
中国西南喀斯特植物群落演替特征及驱动机制
2015-01-18 08:51宋同清王克林彭晚霞曾馥平曾昭霞何铁光
生态学报 2015年17期
文 丽, 宋同清,*, 杜 虎, 王克林, 彭晚霞, 曾馥平, 曾昭霞, 何铁光
1 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 2 中国科学院环江喀斯特生态系统观测研究站, 环江 547100 3 广西农科院资源与环境研究所, 南宁 530000
中国西南喀斯特植物群落演替特征及驱动机制
文 丽1,2, 宋同清1,2,*, 杜 虎1,2, 王克林1,2, 彭晚霞1,2, 曾馥平1,2, 曾昭霞1,2, 何铁光3
1 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 2 中国科学院环江喀斯特生态系统观测研究站, 环江 547100 3 广西农科院资源与环境研究所, 南宁 530000
植被恢复是西南喀斯特石漠化治理和生态重建的关键步骤。通过总结已有的结果和分析中国科学院环江喀斯特试验站部分试验数据,研究了西南喀斯特植被演替过程中群落的动态特征,结果发现:西南喀斯特沿北、中、南亚热带至北热带的海拔递减、气温和降雨量升高的梯度格局下,群落的高度、盖度增加,密度、科属种数量和物种多样性呈递减趋势;不同区域沿草丛、灌丛、灌乔丛、乔丛的顺向演替方向上,群落的高度、盖度增加、密度减小,科属种数量及物种多样性除北亚热带持续增加外,其他3个区域的最大值均出现在灌乔丛,乔丛反而有所下降;西南喀斯特植被演替是生物驱动因素与环境阻力相互作用的结果,不同区域不同演替阶段植被演替的建群种和关键种不同,其植物群落的稳定性受多样性和结构性的共同控制,因地质和生态环境恶劣,植被退化容易恢复难,充分理解和认识植被演替规律对西南喀斯特植被恢复和生态重建意义重大。
植被演替; 群落特征; 驱动机制; 气候带; 西南喀斯特
植物群落是指在特定空间和时间范围内,具有一定的植物种类组成和一定的外貌及结构与环境形成一定相互关系并具有特定功能的植物集合体。群落演替是一个植物群落被另一个植物群落所取代的过程,它是植物群落动态的一个最重要的特征[1]。植物群落最早由林奈(Carolus Linnaeus)于18世纪中叶提出[2],经过近两百年特别是近百余年的积淀,产生了不同的植物群落学派,其中以Clements为代表,提出了整体论观点和顶极群落概念[3-4],而Gleason提出了群落的个体论观点,强调偶然性和随机性对群落形成和维持的重要性[5-6]。Clements与Gleason之争实际上是群落构建中的确定性与随机性之争,也就是当今群落生态学中的两个最重要的理论即生态位理论和中性理论的“较量”。两种理论经过长期的竞争和演绎,进入20世纪90年代后进化并发展了种库假说、中性理论、生态学代谢理论、中域效应假说等新观点、新假说和新理论,生态位理论、能量假说、历史成因假说和稍早提出的Rapoport法则也在这一时期得到了大量数据的检验。
植被是一个地区植物的总体,是生态环境的综合反映,植被演替和分布的格局是多个生态过程的产物,这些生态过程主要包括物种的进化(物种的形成、迁移及灭亡)、地理差异及环境因子(地质、地貌、气候、土壤)等的控制[7-8],同时不同程度上受到植被演化历史和干扰的影响[9]。在气候、地形、地貌、土壤、干扰等自然、人为作用和过程控制下,植被个体之间长期的相互作用导致了不同尺度、不同层次上的时空变异, 形成了有规律的植被时空格局,其分布既反映植被对环境资源的利用与适应关系,也反映植被特性及更新、竞争等种内种间关系[10]。植被在时间和空间上的分布是典型的分维数体[11],其尺度依赖性使得植被的分布格局表现为空间上的不确定性和复杂性[12]。
世界范围内中国西南喀斯特是世界上湿润一半湿润、热带一亚热带中高山、断陷盆地、高原、峡谷、峰丛洼地、槽谷、峰林平原、溶丘洼地喀斯特地貌系列发育最好的地区[13]。各地貌类型景观异质性强,小生境复杂,其特殊的地质、气候和干扰历史,演化出了石面、土面、石土面、石沟、石缝、石坑、石洞等多样小生境类型[14],发育了各式各样的生态系统和生物多样性。西南喀斯特具有从热带雨林到亚热带湿润、半湿润常绿阔叶林,常绿阔叶+落叶阔叶混交林,山地针叶、阔叶混交林,针叶林、灌丛、草甸等多样的植被类型,据不完全统计,中国西南喀斯特有高等植物225 科,1213 属,4287 种[15],分别占中国西南高等植物科的90%,属的60%及种的30%,其中包括140 种国家一级和二级保护植物。中国西南喀斯特顶极群落为亚热带喀斯特常绿落叶阔叶混交林,由于受地球内动力、强烈的地质运动、高温多雨且分布不均、碳酸盐岩溶蚀性强、水文二维结构明显以及其适生植物具有嗜钙性、耐旱性和石生性等限制特点的综合影响,生境和植被具有高度异质性,生态系统的稳定性和抗干扰性差,与黄土、沙漠、寒漠并列为中国四大生态环境脆弱区,尖锐的人地矛盾导致了植物的正向、反向演替,出现了稀疏草丛、草丛、草灌丛、藤刺灌丛、灌丛、乔灌丛、乔丛等各种不同演替阶段或生态群落的共存局面[16],有许多地方甚至出现了生态环境退化十分严重的喀斯特石漠化现象,是我国植被恢复和生态重建的重点和难点地区[17-18],充分理解和认识西南喀斯特植被演替过程中群落的动态特征对岩溶植被退化生态系统的恢复和重建意义重大。
1 研究区概况与数据采集
西南喀斯特植被演替是一个极其漫长的过程,需要上百年甚至更长的时间[19],退化容易恢复难,不同气候带、不同地貌类型、不同小生境、不同人为干扰等条件下植被演替的方向、格局和时间等不同[20]。本文以喀斯特中亚热带的广西环江西南部喀斯特集中连片区为研究对象,在草丛、灌丛、灌乔丛、乔丛4个典型演替阶段各选择2个代表性群落类型,草丛:水竹叶(Mllrdanniatriquetra)、白茅(Imperatacylindrica);灌丛:黄荆(Vitexnegundo)、盐肤木(Rhuschinensis);灌乔丛:八角枫(Alangiumchinense)、栀子皮(Itoaorientalis);乔丛:侧柏(Platycladusorientalis)、铁榄(Sinosideroxylonpedunculatum)。在坡向、坡度、海拔等基本相同或相似的坡中下位分别建立3个20 m × 20 m的样方,共计24个样方。用插值法将样方细分为4个10 m × 10 m的小型样方和16个5 m × 5 m的微型样方,以5 m× 5 m的微型样方为基本单元,调查每个胸径(DBH)≥1 cm的个体,记录树种名称、胸径、树高、冠幅和生长状况等;按梅花型随机抽取5个微型样方,调查灌木和草本的种类、数量、高度和生长状况等。同时,用高精度全球定位系统(global positioning system,GPS) (E640+MobileMapper)记录样方内中心的经纬度、海拔等地理信息,调查记录其坡向、坡位、坡度、岩石裸露率、土层厚度等情况。基于气候带的分布和已有的研究状况,同时采集刘玉国等[21]、司彬等[22]、温远光等[23]、区智等[24]在西南喀斯特北亚热带的云南富源和贵州普定、南亚热带的广西马山、北热带的广西弄岗的研究成果,研究西南喀斯特区域尺度范围内不同演替阶段植被组成、物种多样性及结构的变化规律,从而为西南喀斯特植被的保护、恢复和石漠化治理提供理论依据和参考。不同研究区域基本概况见表1。
2 科属种组成
北亚热带、中亚热带、南亚热带和北热带的植物群落科属种的数量并非随着演替的不断深入而增大(表2),而是从草丛阶段开始,科属种不断增加至灌乔丛阶段达到最大值,到达乔丛阶段时物种种数量却有一定下降。这是因为从草本植物侵人裸岩荒地形成草丛开始一直到灌乔丛阶段,随着生物对环境的作用,恶劣的生态环境不断得到改善,使得环境的容纳量不断提高,故物种数量也不断地提高。而达到乔丛之后,由于群落的稳定性增大,群落建群种和各层优势种逐渐稳定,各物种分别占据分化了的生态位,群落内各种群的竞争趋于稳定,物种浸入较困难,一些物种由于在对环境资源的利用竞争中处于劣势,而无法继续生存,逐渐退出群落,因而科属种的数量又会有所降低,但这种降低,不是群落的衰退,相反是表明群落的成熟和稳定。一般认为,随着气候、土壤等环境条件的改善物种的多样性越来越高,结构性越来越好。西南喀斯特由北亚热带、中亚热带、南亚热至北热带海拔梯度下降,年均温、年降雨量增加的梯度,群落的高度和覆盖度越来越大、密度越来越小、群落的结构越来越好,但由于海拔高度的降低,小生境相对均匀,因此物种多样性呈递减趋势(表2)。

表1 各研究区基本概况
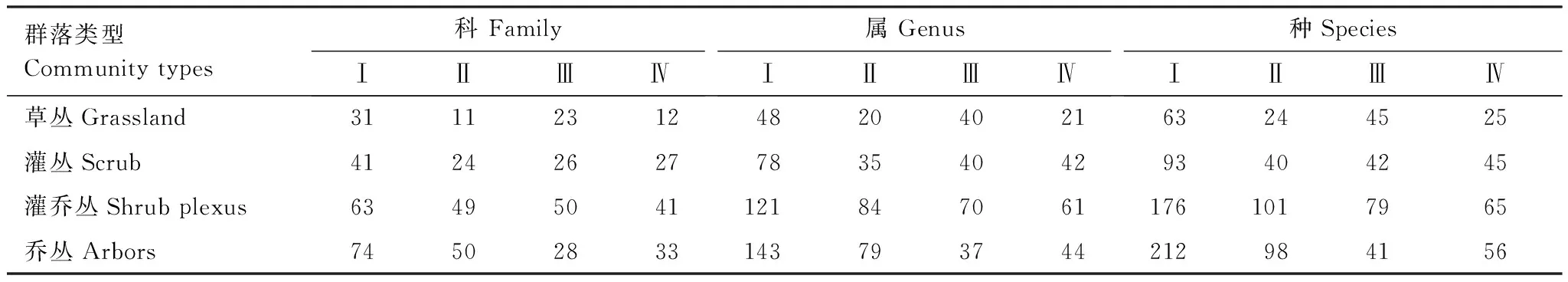
表2 不同区域不同演替阶段群落的科、属、种
3 物种组成
3.1 乔丛
3.1.1 北亚热带
北亚热带湿润季风气候区贵州普定县不存在这原始喀斯特森林植被,现存恢复最好的天然森林为乔丛乔木林群落。该群落主要是川钓樟以圆果化香树(Platycaryalongipes)、窄叶石栎(Lithocarpusconfinis)、滇鼠刺(Iteayunnanensis)、安顺润楠(Machiluscavaleriei)和槲栎(Quercusaliena)为优势种的常绿落叶阔叶混交林。常见的乔木树种还有短萼海桐(Pittosporumbrevicalyx)、黑弹树(Celtisbungeana)、朴树(Celtissinensis)、刺楸(Kalopanaxseptemlobus)、多脉猫乳(Rhamnellamartinii)和云贵鹅耳枥(Carpinuspubescens)等。灌木层以异叶鼠李(Rhamnusheterophylla)、倒卵叶旌节花(Stachyurusobovatus)、刺异叶花椒(Zanthoxylumovalifoliumvar. spinifolium)、薄叶鼠李(Rhamnusleptophylla)、小叶菝葜(Smilaxmicrophylla)和小果蔷薇(Rosacymosa)为优势种, 常见的其他灌木树种还有贵州花椒(Zanthoxylumbungeanum)、瑞香(Daphneodora)、珊瑚冬青(Llexcorallina)、香叶树(Linderacommunis)、铁仔(Myrsineafricana)、黑果菝葜(Smilaxglaucochina)和杭子梢(Campylotropismacrocarpa)等。草本层主要有大披针薹草(Carexlanceolata)、千里光(Senecioscandens)、求米草(Oplismenusundulatifolius)、阔叶山麦冬(Liriopeplatyphylla)、十字薹草(Carexcruciata)、野雉尾金粉蕨(Onychiumjaponicum)、三脉紫菀(Asterageratoides)、对马耳蕨(Polystichumtsus-simense)、三枝九叶草(Epimediumsagittatum)、茜草(Rubiacordifolia)、分枝大油芒(Spodiopogonramosus)、大叶唐松草(Thalictrumfaberi)和画笔南星(Arisaemapenicillatum)等。藤本植物主要有香花崖豆藤(Millettiadielsiana)、青蛇藤(Periplocacalophylla)、异叶爬山虎(Parthenocissusheterophylla)、藤黄檀(Dalbergiahancei)、紫花络石(Trachelospermumaxillare)、柱果铁线莲(Clematisuncinata)、地果(Ficustikoua)和常春藤(Hederanepalensisvar.sinensis)等。林下岩石上主要生长着薜荔(Ficuspumila)、波缘冷水花(Pileacavaleriei)和吊石苣苔(Lysionotuspauciflorus)等攀援或匍匐植物以及石韦(Pyrrosialingua)、瓦韦(Lepisorusthunbergianus)、抱石莲(Lepidogrammitisdrymoglossoides)、西南石韦(Pyrrosiagralla)、线裂铁角蕨(Aspleniumcoenobiale)和肿足蕨(Hypodematiumcrenatum)等蕨类植物。
3.1.2 中亚热带
中亚热带季风气候区广西环江成熟林乔木层主要包括青冈(Cyclobalanopsisglauca)、伊桐(Itoaorientalis)、南酸枣(Choerospondiasaxillaris)、伞花木(Eurycorymbuscavaleriei)、圆果化香树(Platycaryalongipes)、小梾木(Swidapaucinervis)、单性木兰(Kmeriaseptentrionalis)、翠柏(Calocedrusmacrolepis)、青檀(Pteroceltistatarinowii)、铁榄(Sinosideroxylonpedunculatum)、东女贞(Ligustrumjaponicum)、小叶栾树(Boniodendronminus)、掌叶木(Handeliodendronbodinieri)、菜豆树(Radermacherasinica)、广西密花树(Rapaneakwangsiensis)、檵木(Loropetalumchinense)等;灌木层主要包括齿叶黄皮(Clausenadunniana)、九里香(Murrayaexotica)、长叶水麻(Debregeasialongifolia)、白毛长叶紫珠(Callicarpalongifoliavar.floccosa)、石山苎麻(Boehmeriastrigosifolia)、南天竹(Nandinadomestica)、紫凌木(Decaspermumfruticosum)、金樱子(Rosalaevigata)、羊蹄甲(Bauhiniapurpurea)、石岩枫(Mallotusrepandus)、火麻(Cirardiniasuborbiculata)、红背山麻秆(Alchorneatrewioides)、山乌桕(Sapiumdiscolor)、杜茎山(Maesajaponica)、通脱木(Tetrapanaxpapyrifer)等;草本层包括大苞冷水花(Pileabasicordata)、单座苣苔(Metabriggsiaovalifolia)、淡竹叶(Lophatherumgracile)、石韦、蜘蛛抱蛋(Aspidistraelatior)、火炭母(Polygonumchinense)、五节芒(Miscanthusfloridulus)、肾蕨(Nephrolepiscordifolia)、卷柏(Selaginellatamariscina)、千里光(Senecioscandens)、珍珠茅(Scleriaherbecarpa)等。
3.1.3 南亚热带
南亚热季风气候区广西马山县成熟林中乔木层有19种,以青冈栎、南酸枣、海红豆(Adenantherapavonina)、润楠(Machilusnanmu)和仪花(Lysidicerhodostegia)为主。灌木层的种类共有24种,主要以乔木幼树为主,主要有青冈栎、西南山茶(Camelliapitardiivar. yunnanica)、假鹰爪(Desmoschinensis)、海红豆、九节(Psychotriarubra)、野桐(Mallotustenuifolius)和润楠为主。草本层的优势种分别为井栏边草(Pterismultifida)、海南海金沙(Lygodiumcircinnatum)、灰绿耳蕨(Polystichumanomalum)、薯蓣(Dioscoreapolystachya)、薄叶卷柏(Selaginelladelicatula)、崖姜(Pseudodrynariacoronans)和海南海金沙、灰绿耳蕨、团叶鳞始蕨(Lindsaeaorbiculata)、井栏边草、金粟兰(Chloranthusspicatus)。
3.1.4 北热带
北热带季风气候区广西弄岗由于阳性树种的生长,使得林下形成荫蔽环境,年幼时耐荫的中性树种增加,例如:肥牛树(Cephalomappasinensis)、蚬木(Excentrodendronhsienmu)、闭花木(Cleistanthussumatranus)、金丝李(Garciniapaucinervis)等树种的幼树找到了合适的生存环境而侵人。随着时间的推移,这些中性树种的幼树幼苗生长成为大乔木,占据林内的上层,而阳性树种因在林内不能得到更新而逐渐消失,退出竞争舞台,林下长人耐荫的灌木和草本植物。至此,稳定的森林群落就形成了。
3.2 灌乔丛
3.2.1 北亚热带
贵州普定县的灌乔丛多为极度退化群落逐渐向乔丛演替的一个阶段,主要有圆果化香树、安顺润楠、滇鼠刺、麻栎(Quercusacutissima)、槲栎、朴树、刺楸、多脉猫乳、川钓樟(Linderapulcherrimavar. hemsleyana)、窄叶石栎和李(Prunussalicina)等;灌木树种主要有小果蔷薇、异叶鼠李、薄叶鼠李、倒卵叶旌节花、针齿铁仔(Myrsinesemiserrata)、黑果菝葜、雀梅藤(Sageretiathea)、杭子梢、川榛(Corylusheterophyllavar. sutchuenensis)、刺异叶花椒、香叶树和瘤枝密花树(Myrsineverruculosa)等;草本层植物主要有大披针薹草、千里光、阔叶山麦冬、五节芒、对马耳蕨、野雉尾金粉蕨、降龙草(Hemiboeasubcapitata)、分枝大油芒、求米草、十字薹草、三脉紫菀和舌叶薹草(Carexligulata)等;岩石上生长着大量的石韦和抱石莲,藤本植物主要有香花崖豆藤、藤黄檀、青蛇藤、紫花络石、异叶爬山虎和柱果铁线莲等。
3.2.2 中亚热带
广西环江县灌乔丛多为有灌丛向灌乔丛过渡的阶段,乔木层主要树种包括野桐、八角枫(Alangiumchinense)、小叶女贞(Ligustrumquihoui)、盐肤木(Rhuschinensis)、灰毛浆果楝(Cipadessacinerascens)、菜豆树、石山榕(Ficusvirens)等小乔木为主;灌木层代表性种类有黄荆、红背山麻秆、长叶柞木(Xylosmalongifolium)、杜茎山、冻绿(Rhamnusutilis)、东女贞、羊蹄甲、水冬瓜(Saurauiaoldhamii)、鸡矢藤(Paederiascandens)、石山苎麻、紫麻(Oreocnidefrutescens)等;草本层主要包括素馨香(Jasminumofficinalevar. grandiflorum)、纤毛鸭舌草(Murdanniatriquetra)、蕨(Pteridiumaquilinumvar. latiusculum)、竹叶草(Oplismenuscompositus)、沿阶草(Ophiopogonbodinieri)、楼梯草(Elatostemainvolucratum)、千里光、木贼(Equisetumhyemale)等。
3.2.3 南亚热带
广西马山县乔丛共有26种,以青冈栎、海红豆、苦木(Picrasmaquassioides)、乌材(Diospyroseriantha)、厚壳桂(Cryptocaryachinensis)、乌桕(Sapiumsebiferum)、虎皮楠(Daphniphyllumoldhami)和仪花为主;灌木层的种类最丰富,共51种,主要有青冈栎、假鹰爪、九节、网脉酸藤子(Embeliarudis)、龙须藤(Bauhiniachampionii)、乌材、杜茎山、齿叶黄皮、山茶(Camelliajaponica)和海红豆;草本层的优势种与成熟林相似。
3.2.4 北热带
广西弄岗灌乔丛由于灌木的定居与生长,使得光照、温度等环境条件进一步得到改善,为一些阳性树种的生存提供条件。任豆(Zeniainsignis)、东京桐(Deutzianthustonkinensis)、海南椴(Diplodiscustrichosperma)、翻白叶树(Pterospermumheterophyllum)等主要树种得以发展。继而,阳性的乔木树种生长,逐渐形成岩溶森林。
3.3 灌丛
3.3.1 北亚热带
贵州普定县的灌丛多为极度退化后的藤刺灌丛向灌乔丛演替的阶段,以小果蔷薇、火棘(Pyracanthafortuneana)、异叶鼠李、野拔子(Elsholtziarugulosa)和竹叶花椒(Zanthoxylumarmatum)为优势种。常见的灌木树种还有六月雪(Serissajaponica)、多叶勾儿茶(Berchemiapolyphylla)、马棘(Indigoferapseudotinctoria)、珍珠荚蒾(Viburnumfoetidumvar. ceanothoides)、铁仔、香薷(Elsholtziaciliata)、中华绣线菊(Spiraeachinensis)、黄脉莓(Rubusxanthoneurus)、金丝桃(Hypericummonogynum)、马桑(Coriarianepalensis)、红叶木姜子(Litsearubescens)、来江藤(Brandisiahancei)、软条七蔷薇(Rosahenryi)、薄叶鼠李、杭子梢、匍匐栒子(Cotoneasteradpressus)、黑果菝葜和悬钩子蔷薇(Rosarubus)等。乔木树种较少,零星可见安顺润楠、滇鼠刺、构树(Broussonetiapapyrifera)、香椿(Toonasinensis)、盐肤木、刺楸和圆果化香树等。草本层物种丰富,主要有黄背草(Themedajaponica)、大披针薹草、对马耳蕨、矛叶荩草(Arthraxonlanceolatus)、三脉紫菀、硬秆子草(Capillipediumassimile)、千里光、臭根子草(Bothriochloabladhii)、大丁草(Gerberaanandria)、牡蒿(Artemisiajaponica)、堇菜(Violaverecunda)、五节芒、爵床(Justiciaprocumbens)、假俭草(Eremochloaophiuroides)和蒲公英(Taraxacummongolicum)等。层间植物种类较少,主要有地果(Ficustikoua)和毛葡萄(Vitisheyneana)等。
3.3.2 中亚热带
广西环江县灌丛主要有麻疯树(Jatrophacurcas)、白竹(Fargesiasemicoriacea)、桂楠(Phoebekwangsiensis)、黄荆、红背山麻杆、羊蹄甲、香叶树、粗糠柴(Mallotusphilippinensis)、鸭脚木、算盘子(Glochidionpuberum)、粗叶悬钩子(Rubusalceifolius)、密榴木(Miliusachunii)、紫麻、灰岩棒柄花(Cleidionbracteosum)、石山苎麻、圆叶乌桕(Sapiumrotundifolium)、假苹婆(Sterculialanceolata)、火棘、厚叶楤木(Araliadasyphylloides)、老虎刺(Pterolobiumpunctatum)、小芸木(Micromelumintegerrimum)等;草本层主要有五节芒、珍珠茅、海金沙(Lygodiumjaponicum)、翠云草(Selaginellauncinata)、千里光、野葛(Puerariamontana)、肾蕨、蔓生莠竹(Microstegiumfasciculatum)、类芦(Neyraudiareynaudiana)、蜈蚣草(Eremochloaciliaris)、纤毛鸭舌草等;少量乔木树种包括粉苹婆(Sterculiaeuosma)、虾公木(Brideliaretusa)、菜豆树、小果白桐(Mallotusmicrocarpus)、麻楝(Chukrasiatabularis)、通脱木等。
3.3.3 南亚热带
广西马山县灌丛阶段的灌木层物种也较丰富,共 21种,以黄荆、雀梅藤、红背山麻杆、地桃花(Urenalobata)和花椒(Zanthoxylumbungeanum)为主。灌丛阶段草本优势种有荩草(Arthraxonhispidus)、蔓生莠竹、小苦荬(Lxeridiumdentatum)、兰香草(Caryopterisincana)和弓果黍(Cyrtococcumpatens)。
3.3.4 北热带
广西弄岗的灌丛主要以草丛向灌丛过渡的阶段为主,在草丛群落发展到一定时期,群落环境条件由于草本植物与恶劣的环境发生了能量与物质的交换而得到一定的改善,一些喜光的阳性灌木藤本植物出现,例如酒饼叶(Moslacavaleriei)、山石榴(Catunaregamspinosa)、鸡爪簕(Oxycerossinensis)、斜叶澄广花(Oropheaanceps)、剑叶龙血树(Dracaenacochinchinensis)等与一些高草混生形成灌草丛(也可认为是草丛阶段向灌丛阶段演替的中间过渡阶段),以后灌木、藤本植物大量增加,占据优势成为灌丛群落。这个阶段最明显的特征是刺生藤本大量出现。
3.4 草丛
3.4.1 北亚热带
贵州普定县的草丛群落是植被恢复破坏后恢复的初始阶段,以黄背草、大披针薹草、黄茅(Heteropogoncontortus)和阔叶山麦冬为优势种。其他常见草本植物主要有抱石莲、臭根子草、石韦、细柄草(Capillipediumparviflorum)、千里光、东亚唐松草(Thalictrumminusvar. hypoleucum)、细锥香茶菜(Rabdosiacoetsa)、火绒草(Leontopodiumleontopodioides)、矛叶荩草和野雉尾金粉蕨等。灌木主要是杭子梢、金丝桃、马棘和野拔子等矮小灌木或半灌木。
3.4.2 中亚热带
广西环江县草丛主要有斑茅(Saccharumarundinaceum)、白茅(Imperatacylindrica)、蔓生莠竹、五节芒、指叶艾(Artemisiasubdigitata)、毛蕨(Cyclosorusinterruptus)、翠云草等;草丛内散生的灌木种类有冻绿、构树、盐肤木、火棘、马桑、白饭树(Flueggeavirosa)等。
3.4.3 南亚热带
广西马山县草丛优势种为臭根子草、类芦、弓果黍、蜈蚣草和荩草;草丛阶段灌木种类较丰富,有26种,优势种为黄荆、老鼠耳(Berchemialineata)、黄牛木(Cratoxylumcochinchinense)、余甘子(Phyllanthusemblica)、岗松(Baeckeafrutescens)、青篱柴(Tirpitziasinensis)和黄花稔(Sidaacuta)。
3.3.4 北热带
广西弄岗草丛撂荒地、裸岩荒山在没有人为干扰的情况下,首先是一年生草本定居,如:白花丹(Plumbagozeylanica)、胜红蓟(Ageratumconyzoides)等,然后一些喜阳、耐旱、抗逆性强的草本植物,例如:飞机草(Eupatoriumodoratum)、蔓生莠竹、白茅等,能够适应裸岩石山的恶劣环境相继侵人定居,并形成优势群落。
4 物种多样性
随着立地条件、气候等环境的改善和植被的顺向演替发展,群落的物种多样性增加,喀斯特地区不同区域、不同演替阶段、不同层次的物种多样性不同(表3)。
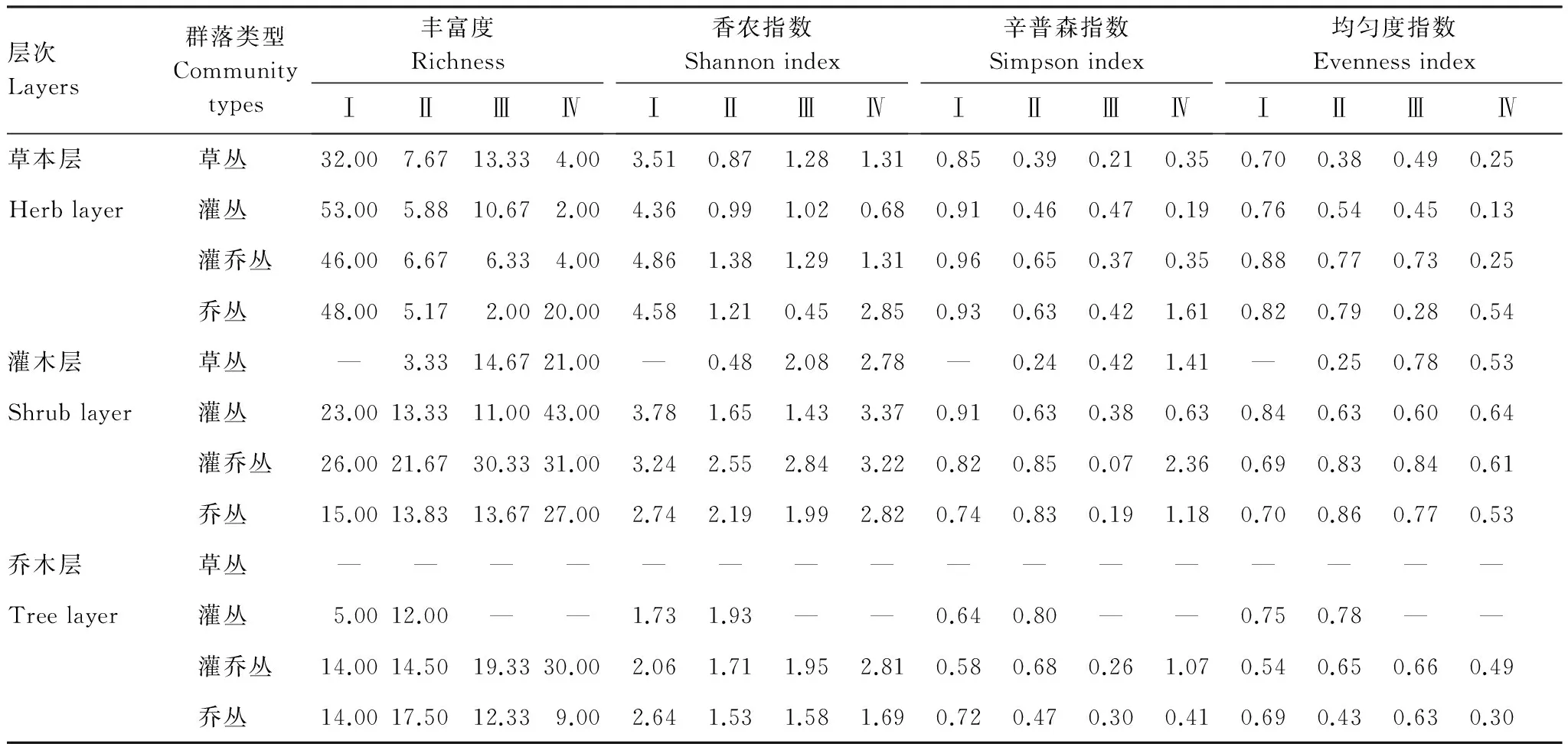
表3 不同区域不同演替阶段物种多样性
草本层的丰富度总体上是北亚热带>南亚热带>中亚热带>北热带,其中北亚热带灌丛最高,中亚热带差别不大,南亚热带草丛最高,北热带乔丛最高;草本的多样性指数总体上是北亚热带>北热带>南亚热带>中亚热带,除北热带乔丛最高外,其他地区均以灌乔丛的多样性指数最高;植物群落的优势度变化趋势总体上与多样性相反。
灌木层的物种丰富的总体趋势是北热带>北亚热带>南亚热带>中亚热带,除北热带在灌丛阶段最高外,其他演替阶段均以灌乔丛的物种丰富度最高;灌木层的多样性指数北热带和北亚热带差别不大,且均在灌丛阶段最高,明显高于南亚热带和中亚热带,南亚热带和中亚热带以灌乔丛最大;灌木层植物群落生态优势度总体趋势上以北热带最高,其他变化规律不强。
乔木层的物种丰富度随着北亚热带、南亚热带、中亚热带到北亚热带温度水分的提高而逐渐增大,其中北、中亚热带灌乔丛和乔丛的差别不大,但在南亚热带和北热带灌乔丛的物种丰富度明显增高;乔木层的物种多样性总的趋势是北热带和北亚热带大于中、南亚热带,其中除北亚热带灌乔丛的物种多样性最高外,其他3个区域均以灌乔丛物种丰富度最大,群落优势度除南亚热带较低之外,其他区域及不同演替阶段变化规律不强。
5 群落结构
植物群落的高度受环境的影响较大,西南喀斯特植物群落随北亚热带、中亚热带、南亚热带由北向南气温升高、降雨量增大梯度,草丛和灌丛的差异不大,但灌乔丛和乔丛植物的高度逐渐增加,其中乔丛的高度分别为7.85、8.74、11.51 m;各区域随草丛、灌乔丛、乔丛顺向演替,植物群落的高度明显增加(表4)。不同区域不同演替阶段植物群落的密度不同,喀斯特草丛的密度以中亚热带和北亚热带最高,但灌丛、灌乔丛和乔丛的密度南亚热带明显增多;不同区域随植被的顺向演替密度均呈逐渐减小的趋势。
植物群落的盖度受气候的影响较小,不同区域盖度的变化不大,但盖度是高度和密度共同作用的结果,不同演替阶段植物群落的盖度不同,在草本和灌丛阶段个体虽小,但密度较大,植物盖度较高;灌乔丛个体居中,但密度较小,盖度下降,林内光照较好,有利于植被迅速生长;乔丛的密度虽小,但个体较大,其盖度最高(表4)。

表4 不同区域不同演替阶段群落结构
6 植物群落演替的驱动机制
6.1 群落演替的过程与方向
中国西南喀斯特景观异质性强,岩石裸露率高、土层瘠薄且不连续、生态条件恶劣而导致其造林技术、造林条件、造林成活率、生长抚育等各方面受诸多因子控制,植被恢复主要依靠自然演替,与常态地貌上植被演替的一般规律一样分为6个阶段:草本群落阶段、灌草群落阶段、灌木群落阶段、灌木灌丛阶段、灌乔过渡阶段、乔丛阶段。其乔丛亚热带喀斯特常绿落叶阔叶混交林是基于光、热、水、土及地貌单元的组合差异与植物生理生态生长习性长期竞争、适应演化而成的以不同常绿、落叶阔叶混交林为背景的各种植被的镶嵌格局,优势种不明显,多为共有种群落,大多数植物具有喜钙性、石生性、耐旱性、耐瘠性[25]。由于喀斯特地区生境复杂,能适应同一演替阶段生境物种不单是一个,而是具有相同适应性的多个物种构成的种组,主要包括先锋种、次先锋种、过渡种、次顶极种、顶极种5个种组[26]。
中国西南喀斯特石面、土面、石土面、石沟、石缝、石坑、石洞等各种各样的小生境导致产生了不同的基质和斑块[14],能为不同演替阶段提供不同的物种种源,在演替初期阶段如草丛甚至还有顶极物种,其定居格局没有遵循经典的演替原则,演替一开始就具有多阶段性,加上岩溶区经常性的人为干扰使任何物种能被其他物种所代替,演替的方向并不单一,如草本群落可同时分别向灌丛、灌木林、乔木林等阶段发展。目前多采用的“空间代替时间”的研究方法很难证明不同演替阶段群落起源、发生、发展、经历的同质性,研究结果的普遍性和真实性应更依赖于长期定位观测。由于喀斯特生境的脆弱性,当干扰的强度、频度、持续时间超过一定的阈值时群落极易发生逆向演替,植被产生双向的自然演替[27],又加上喀斯特地区二元结构明显、水土资源少且流失快,逆向演替速度远比顺序演替的快,退化容易恢复难。
6.2 群落演替的主要驱动力
植被演替驱动力的理论主要有种群竞争更替(生物)假说和环境变化适应(非生物)假说[28-29]。一般认为,区域尺度上,气候、母质、植物区系决定了植被类型,在景观及更小的尺度上,非地带性环境因子主导着植被的演替和分布格局,其中地形、土壤理化性质和微生物环境是影响景观尺度植被演替和分布格局最主要的因子之一,他们之间的协同作用形成了不同的植被演替和分布格局。西南喀斯特植被恢复与气候、母质、土壤等区域环境因子关系密切,但主要驱动力是高度的景观异质性、地形、土壤理化性状和微生物环境、物种的生理生态生长特性及其相互竞争关系[20,30]。
6.2.1 非生物因素
中国西南喀斯特地区水热条件优越,但受地球内动力、强烈的地质运动、高温多雨且分布不均、碳酸盐岩溶蚀性强、水文二维结构明显与景观异质性高的影响,具有岩石裸露率和钙镁含量高,土壤瘠薄,季节性岩溶干旱严重等自然胁迫环境,其适生植物具有石生性、嗜钙性、耐瘠性和耐旱性等限制性特点[31]。
中国西南喀斯特景观异质性高,众多裸露的岩石附近形成了许多如窝窝土、凹面地形等极有利于植物进入、定居和生长的“安全岛”[32]和“避风区”[33],这些微区域风速较低,水流速度较慢,积水效应明显,突出的岩石为水汽的凝结创造了条件,形成了相对优越的各种微区域环境,有利于植物的着落、萌发和生长。
土壤基质的理化特征在演替的早期阶段对侵入的物种有某种“过滤作用”[34]。中国西南喀斯特土壤是由碳酸盐岩发育而成的石灰土,钙镁含量与pH值高,植物具有嗜钙性和耐碱性,茶科植物、马尾松等地带性酸性植物在石灰土上根本无法定居和生存。土壤缺乏是植被难以生长的最主要因素,植被演替的关键是以土壤为核心的土壤-水-植被紧密相关复合生态系统的养分积累,植物演替伴随着土壤累积。但喀斯特地区碳酸盐岩发育的石灰土缺乏C层,土壤与岩石之间直接刚性接触,其亲和力和粘着力差,极易产生水土流水和块体滑移[35],二元结构明显导致地表径流虽小、但漏失现象严重,且碳酸盐岩溶蚀性强,成土速度缓慢,其成土时间是其他母岩10倍以上[36],因此植物的自然定居过程极其缓慢,从草丛、灌丛等低级阶段演替进化到乔丛群落少则需要几十年、多则上百年。
6.2.2 生物因素
植物的入侵、定居和种间竞争对群落演替至关重要,早期阶段,生物之间的互助关系比竞争关系对演替的进程影响更大[37- 39]。西南喀斯特众多裸露的岩石表面存在丰富的苔藓类植物,改善了土壤的水分、养分状况,提高了土壤的有机质含量,降低了pH值,有利于维管束植物的定居、生存和发展[40]。同时一些高等植物在演替的早期能够定居成功,得益于他们和一些具有特殊功能的微生物存在共生关系,如固碳细菌、菌根真菌。彭晚霞[41]等基于喀斯特人工林、次生林、原生林3个不同的森林植被恢复阶段动态监测样地(200 m×40 m)调查发现,其豆科植物的重要值为39.383、5.607、4.875,具有明显的下降趋势,这种高等植物与微生物之间的互利关系在植物定居中的作用是目前该领域研究的热点之一。
西南喀斯特不同演替阶段的环境不同,种间竞争关系也不同,草丛阶段岩石裸露率高,土壤和植被的覆盖度低,光、温、水、热变化剧烈,生境条件恶劣,喜光、耐寒、繁殖力强的草种成为驱动群落演替的关键种;随着草丛群落覆盖度的提高和环境的改善,具有显著的有性和无性繁殖能力及克隆繁殖能力特别强的灌木是灌丛阶段的关键种,不同区域草、灌木关键种不同,其中北亚热带草本为黄背草、大披针薹草、黄茅、阔叶山麦冬,灌木为小果蔷薇、火棘、异叶鼠李、野拔子、竹叶花椒;中亚热带草本为斑茅、白茅、蔓生莠竹、指叶艾、干旱毛蕨(Cyclosorusaridus)、鬼针草(Bidenspilosa),灌木为黄荆、红背山麻秆、白竹、聚果羊蹄甲、麻疯树、桂楠;南亚热带草本为臭根子草、类芦、弓果黍、蜈蚣草、荩草,灌木为黄荆、雀梅藤、红背山麻秆、地桃花、花椒;北热带草本为白花丹、胜红蓟、飞机草、蔓生莠竹、白茅,灌木为酒饼叶、山石榴、鸡爪簕、斜叶澄广花、剑叶龙血树。灌丛的恢复和生长为乔木树种的定居创造了条件,在人为干扰较少,土壤种子库资源丰富(若干扰严重、资源缺乏,灌木将长期保持其优势地位而阻止群落的正向演替)情况下,西南喀斯特保留较丰富的顶极群落的优势种和关键种促进灌丛向灌乔丛和乔丛发展。其中北亚热带的关键种为圆果化香、窄叶石栎、滇鼠刺、安顺润楠、槲栎、朴树、刺楸中;亚热带:青冈、伊桐、南酸枣、圆果化香、单性木兰、铁榄、东女贞、小叶栾树、菜豆树、广西密花树、檵木;南亚热带:青冈栎、南酸枣、海红豆、润楠、仪花、厚壳桂、乌桕;北热带:肥牛树、蚬木、闭花木、金丝李、任豆树、东京桐、海南锻、翻白叶树。
6.3 群落演替的物种多样性与稳定性
一般认为随着气候、土壤等环境条件的改善物种的多样性越来越高,结构性越来越好。西南喀斯特由北亚热带、中亚热带、南亚热至北热带年均温、年降雨量增加的梯度,群落的高度和覆盖度越来越大、密度越来越小、群落的结构越来越好,从区域角度出发,物种的多样性越来越高,但由于受小环境和人为干扰的影响,从群落水平来看物种多样性呈递减趋势。
生态学理论认为生态系统的稳定性与植物多样性密切相关[42- 44],在目前尚没有合适的指标对稳定性进行衡量时,多数学者仍主要以群落的复杂性(多样性)作为衡量稳定性的指标[45],随着群落演替阶段发展,群落的稳定性越来越好,物种多样性越来越高,顶极群落的稳定性一般要高于演替中的群落,植物物种多样性可以导致群落稳定性[16,26,46- 51]。西南喀斯特不同区域随着草丛、灌丛、灌乔丛、乔丛正向演替发展,植物群落高度和覆盖度增大,密度减小,结构越来越好;西南喀斯特亚热带季风湿润气候区的贵州普定没有发现原生性很强的乔丛,只存在发育较成熟的灌乔丛,其植物群落的科属种数量及物种多样性随演替的深入而直线上升,但中亚热带、南亚热带和北热带不同层次的植物群落科属种数量和物种多样性并不完全随群落的顺向演替而升高,其最大值出现在灌乔丛,至乔丛时反而有所下降。研究结果说明西南喀斯特植物群落的稳定性与多样性具有更为复杂的关系,植物多样性是群落稳定的基础或必要条件,但并不能完全代表群落的稳定性,群落的稳定性既要有丰富的植物多样性,同时也要保证群落结构的合理性[52-53]。
[1] 骆林川, 熊文愈. 琅琊山森林群落演替预测. 南京林业大学学报, 1989, 13(1): 1- 9.
[2] Makoto N. Methods for Ecological Study. Tokyo: Kokin Shou-in, 1979.
[3] Clements F E. Nature and structure of the climax. Journal of Ecology, 1936, 24(1): 252- 284.
[4] Clements F E. Plant Succession: An Analysis of the Development of Vegetation. Washington: Carnegie Institution of Washington, 1916.
[5] Gleason H A. The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club, 1926, 53(1): 7- 26.
[6] Gleason H A. The structure and development of the plant association. Bulletin of the Torrey Botanical Club, 1917, 44(10): 463- 481.
[7] Whittaker R J, Willis K J, Field R. Scale and species richness: towards a general, hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4): 453- 470.
[8] Willis K J, Whittaker RJ. Species diversity scale matters. Science, 2002, 295(5558): 1245- 1248.
[9] Qian H, Ricklefs R E. A comparison of the taxonomic richness of vascular plants in China and the United States. The American Naturalist, 1999, 154(2): 160- 181.
[10] Legendre P, Fortin M J. Spatial pattern and ecological analysis. Vegetation, 1989, 80(2): 107- 138.
[11] 柳锦宝, 姚云军, 张永福. 基于分维的多尺度植被空间格局分析. 水土保持研究, 2007, 14(1): 155- 158
[12] 杜华强, 汤孟平, 周国模, 徐文兵, 刘恩斌, 施拥军. 天目山物种多样性尺度依赖及其与空间格局关系的多重分形. 生态学报, 2007, 27(12): 5038- 5049.
[13] 朱华. 中国南方石灰岩(喀斯特)生态系统及生物多样性特征. 热带林业, 2007, 35(S1): 44- 47.
[14] 王世杰, 卢红梅, 周运超,谢丽萍, 肖德安. 茂兰喀斯特原始森林土壤有机碳的空间变异性与代表性土样采集方法. 土壤学报, 2007, 44(3):475- 483.
[15] 文和群,许兆然,J. Villa-Lobos, L. E. Skon. 中国南部石灰岩濒危植物的初步研究. 广西植物, 1993, 13(1): 41- 47.
[16] Wen Y G, Sun D J, Zhu H G, Liu J T, Liu S R, Shi Z M. Changes in aboveground biomass and diversity between different stages of secondary succession of a karst vegetation in Guangxi, China // Hu J, Advances in Biomedical Engineering. USA: Information Engineering Research Inst, Newark, 2011: 420- 423.
[17] 袁道先. 现代岩溶学在我国的发展. 中国科学基金, 2005, 19(3): 139- 141.
[18] 贺庆棠, 陆佩玲. 中国岩溶山地石漠化问题与对策研究. 北京林业大学学报, 2006, 28(1): 117- 120.
[19] 曾馥平, 彭晚霞, 宋同清, 王克林, 吴海勇, 宋希娟, 曾昭霞. 桂西北喀斯特人为干扰区植被自然恢复22年后群落特征. 生态学报, 2007, 27 (12): 5110- 5119.
[20] 李先琨, 蒋忠诚, 黄玉清, 向悟生, 吕仕洪, 叶铎, 苏宗明. 桂西南岩溶山地优势植物种群动态及其对岩溶作用的影响. 地球学报, 2008, 29(2): 253- 259.
[21] 刘玉国, 刘长成, 魏雅芬, 刘永刚,郭柯. 贵州省普定县不同植被演替阶段的物种组成与群落结构特征. 植物生态学报, 2011, 35(10): 1009- 1018.
[22] 司彬, 姚小华, 任华东, 李生, 何丙辉. 滇东喀斯特植被恢复演替过程中物种多样性研究. 西南大学学报: 自然科学版, 2009, 31(1): 132- 139.
[23] 温远光, 雷丽群, 朱宏光, 刘虹, 覃林, 马祖陆, 王克林, 庄嘉, 蓝嘉川, 龙涛, 陆晓明, 邓艳, 谢益君, 王家妍. 广西马山岩溶植被年龄序列的群落特征. 生态学报, 2013, 33(18): 5723- 5730
[24] 区智, 李先琨, 吕仕洪, 向悟生, 苏宗明, 陆树华. 桂西南岩溶植被演替过程中的植物多样性. 广西科学, 2003, 10(1): 63- 67.
[25] 彭晚霞, 王克林, 宋同清, 曾馥平, 王久荣. 喀斯特脆弱生态系统复合退化控制与重建模式. 生态学报, 2008, 28(2): 811- 820
[26] 喻理飞, 朱守谦, 叶镜中, 魏鲁明, 陈正仁. 退化喀斯特森林自然恢复过程中群落动态研究. 林业科学, 2002, 38(1): 1- 7.
[27] 马遵平, 谢泽氡. 南方岩溶区植被自然演替恢复研究综述. 四川林勘设计, 2006, (1): 1- 6.
[28] 彭少麟, 王伯荪. 鼎湖山森林群落演替之研究. 华南植物学报, 1993, (试刊2):34- 42.
[29] 王伯荪, 彭少麟. 鼎湖山森林群落分析 V.群落演替的线性系统与预测. 中山大学学报, 1985, (4): 75- 80.
[30] Zimmerman J K, Aide T M, Rosario M, Serrano M, Herrera L. Effects of land management and a recent hurricane on forest structure and composition in the Luquillo experimental forest, Puerto Rico. Forest Ecology and Management, 1995, 77(1/3): 65- 76.
[31] 袁道先. 我国西南岩溶石山的环境地质问题. 世界科技研究与发展,1997, 19(5): 41- 43..
[32] Harper J L, Clatworthy J N, Mcnaughton I H. The evolution of closely related species living in the same area. Evolution, 1961, 15: 451- 457.
[33] Jumppone A, Väre H, Matt son K G, Ohtonen R, Trappe J M. Characterization of ‘safe sites’ for pioneers in primary succession on recently deglaciated terrain. Journal of Ecology, 1999, 87(1): 98- 105.
[34] Pietsch W, Schtoz A. Vegetations entwicklung oauf Kipprohobden deroffen landschaft-Rollefr die Bioindikation. // Httl, R, Weber, E, Klem, D. Okologisches Entwicklung Spotential Von Bergbaufol Geland Schaften. Berlin: De Gruyter, 1999: 101- 118.
[35] 曹建华, 袁道先, 章程, 蒋忠诚. 受地质条件制约的中国西南岩溶生态系统. 地球与环境, 2004, 32(1): 1- 8.
[36] 蒋忠诚, 袁道先. 表层岩溶带的岩溶动力学特征及其环境和资源意义. 地球学报, 1999, 20(3): 302- 308
[37] Bliss L C, Peterson K M. Plant succession, competition and the physiological constraints of species in the Arctic. // Chapin F S III, Jeffries R L, Reynolds J F, Shaver G R, Svoboda J. Physiological Ecology Series: Arctic Ecosystems in a Changing Climate, an Ecophysiological Perspective. San Diego, CA: Academic Press, 1992: 111- 136.
[38] Walker L R, Chapin F S III. Interactions among processes controlling successional change. Oikos, 1987, 50(1): 131- 135.
[39] Chapin F S III, Walker L R, Fastie C L, Sharman L C. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska. Ecological Monographs, 1994, 64(2): 149- 175.
[40] Sohlberg E, Bliss L C. Microscale pattern of vascular plant distribution in two high arctic plant communities. Canadian Journal of Botany, 1984, 62(10): 2033- 2042.
[41] 彭晚霞. 喀斯特峰丛洼地森林植被分布格局及其维持机制研究[D]. 长沙: 湖南农业大学, 2009.
[42] Tilman D, Downing J A. Biodiversity and stability in grasslands. Nature, 1994, 367(6461): 363- 365.
[43] Baskin Y. Ecosystem function of biodiversity. Biological Science, 1995, 44: 657- 660.
[44] Lehman C L, Tilman D. Biodiversity, stability, and productivity in competitive communities. The American Naturalist, 2000, 156(5): 534- 532.
[45] 张金屯. 植被数量生态学方法. 北京: 中国科学技术出版社, 1995.
[46] 马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究 Ⅱ.丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268- 277.
[47] 刘京涛, 温远光, 周峰. 桂西南退化喀斯特植被自然恢复研究. 水土保持研究, 2009, 16(3): 65- 69.
[48] 穆彪, 杨立美, 张莉. 喀斯特植被恢复过程的群落演替特征. 西南大学学报: 自然科学版, 2008, 30(6): 91- 95.
[49] 宋同清, 彭晚霞, 曾馥平, 王克林, 欧阳资文. 桂西北喀斯特人为干扰区植被的演替规律与更新策略. 山地学报, 2008, 26(5): 597- 604.
[50] 宋同清, 彭晚霞, 曾馥平, 王克林, 覃文更, 谭卫宁, 刘璐, 杜虎, 鹿士杨. 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报, 2010, 34(3): 298- 308.
[51] 彭晚霞, 宋同清, 曾馥平, 王克林, 傅伟, 刘璐, 杜虎, 鹿士杨, 殷庆仓. 喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系. 生态学报, 2010, 30(13): 3472- 3481.
[52] 王国宏. 再论生物多样性与生态系统的稳定性. 生物多样性, 2002, 10(1): 126- 134.
[53] 杜虎, 彭晚霞, 宋同清, 王克林, 曾馥平, 鹿士杨, 时伟伟, 唐成, 谭秋锦. 桂北喀斯特峰丛洼地植物群落特征及其与土壤的耦合关系. 植物生态学报, 2013, 37(3): 197- 208.
The succession characteristics and its driving mechanism of plant community in karst region, Southwest China
WEN Li1,2, SONG Tongqing1,2,*, DU Hu1,2, WANG Kelin1,2, PENG Wanxia1,2, ZENG Fuping1,2, ZENG Zhaoxia1,2, HE Tieguang3
1KeyLaboratoryofAgro-ecologicalProcessesinSubtropicalRegion,InstituteofSubtropicalAgriculture,ChineseAcademyofSciences,Changsha410125,China2HuanjiangObservationandResearchStationofKarstEcosystem,ChineseAcademyofSciences,Huanjiang547100,China3AgriculturalResourceandEnvironmentResearchInstitute,GuangxiAcademyofAgriculturalSciences,Nanning530000,China
Vegetation restoration is the key step of desertification prevention and ecological rehabilitation in the karst region in Southwest China. A sound understanding of the vegetation composition, species diversity, and structural variation with succession is of great significance for vegetation protection and rehabilitation. However, until now, a systematic synthesis of information was not available for this region. In this study, we thoroughly collected the relevant data from the literature and unpublished data from Huanjiang Observation and Research Station of Karst Ecosystem, Chinese Academy of Sciences. We then systematically analyzed the data to determine the dynamics and driving mechanisms of plant community succession in the karst region of southwest China. To facilitate the analyses, the whole region was divided into northern subtropical, mid subtropical, southern subtropical, and northern tropical regions. The height of the plant community and vegetation coverage increase in the mean annual temperature and precipitation or with a decline in altitude in subtropical and northern tropical regions. Along with decreasing altitude, community structure improved, however, the community density, number of families, genera, species, and diversity decreased probably due to the increase in landscape homogeneity. Along the succession from grassland to shrubland, shrub/arbor mixed woodland, and to arborland, the height of the community and the coverage increased, but density decreased for the entire region. The number of families, genera, species, and diversity increased along the succession in the northern subtropical region, but these indices reached maximum in shrub/arbor mixed woodlands in the remaining three regions. The vegetation succession and distribution pattern were the result of multiple ecological processes including species evolution (speciation, migration, and extinction), geographical differentiation and environmental factors (e.g., geology, topography, climate, soil), and other processes. Under the impacts of natural factors (soil, climate, topography), human disturbances, and other ecological factors, long-term interactions among plant individuals resulted in spatial and temporal variation of vegetation distribution at different scales and levels. The main driving forces of vegetation succession included biological factors (microbial community, physiological ecology, and growth characteristics of species) and abiotic factors (e.g., landscape heterogeneity, topography, physical and chemical properties of the soil) and their interactions. The constructive species and key specie were variable for different successional stages and regions. With vegetation development, the climax species became richer, and the key species had a tendency to be more advanced, large in size, and long-living. The relationship between stability and diversity of plant communities was more complex. In general, along the community succession sequence, the stability was greater for the climax community than for the earlier succession stages. However, although plant species diversity helped to sustain community stability, a stable community was not only controlled by high diversity of plants, but also by reasonable community structure. In the karst region of the Southwest China, vegetation was found to degrade more easily than to recover, owing to the dual karst flow systems, high landscape heterogeneity, and fragile ecological conditions. In summary, the present study thoroughly synthesized the information in terms of dynamics of vegetation composition, diversity, and structure along with succession and the underlying mechanisms in the karst region of the Southwest China. Such information will be useful for the vegetation protection and rehabilitation in this region.
vegetation succession; community characteristics; driving mechanism; climatic zone; karst region in Southwest china
中国科学院西部行动计划项目(KZCX2-XB3- 10); 国家自然科学基金项目(31370485, 31370623); 国家科技支撑计划(2011BAC09B02); 中国科学院战略性先导科技专项(XDA05050205, XDA05070404)
2013- 10- 19;
日期:2014- 11- 03
10.5846/stxb201310192524
*通讯作者Corresponding author.E-mail: songtongq@isa.ac.cn
文丽, 宋同清, 杜 虎, 王克林, 彭晚霞, 曾馥平, 曾昭霞, 何铁光.中国西南喀斯特植物群落演替特征及驱动机制.生态学报,2015,35(17):5822- 5833.
Wen L, Song T Q, Du H, Wang K L, Peng W X, Zeng F P, Zeng Z X, He T G.The succession characteristics and its driving mechanism of plant community in karst region, Southwest China.Acta Ecologica Sinica,2015,35(17):5822- 5833.
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
乡村地理(2019年2期)2019-11-16
亚热带农业研究(2019年2期)2019-09-05
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
星星·诗歌原创(2017年5期)2017-08-13
中国果菜(2015年2期)2015-03-11
亚热带资源与环境学报(2015年4期)2015-01-22
文化月刊·下旬刊(2014年6期)2014-08-28
河南科技(2014年18期)2014-02-27
