
血管性认知障碍小鼠脑内少突胶质细胞再生分化障碍特点研究
2015-01-11 18:52李海龙郑惠文刘敏黎佳思王云霞赵善民毕晓莹并列第一作者
中国卒中杂志 2015年11期
李海龙,郑惠文,刘敏,黎佳思,王云霞,赵善民,毕晓莹(并列第一作者)
血管性痴呆(vascular dementia,VD)是全球导致痴呆的第二位疾病,仅次于阿尔茨海默症[1]。缺血性脑血管病变所引起的额叶、颞叶、边缘系统等部位神经元大量死亡是引起其认知障碍重要的病理基础,但目前针对其认知功能的下降尚无有效的治疗。近些年的研究发现,在成年哺乳动物的脑内存在大量的神经干/祖细胞,且这些细胞终身具有产生新生神经元的能力[1-2]。室管膜下区(subventricular zone,SVZ)及海马齿状回(dentate gyrus,DG)的颗粒下层(subgranular zone,SGZ)是神经干/祖细胞分化产生新生神经细胞的主要区域。有研究发现,脑缺血可诱发成年沙鼠持续的神经再生[3],且脱髓鞘损伤后少突胶质细胞祖细胞(oligodendrocyte progenitorcells,OPCs)数量反应性增多[4]。促进血管性痴呆中受损神经元的再生修复,从而替代死亡的神经细胞功能可能是未来VD重要的治疗靶点。本研究旨在通过建立VD小鼠模型,观察全脑缺血后室管膜下区和海马齿状回神经元再生以及少突胶质细胞再生、分化、成熟的特点,为寻找血管性认知障碍的治疗靶点提供理论依据。
1 材料方法
1.1 动物及分组 研究采用健康CD1雄性小鼠48只,8~10周,体重25~30 g,由第二军医大学实验动物中心提供。编号后采用SAS软件随机分为模型组和假手术组,每组24只,造模前常规条件下饲养1周。
1.2 血管性痴呆小鼠模型构建 参照通用的反复缺血再灌注法进行血管性痴呆小鼠造模[5]。步骤如下:0.3%戊巴比妥(75 mg/kg)腹腔麻醉,取仰卧位,门齿及四肢固定,分离双侧颈动脉,动脉夹阻断供血15 min后恢复供血10 min,再次阻断供血15 min后移除动脉夹,术中观察小鼠呼吸、心跳,术毕缝合皮肤,置于恒温板上复苏,小鼠清醒后放回笼中饲养。假手术组除不阻断颈动脉供血外,其余手术操作与模型组相同。术后14 d和28 d每组分别随机取12只小鼠处死后取脑组织进行检测。
1.3 新生细胞标记及取材 术后第4天起予腹腔注射5-溴脱氧尿嘧啶核苷(bromodeoxyuridine,BrdU,美国Sigma公司)连续3 d(150 mg/kg,1次/日)标记新生细胞。动物生理盐水灌心处死后,剥离脑组织,4%多聚甲醛固定,30%蔗糖水脱水后,制备冰冻切片,切片厚度30μm,在前囟后1.28 mm到2.12 mm间,自前向后用冰冻切片机切取脑组织冠状面切片,裱片后-20°冰箱保存。
1.4 免疫组化染色 组织切片经晾片、漂洗、穿膜后分别与一抗4°孵育过夜。一抗包括:小鼠BrdU单克隆抗体(1∶200稀释,美国Sigma公司)标记细胞增殖;兔神经胶质抗原2(neural/glial antigen2,NG2)抗体(1∶200稀释,美国Millipore公司)标记少突胶质细胞祖细胞;小鼠少突胶质细胞O4(oligodendrocyte marker O4)单克隆抗体(1∶300稀释,美国Millipore公司)标记少突胶质细胞前体细胞;兔谷胱甘肽S转移酶(glutathione-S-transferase pi,GST-pi)抗体(1∶250稀释,美国Epitomics公司)标记成熟少突胶质细胞。次日加入辣根过氧化物酶标记的羊抗兔或羊抗小鼠的相应二抗(1∶500稀释),室温孵育75 min,二氨基联苯胺显色,封片后显微镜下观察。
1.5 免疫荧光双标共聚焦显像 组织切片脱氧核糖核酸变性后分别与BrdU抗体和兔神经元特异性核蛋白(neuronal specific neucleoprotein,NeuN)抗体(1∶400,美国Abcam公司)或兔胶质纤维酸性蛋白(glial fibrillary acidic protein,GFAP)抗体(1∶2000稀释,美国Epitomics公司)的混合物4℃共同孵育过夜。再分别用相应的荧光素标记的二抗孵育后,封片在激光共聚焦显微镜下观察神经元的再生分化。
1.6 细胞计数 每只小鼠选取5张切片,通过观察每张脑切片中SVZ及DG区6个不同视野免疫组化染色为阳性的细胞数进行计数,细胞计数以每mm2的细胞数表示,计算全部视野的平均值作为其对应的细胞数。免疫荧光双标染色采用激光共聚焦显微镜成像,每张切片选5个视野,采用Image-Pro Plus 6.0图像分析软件进行双染细胞数计数,计算全部视野双染细胞数的平均值。
2 结果
2.1 血管性痴呆小鼠室管膜下区细胞增殖和再生能力增强 免疫组化染色显示,术后14 d与28 d模型组与假手术组室管膜下区BrdU阳性细胞均有表达(图1)。术后14 d模型组与假手术组BrdU阳性细胞数分别为161.1±16.9/mm2、85.7±7.3/mm2,术后28 d有所回落,分别为99.2±11.0/mm2、59.5±5.7/mm2,术后2周和4周时模型组较假手术组BrdU阳性细胞数均显著增多(t=13.64,P<0.001;t=18.79,P<0.001)。鉴于新生神经元形成需要一定时间周期,因此仅对术后28 d的脑组织进行新生神经元的观察。采用BrdU、NeuN抗体分别标记新生细胞和神经元细胞核,免疫荧光双标共聚焦显像,绿色荧光标记新生细胞,红色荧光标记神经元胞核(图2A)。BrdU/NeuN双阳性细胞进行计数后显示,模型组与假手术组术后28 d的BrdU/NeuN双阳性细胞数分别为66.3±4.6/mm2、37.9±5.9/mm2,模型组较假手术组新生神经元数量显著增多(t=13.04,P<0.001)。

图1 术后14 d与术后28 d室管膜下区BrdU标记的新生细胞增殖情况对比
2.2 血管性痴呆小鼠海马齿状回神经祖细胞向少突胶质细胞再生但进一步分化成熟障碍基于神经祖细胞向少突胶质细胞的再生分化同样具有一定的周期,故选用术后28 d的组织切片对少突胶质细胞再生过程中不同分化阶段的细胞进行观察。免疫组化显示,NG2抗体标记的少突胶质细胞祖细胞、O4抗体标记的少突胶质细胞前体细胞、GST-pi抗体标记的成熟少突胶质细胞在两组均有表达(图3A、3B、3C)。细胞计数显示,模型组NG2阳性细胞数为511.0±22.8/mm2,假手术组NG2阳性细胞数为448.8±20.9/mm2,模型组较假手术组NG2阳性细胞数量显著增多(P<0.001)。模型组O4阳性细胞数为66.8±11.4/mm2,假手术组O4阳性细胞数为80.5±10.5/mm2,模型组较假手术组O4阳性细胞数量减少(P=0.006)。模型组GST-pi阳性细胞数为133.5±11.6/mm2,假手术组GST-pi性细胞数为140.2±10.9/mm2,但两组无显著差异(P=0.161)。
2.3 血管性痴呆小鼠海马齿状回新生星形胶质细胞增多 对术后28 d的小鼠脑组织海马齿状回采用免疫荧光共聚焦成像进行新生星形胶质细胞的观察。BrdU、GFAP抗体分别标记新生细胞和星形胶质细胞,红色荧光标记新生细胞,绿色荧光标记星形胶质细胞(图2B)。细胞计数显示,模型组与假手术组术后28 d的BrdU/GFAP双阳性细胞分别为8±2.9/mm2、5.3±2.2/mm2,模型组较假手术组新生的星形胶质细胞数量显著增多(P=0.015)。
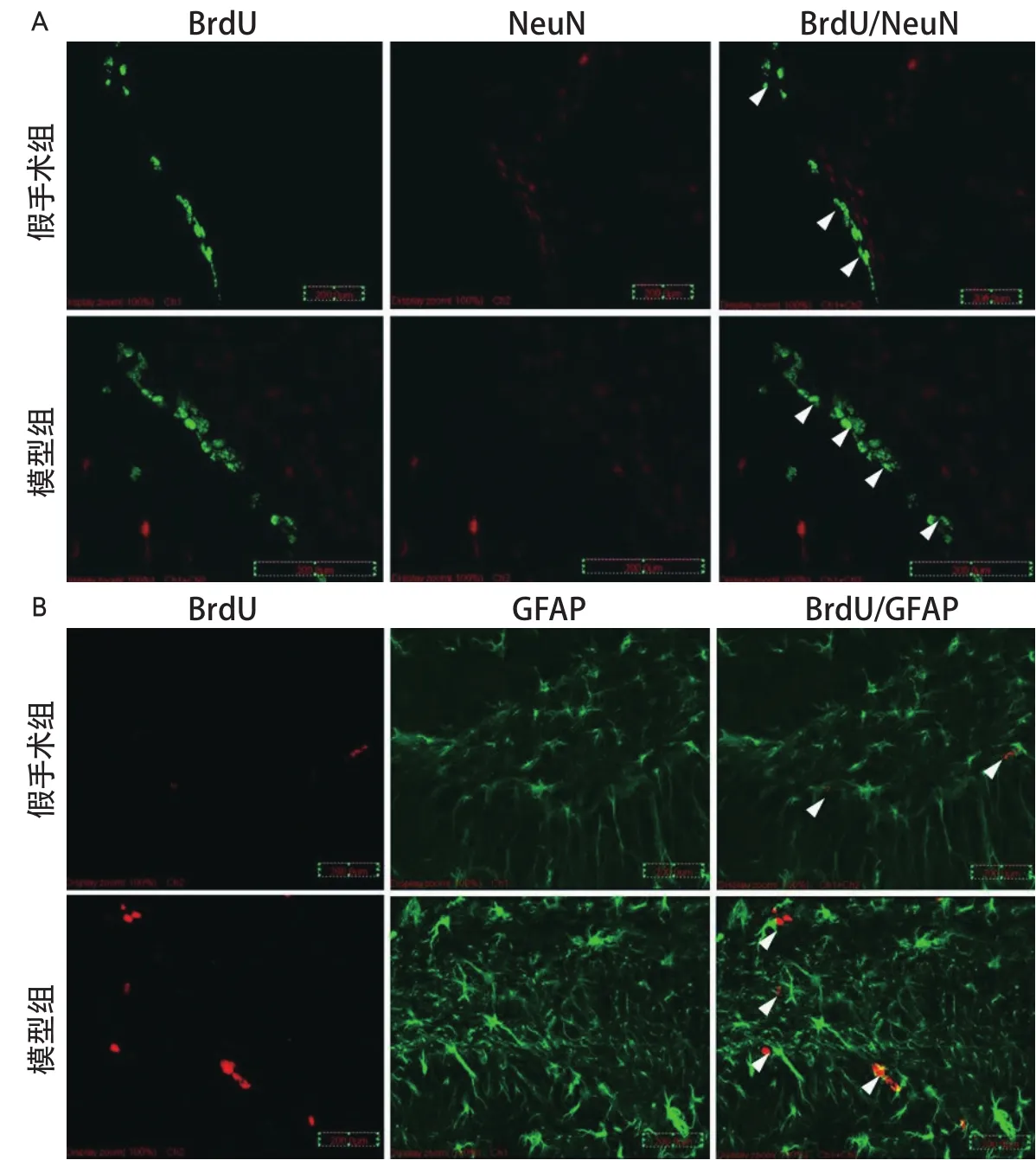
图2 术后28 d免疫荧光双标SVZ新生神经细胞与DG新生星形胶质细胞
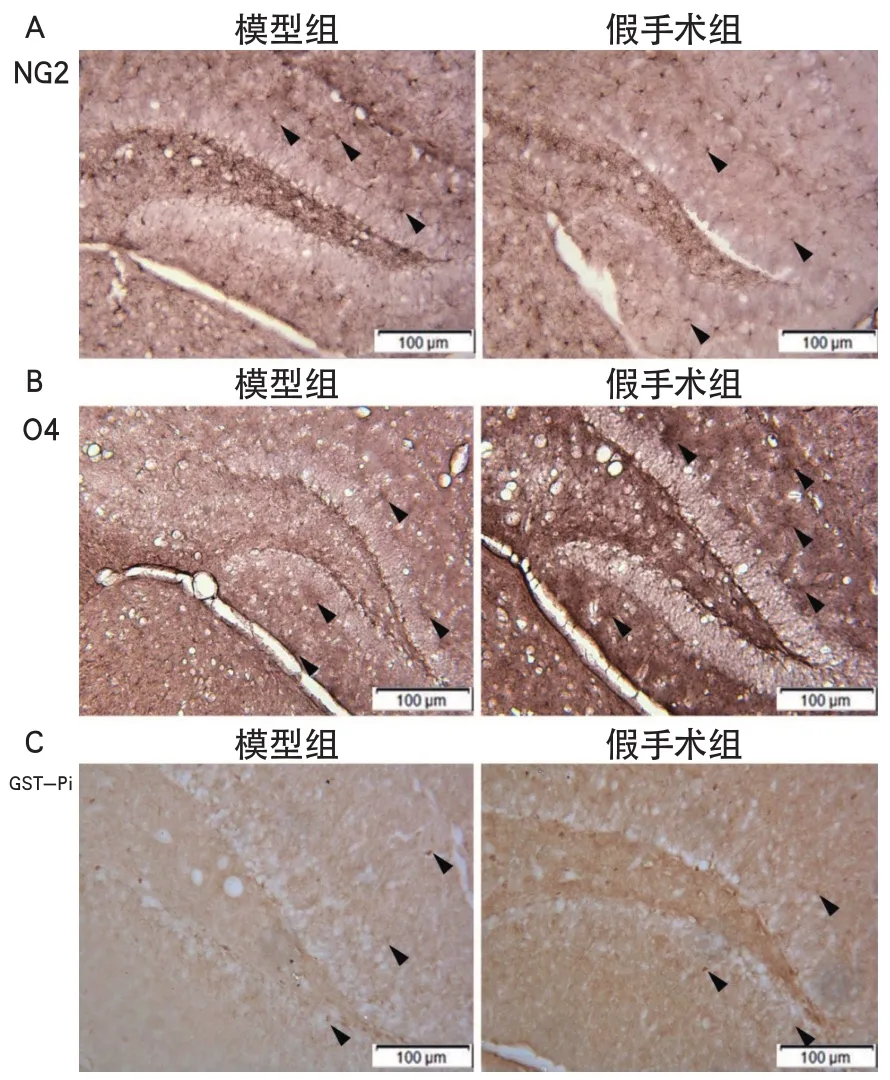
图3 免疫组织化学显示海马齿状回少突胶质细胞分化和成熟情况
3 讨论
目前,多种血管性危险因素所导致的脑血流低灌注而引起的脑深部白质损伤已被认为是血管性认知障碍的重要病理生理基础[6]。采用反复阻断双侧颈总动脉模拟脑血流量下降来制备的血管性痴呆模型具有显著的学习记忆损害[7]。我们先前的研究中发现,该模型动物术后可出现显著的学习记忆能力下降并伴有海马椎体神经元的坏死和凋亡及以少突胶质细胞死亡为表现的脑白质损伤[8]。因此,该模型是研究血管性痴呆的可靠动物模型。本研究中,我们在该模型中使用相关的神经标记物对目标细胞进行标记,通过与假手术的对照组动物进行对比,观察小鼠术后2周及4周2个神经再生区域的细胞增殖及少突胶质细胞分化成熟情况。
室管膜下区是新生儿脑内神经元、少突胶质细胞及星形胶质细胞的重要来源,近年来的研究认为成年哺乳动物脑内室管膜下区同样存在神经再生且可在多种病理条件下激活[9]。本研究中通过使用BrdU标记新生细胞发现,血管性痴呆小鼠造模后14 d及28 d后室管膜下区BrdU标记的新生细胞较假手术组增多,且术后14 d时最为显著,且同时伴有新生神经元的显著增多。表明全脑反复缺血低灌注可诱发室管膜下区新生细胞的增殖及神经元的内源性再生增强。而有关缺血性脑损伤后神经再生的一些研究显示,在大脑中动脉阻断(middle cerebral artery occlusion,MCAO)造成脑局部缺血性损伤的动物模型中,阻断供血后同样出现了室管膜下区新生细胞的显著增多,且在1~2周达到高峰[10-11]。这与本研究中室管膜下区新生细胞增殖的特点一致。但与MCAO模型不同的是,本研究仅为短暂性的缺血再灌注,手术并未导致动物出现神经功能缺损。由此可见,尽管未导致局灶性的缺血性损伤,血管性痴呆模型中全脑缺血低灌注条件同样可诱发室管膜下区内源性细胞增殖及神经元再生。研究发现,卒中导致的神经元死亡可介导内源性神经干/祖细胞分化再生为新生神经元并迁徙至损伤部位修复受损的神经元[12]。同时,室管膜下区的神经祖细胞也可以分化再生为少突胶质细胞进而修复髓鞘损伤[13]。因此,缺血低灌注所导致的以少突胶质细胞死亡及髓鞘脱失为特征性的脑白质损伤可能是诱发室管膜下区细胞增殖增强的重要因素。
有研究认为,缺血所激发的海马齿状回神经再生是对短暂全脑缺血导致的海马椎体神经元死亡的一种反应,而海马齿状回神经再生与室管膜下区新生细胞的迁徙并不一致,其在缺血性损伤中可起独立修复受损神经元的作用[14]。海马齿状回神经干/祖细胞向少突胶质细胞分化可促进脱髓鞘损伤的修复[15]。本研究对海马齿状回少突胶质细胞再生不同阶段的主要细胞进行观察后发现,模型组少突胶质细胞祖细胞增多,少突胶质细胞前体细胞减少,而成熟少突胶质细胞未见显著差异,提示海马齿状回神经干/祖细胞向少突胶质细胞祖细胞分化增加,但少突胶质细胞祖细胞向少突胶质细胞前体细胞进一步分化成熟障碍。有关血管性痴呆患者脑白质的尸检研究发现,脑白质中少突胶质细胞祖细胞增多,而成熟少突胶质细胞却显著减少,这同样提示了少突胶质细胞祖细胞向成熟少突胶质细胞分化过程可能受到了抑制[16-17]。鉴于在缺血性白质损伤的修复过程中,少突胶质细胞祖细胞聚集于神经元轴突周围并向成熟少突胶质细胞分化是髓鞘修复的关键环节[18]。因此,尽管以少突胶质细胞死亡为特征的白质损伤可通过释放多种信号分子促进内源性髓鞘再生,从而发挥自我保护和修复作用,但由于少突胶质细胞再生过程中被抑制,海马新生的少突胶质细胞祖细胞并不能进一步分化为成熟的少突胶质细胞发挥修复髓鞘损伤的作用。最新的研究发现,神经营养因子可通过促进少突胶质细胞再生来改善缺血后的髓鞘损伤,有望成为未来血管性痴呆新的治疗策略[19]。
本研究中,模型组海马齿状回新生星形胶质细胞显著增多。既往研究认为,在缺血性损伤下,血脑屏障破坏可引起星形胶质细胞的反应性增生,释放炎性因子参与缺血性炎症反应[20]。而另有研究认为从起源上讲,GFAP阳性星形胶质细胞可进一步分化形成少突胶质细胞从而参与髓鞘损伤的修复[13]。据此,我们推测,在该模型中由于海马神经干/祖细胞向少突胶质细胞再生分化抑制,因而转为分化为反应性星形胶质细胞的数量增加。因此,本研究中新生反应性星形胶质细胞的增多除了为缺血后的炎性增生外,同样也可能为机体针对缺血性白质脱髓鞘损伤所表现出的神经再生被异常调控的结果,但目前尚缺乏海马齿状回GFAP阳性星形胶质细胞在神经再生中作用的研究,相关机制有待进一步阐明。本研究在血管性痴呆动物模型水平证实了,反复缺血低灌注即可诱发室管膜下区显著的细胞增殖及神经元再生,海马齿状回的少突胶质细胞再生障碍。其中少突胶质细胞的再生障碍可影响白质髓鞘损伤的修复,这可能是血管性痴呆缺乏有效治疗的原因之一。神经干/祖细胞分化为少突胶质细胞祖细胞,再分化为少突胶质细胞的调控机制仍然不清,缺氧低灌注条件下细胞因子、酶及信号蛋白变化均可影响神经再生的强度和方向[15,18-19]。这一系列的调节通路中可能存在影响血管性痴呆神经再生修复的治疗靶点,如何对可能的靶点进行有效干预,从而促进缺血低灌注条件下白质损伤的修复,有待进一步研究。
1 贾建平,江汉秋.进一步重视血管性痴呆的防治[J].内科理论与实践,2007,2:67-69.
2 Eriksson PS,Perfilieva E,Bjork-Eriksson T,et al.Neurogenesis in the adult human hippocampus[J].Nat Med,1998,4:1313-1317.
3 Liu J,Solway K,Messing RO,et al.Increased neurogenesis in the dentate gyrus after transient global ischemia in gerbils[J].J Neurosci,1998,18:7768-7778.
4 Nait-Oumesmar B,Picard-Riera N,Kerninon C,et al.Activation of the subventricular zone in multiple sclerosis:evidence for early glial progenitors[J].Proc Natl AcadSci USA,2007,104:4694-4699.
5 Yan B,He J,Xu H,et al.Quetiapine attenuates the depressive and anxiolytic-like behavioural changes induced by global cerebral ischemia in mice[J].Behav Brain Res,2007,182:36-41.
6 Iadecola C.The pathobiology of vascular dementia[J].Neuron,2013,80:844-866.
7 靳玮.反复缺血再灌注制备小鼠血管性痴呆模型的评价[J].疑难病杂志,2008,7:131-132.
8 Yan B,Bi X,He J,et al.Quetiapine attenuates spatial memory impairment and hippocampal neurodegeneration induced by bilateral common carotid artery occlusion in mice[J].Life Sci,2007,81:353-361.
9 Braun SM,Jessberger S.Adult neurogenesis and its role in neuropsychiatric disease,brain repair and normal brain function[J].Neuropathol Appl Neurobiol,2014,40:3-12.
10 Parent JM,Vexler ZS,Gong C,et al.Rat forebrain neurogenesis and striatal neuron replacement after focal stroke[J].Ann Neurol,2002,52:802-813.
11 Komitova M,Mattsson B,Johansson BB,et al.Enriched environment increases neural stem/progenitor cell proliferation and neurogenesis in the subventricular zone of stroke-lesioned adult rats[J].Stroke,2005,36:1278-1282.
12 Arvidsson A,Collin T,Kirik D,et al.Neuronal replacement from endogenous precursors in the adult brain after stroke[J].Nat Med,2002,8:963-970.
13 Menn B,Garcia-Verdugo JM,Yaschine C,et al.Origin of oligodendrocytes in the subventricular zone of the adult brain[J].J Neurosci,2006,26:7907-7918.
14 Salazar-Colocho P,Lanciego JL,Del Rio J,et al.Ischemia induces cell proliferation and neurogenesis in the gerbil hippocampus in response to neuronal death[J].Neurosci Res,2008,61:27-37.
15 Jessberger S,Toni N,Clemenson GD,et al.Directed differentiation of hippocampal stem/progenitor cells in the adult brain[J].Nat Neurosci,2008,11:888-893.
16 Miyamoto N,Tanaka R,Shimura H,et al.Phosphodiesterase III inhibition promotes differentiation and survival of oligodendrocyte progenitors and enhances regeneration of ischemic white matter lesions in the adult mammalian brain[J].J Cereb Blood Flow Metab,2010,30:299-310.
17 Ekonomou A,Ballard CG,Pathmanaban ON,et al.Increased neural progenitors in vascular dementia[J].Neurobiol Aging,2011,32:2152-2161.
18 Mifsud G,Zammit C,Muscat R,et al.Oligodendrocyte pathophysiology and treatment strategies in cerebral ischemia[J].CNS Neurosci Ther,2014,20:603-612.
19 Ramos-Cejudo J,Gutierrez-Fernandez M,Otero-Ortega L,et al.Brain-derived neurotrophic factor administration mediated oligodendrocyte differentiation and myelin formation in subcortical ischemic stroke[J].Stroke,2015,46:221-228.
20 Simpson JE,Fernando MS,Clark L,et al.White matter lesions in an unselected cohort of the elderly:astrocytic,microglial and oligodendrocyte precursor cell responses[J].Neuropathol Appl Neurobiol,2007,33:410-419.
猜你喜欢
中南药学(2022年2期)2022-11-14
昆明医科大学学报(2021年4期)2021-07-23
大自然探索(2019年9期)2019-12-13
中国现代药物应用(2019年16期)2019-09-06
中国临床医学影像杂志(2019年4期)2019-06-18
中国临床医学影像杂志(2019年2期)2019-04-25
中医眼耳鼻喉杂志(2019年3期)2019-04-13
中成药(2017年6期)2017-06-13
中国介入影像与治疗学(2017年4期)2017-04-24
转化医学电子杂志(2017年7期)2017-01-12
