pig manure mixed with chemical fertilizer) in a factorial experimental design to rice-and wheat-cropland in Jintan, Jiangsu Province. We measured soil nematode abundance and diversity, as well as soil nutrients and soil environmental parameters in the rice and wheat rhizospheres. Two-way analysis of variance (ANOVA) was performed after testing variable normality and homogeneity of variance. Duncan tests were used for post-hoc comparisons.Cephalobus,AcrobeloidesandFilenchuswere the dominant genera in the wheat-growing season; whileRhabdolaimus,HirschmanniellaandFilenchusdominated in the rice-growing season. Total nematode abundance was significantly higher in the wheat-growing season than rice-growing season; SF and PSF increased the total abundance of nematodes compared to PF and PMF; there was no significant difference between CK and F. For nematode trophic groups, the relative abundance of fungivores was significantly higher in the wheat-growing season than the rice-growing season, while plant-parasites and omnivores/predators tended to be more abundant in the rice-growing season. In terms of individual abundance and diversity, the effects of organic fertilization on the soil nematode community were more pronounced in the rice-growing season than in the wheat-growing season. For example, the straw addition tended to increase the Wasilewska index (WI), while both 100% chemical fertilizer and all the combined organic fertilizer treatments increased the Shannon-weaver index (H′) but deceased the Dominance index (λ). This suggests that the responses of soil nematodes to fertilization depend not only on crop type, but also on fertilizer quality and nutrient content. For example, combined addition of straw and pig manure was most advantageous for improving both nematode abundance and the relative abundance of microbivores in our study system.
rhizosphere; organic fertilizer combined with chemical fertilizer; rice-wheat rotation agro-ecosystem
农业部行业公益性项目-利用有机(类)肥料调控我国土壤微生物区系关键技术研究(201103004)
2014-02-16; < class="emphasis_bold">网络出版日期:
日期:2014-12-04
10.5846/stxb201402160264
*通讯作者Corresponding author.E-mail: huixinli@njau.edu.cn
刘婷, 叶成龙, 李勇, 陈小云, 冉炜, 沈其荣, 胡锋, 李辉信.不同有机类肥料对小麦和水稻根际土壤线虫的影响.生态学报,2015,35(19):6259-6268.
Liu T, Ye C L, Li Y, Chen X Y, Ran W, Shen Q R, Hu F, Li H X.Effects of different organic fertilizers on soil nematodes in the rhizosphere of rice and wheat cropland.Acta Ecologica Sinica,2015,35(19):6259-6268.
猜你喜欢
今日农业(2022年14期)2022-09-15
中国土壤与肥料(2021年5期)2021-12-02
现代畜牧科技(2021年3期)2021-07-21
今日农业(2020年19期)2020-12-14
猪业科学(2018年4期)2018-05-19
中成药(2018年2期)2018-05-09
天然产物研究与开发(2018年3期)2018-05-07
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
中国蔬菜(2016年8期)2017-01-15
天然产物研究与开发(2016年6期)2016-06-05
