
7种水稻育种亲本对福建省稻瘟病菌的抗病性分析
2015-01-06 00:38杜宜新石妞妞阮宏椿杨秀娟陈福如
西北农林科技大学学报(自然科学版) 2015年12期
杜宜新,石妞妞,阮宏椿,甘 林,杨秀娟,陈福如
(福建省农业科学院 植物保护研究所,福建 福州 350013)
7种水稻育种亲本对福建省稻瘟病菌的抗病性分析
杜宜新,石妞妞,阮宏椿,甘 林,杨秀娟,陈福如
(福建省农业科学院 植物保护研究所,福建 福州 350013)
【目的】 评价12S、M76A、闽香A、明兴A、延源A、福丰5A和福香1A 7种水稻育种亲本对福建省稻瘟病的抗病性,为水稻抗病育种及抗病品种合理布局提供参考。【方法】 采用田间自然诱发的方法,测定7种供试水稻育种亲本对水稻叶瘟和穗颈瘟的抗病性;并采用室内人工接种法,测定7种供试水稻育种亲本对水稻苗瘟的抗病性及对福建省稻瘟病菌4个主要致病型的抗性频率。【结果】 M76A、明兴A、延源A、福丰5A及福香1A对苗瘟的抗性频率均大于75%,其中M76A、福丰5A、福香1A对叶瘟、穗颈瘟也均表现出较高的抗病性,而明兴A、延源A较感叶瘟和穗颈瘟,12S、闽香A较感苗瘟、叶瘟和穗颈瘟;明兴A、延源A、福丰5A对福建省稻瘟病菌4个主要致病型 I34.1、I24.1、I36.1和I35.1菌株的抗性频率均较高,而M76A、福香1A分别对I24.1和 I36.1菌株的抗性频率较低,12S对I34.1和I36.1 2个致病型菌株的抗性频率均较低,闽香A对I34.1、I36.1和I35.1 3个致病型菌株的抗性频率均较低。【结论】 M76A、福丰5A和福香1A对福建省稻瘟病的抗病性较强,可作为抗瘟育种的候选亲本;明兴A、延源A、12S和闽香A则应与抗病性较强的恢复系杂交育种。
稻瘟病菌;抗性频率;抗瘟基因;育种亲本
由稻瘟病菌(MagnaporthegriseaBarr.)引起的稻瘟病是水稻的主要病害,其严重阻碍水稻高产、稳产。选育和利用水稻抗病品种是防治稻瘟病最经济有效的措施,而发掘和利用水稻抗性遗传资源和抗性基因对水稻抗稻瘟病育种具有重要意义[1]。水稻品种的抗瘟性大多数由1~2对显性基因控制,这就意味着只要杂交稻双亲之一具有抗瘟性,其相应的杂交稻F1代就可能具有抗瘟性,杂交稻组合的抗瘟性受不育系和恢复系的影响,不育系的抗瘟性可通过核质遗传给F1代,恢复系的抗瘟性可通过细胞核遗传给F1代[2-4]。因此,明确水稻育种亲本对区域稻瘟病菌群体的抗瘟性,是合理使用抗瘟亲本、选育优良抗瘟品种的重要基础。本研究测定了7种水稻育种亲本(不育系)对福建省稻瘟病菌不同致病型的抗病性及其在田间自然诱发条件下的抗病性,旨在明确这些亲本作为抗性育种材料的利用价值,为水稻抗病品种的选育和水稻稻瘟病的控制提供科学依据。
1 材料与方法
1.1 供试材料
2012-2014年,以普感稻瘟病的水稻品种丽江新团黑谷为诱发品种,将其种植于闽东、闽南、闽西、闽北和闽中5个主要稻区,采集其稻瘟病标本,单孢分离,随机选取其中176个单孢菌株作为供试菌株。
供试7种水稻育种亲本为12S、M76A、闽香A、明兴A、延源A、福丰5A和福香1A。
供试鉴别用水稻品种为CO39近等基因系(Near-isogenic lines,NILs),其品种及所携带的抗瘟基因(括号内为抗瘟基因)为C101A51(Pi-2)、C101LAC[Pi-1(2)]、C101PKT(Pi-4a)、C105TTP-4-L-23(Pi-4b)、C104PKT[Pi-3(2)]和CO39[Pi-a(2)]。
供试单基因水稻品种为IRRI-日本合作育成的24个水稻抗稻瘟病单基因系,其品种及携带的抗病基因(括号内为抗瘟基因)为IRBLkm-Ts(Pi-km)、IRBL7-M[Pi-7(t)]、IRBLkp-K60(Pi-kp)、IRBL9-W[Pi-9(t)]、IRBLkh-K3(Pi-kh)、IRBLk-Ka(Pi-k)、IRBLz5-CA(Pi-z5)、IRBLta2-Pi[Pi-ta(1)]、IRBL1-CL[Pi-1(1)]、IRBL20-IR24(Pi-20)、IRBLzt-T(Pi-zt)、IRBLta-K1[Pi-ta(2)]、IRBL11-Zh(Pi-11)、IRBLi-F5(Pi-i)、IRBLb-B(Pi-b)、IRBL5-M[Pi-5(t)]、IRBL12-M[Pi-12(t)]、IRBLa-A[Pi-a(1)]、IRBLsh-B(Pi-sh)、IRBLz-Fu(Pi-z)、IRBL3-CP4[Pi-3(1)]、IRBLt-K59(Pi-t)、IRBL19-A[Pi-19(t)]、IRBLks-F5(Pi-ks),均由中国农业大学彭友良教授转赠。
1.2 方 法
1.2.1 稻瘟病菌孢子悬浮液的制备 将保存于滤纸片上的供试菌株转至淀粉培养基(可溶性淀粉10 g、酵母浸膏2 g、碳酸钙3 g、琼脂18 g、水1 000 mL),28 ℃培养7 d,再转接至米糠培养基(米糠20 g、琼脂20 g、水1 000 mL),28 ℃培养10 d,然后用已灭菌的细毛笔刮除气生菌丝,置于30 W黑光灯下,在28 ℃、空气相对湿度85%~90%条件下诱导产孢,3 d后用无菌水洗下分生孢子,用0.1%吐温80水溶液配制成1.0×105CUF/mL的孢子悬浮液,备用[5]。
1.2.2 供试水稻育种亲本对叶瘟和穗颈瘟的抗病性测定 供试水稻育种亲本对叶瘟、穗颈瘟的抗病性鉴定采用田间自然诱发的方法,选择福建省稻瘟病发生较重的水稻主栽区上杭县茶地乡和建阳市麻沙镇设置田间鉴定病圃。所选田块排灌方便,肥力中等。每品种种植1小区,小区面积2 m2,株行距15 cm×20 cm,小区随机排列,小区四周设置诱发品种保护行,诱发品种选用汕优63,试验设置3次重复。肥水管理同常规生产,防治虫害不防治病害,于诱发品种的叶瘟或穗颈瘟发病稳定时进行发病程度调查[6-7]。病情指数平均值0为高抗(HR),0.1%~5%为抗(R),5.1%~15%为中抗(MR),15.1%~25%为中感(MS),25.1%~50%为感(S),≥50.1%为高感(HS)[8]。
1.2.3 供试水稻育种亲本对苗瘟和稻瘟菌主要致病型菌株的抗病性测定及其携带抗瘟基因的推导 供试水稻育种亲本对苗瘟、稻瘟病菌主要致病型菌株的抗病性测定及其携带的抗病基因推导均采用温室内人工接种的方法。将供试的水稻育种亲本、CO39近等基因系品种及供试单基因品种的水稻种子催芽后播种于50孔塑料育秧盘内,育秧盘孔尺寸为5 cm×5 cm×5 cm,每个品种播种3孔,每盘播种16个水稻品种,每孔播种约10粒,播种后将育秧盘置于可控温、控湿的温室水泥池中,待水稻秧苗生长至3叶1心时分别喷雾接种176个稻瘟病菌孢子,接种后保湿48 h,7 d后调查病情[9]。病情调查参照周江鸿等[10]的方法。
(1)单个水稻品种抗性频率的计算。单个水稻品种的抗性频率=(对单个水稻品种无毒的菌株数/总菌株数)×100%[9]。
(2)对稻瘟病菌主要致病型的抗病性。稻瘟病菌致病型划分按Gilomour[11]的八进制法进行编码,其中CO39近等基因系的6个鉴别品种C101A51、C101LAC、C101PKT、C105TTP-4-L-23、C104PKT和CO39分别被赋值为1,2,4,10,20和0.1[12]。利用CO39近等基因系的6个品种对176株稻瘟病菌的致病型进行鉴别,并进一步统计7种水稻育种亲本对主要致病型的抗性频率。
(3)抗病基因推导。根据基因对基因假说[13-14],通过单基因水稻品种对供试菌株的抗病性,来推导供试菌株携带的无毒基因;再根据7个水稻育种亲本对供试菌株的感病性,推导出供试水稻育种亲本未携带的供试抗瘟基因,反之,根据水稻育种亲本对供试菌株的抗病性,推导出供试水稻育种亲本可能携带的供试抗瘟基因。
2 结果与分析
2.1 供试水稻育种亲本对福建省稻瘟病的抗病性
由表1可知,供试7种水稻育种亲本对176个稻瘟病菌(苗瘟)的抗病性频率均大于60%,其中M76A、明兴A、延源A、福丰5A及福香1A对供试菌株的抗性频率较高,均大于75%。M76A、福丰5A和福香1A在建阳市麻沙镇、上杭县茶地乡两地对叶瘟和穗颈瘟均表现出较高的抗病性,有较好的利用价值;明兴A在建阳市麻沙镇对叶瘟的抗病性较差,在建阳市麻沙镇和上杭县茶地乡两地对穗颈瘟均表现感病;延源A在建阳市麻沙镇对叶瘟的抗病性较差,在建阳市麻沙镇、上杭县茶地乡两地对穗颈瘟分别表现为高感和感病;12S和闽香A苗期对供试菌株抗病性较差,在田间叶瘟和穗颈瘟的病情指数也较高,作为育种材料时应考虑与抗病性较好的恢复系亲本材料杂交使用。
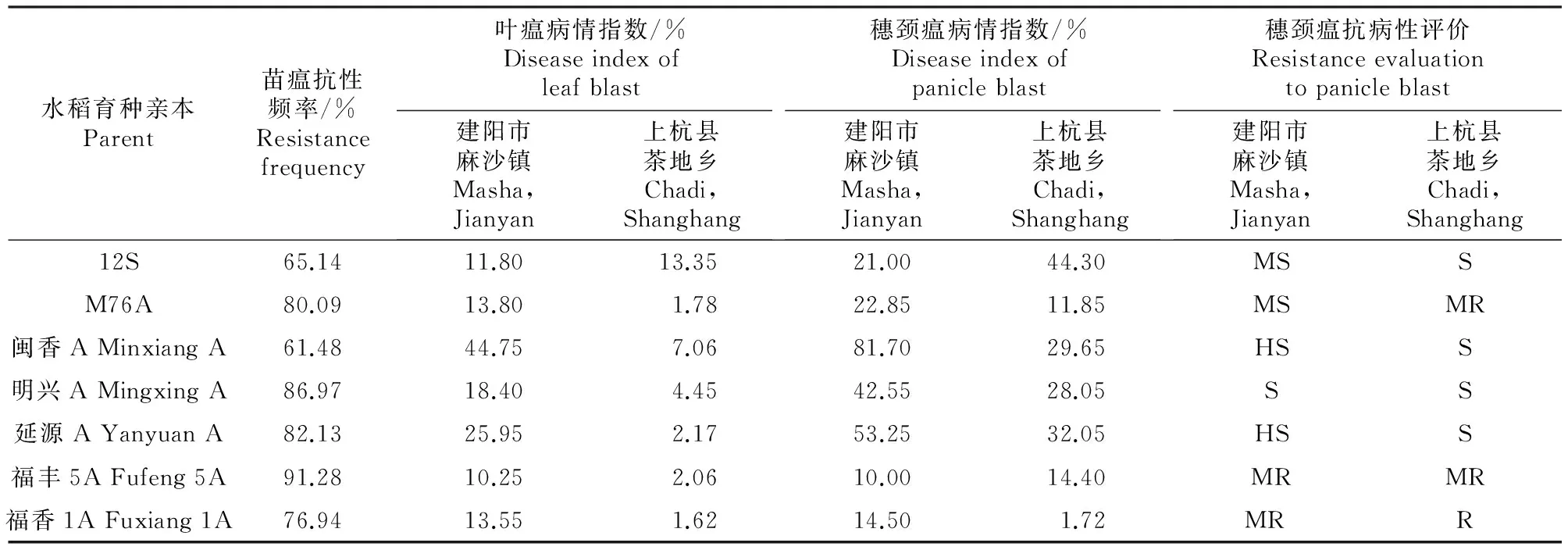
表1 7种水稻育种亲本对福建省稻瘟病菌的抗病性Table 1 Resistances of 7 rice breeding parents to Magnaporthe grisea isolates in Fujian
2.2 供试水稻育种亲本对福建省稻瘟病菌主要致病型的抗病性
利用CO39近等基因系鉴别品种可以将供试的176株稻瘟病菌鉴定为16个致病型,其中I34.1,I24.1,I36.1和I35.1为4个主要致病型,出现频率分别为40.34%,13.64%,11.93%和10.23%。由表2可知,12S对I34.1和I36.1 2个致病型菌株的抗性频率均较低,分别为60.56%和50.00%;M76A对I24.1菌株的抗性频率较低;闽香A对I34.1,I36.1和I35.1 3个致病型菌株的抗性频率均较低;福香1A对I36.1菌株的抗性频率较低;而明兴A、延源A和福丰5A对4个主要致病型菌株均有较高的抗性频率。
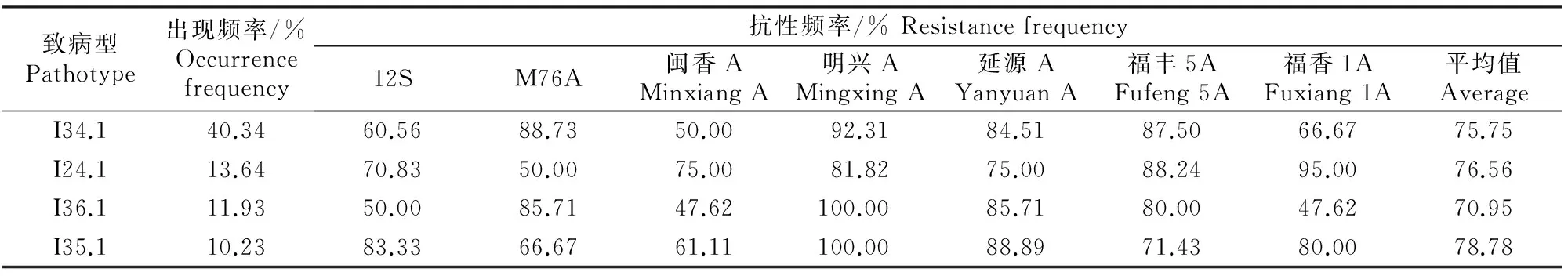
表2 7种水稻育种亲本对福建省稻瘟病菌4个主要致病型的抗性频率Table 2 Resistance frequencies of 7 rice breeding parents to four main pathotypes of Magnaporthe grisea in Fujian
2.3 供试水稻育种亲本抗病基因推导
由表3可知,供试7种水稻育种亲本中,闽香A不含有供试抗瘟基因中的任何一个基因,12S、福香1A则可能分别含有Pi-h和Pi-4b,肯定不含有供试的其他抗瘟基因;M76A、明兴A、延源A和福丰5A则可能含有3个及以上的供试抗瘟基因,这与其对供试稻瘟病菌的抗性频率吻合。

表3 7种水稻育种亲本可能携带的抗瘟基因Table 3 Postulated resistance genes of seven rice breeding parents
3 讨 论
张长伟等[3]研究表明,杂交稻抗瘟性受加性和非加性遗传的共同控制,但以加性效应为主,源于恢复系的加性效应明显比不育系的加性效应重要。然而,不育系作为杂交稻的重要亲本,对杂交水稻新品种(组合)的培育起着十分关键的作用,不育系有更多的必要性状要考虑,如不育性、异交习性、稻米品质等,其选育难度更大,周期更长,因而育种上对不育系抗病性的改良研究也较恢复系滞后[4,15]。本研究表明,供试的7种水稻育种不育系亲本对福建省稻瘟病菌均有一定的抗性频率,其中M76A、明兴A、延源A、福丰5A及福香1A抗性频率较高,并且M76A、福丰5A及福香1A在田间自然诱发的条件下对叶瘟和穗颈瘟也有较高的抗病性,可作为抗瘟育种的候选亲本。明兴A和延源A对供试菌株的抗性频率也较高,并且对供试水稻育种亲本可能携带的抗瘟基因推导结果也表明,明兴A对携带Avr-ta(1)、Avr-a(2)、Avr-h无毒基因的菌株表现为抗性,延源A则对携带Avr-5(t)、Avr-ta(1)、Avr-ks、Avr-3(1)、Avr-z、Avr-3(2)、Avr-20、Avr-t、Avr-i、Avr-4b、Avr-2、Avr-b、Avr-12(t)、Avr-4a等多个无毒基因的菌株表现为抗病,但在建阳市麻沙镇、上杭县茶地乡两地田间自然诱发条件下则较感穗颈瘟,这可能与建阳市麻沙镇、上杭县茶地乡两地稻瘟病菌群体毒力组成有关,因此仍要考虑与抗病性较强的恢复系杂交育种。
稻瘟病菌群体毒力结构变化,是导致水稻抗病品种抗病性降低和丧失的主要原因之一,稻瘟病菌种群的毒力影响田间主栽品种的抗病性,同时田间主栽品种的布局又影响稻瘟病菌稳定化的选择,从而影响区域稻瘟病菌种群组成,两者相互选择、密切相关[16],因此明确区域稻瘟病菌种群的毒力组成,对水稻抗病品种推广、区域抗病品种布局尤为重要,而供试稻瘟病菌的样本选择是研究稻瘟病菌群体毒力组成的关键。研究表明,丽江新团黑谷具有普感性[17],本研究以丽江新团黑谷作为诱发品种采集分离的稻瘟病菌群体,能更真实地反应稻瘟病菌自然种群的毒力结构。供试的7种水稻育种亲本对 I34.1、I24.1、I36.1和I35.1 4个主要致病型的抗病性存在较大差异,如12S对致病型I36.1的抗性频率较低,仅为50.00%,而对I35.1的抗性频率则较高,为83.33%;同样,M76A对致病型I24.1和 I35.1、闽香A对致病型I34.1、I36.1和I35.1,福香1A对致病型I34.1和I36.1的抗性频率均较低,因此在水稻抗病育种及抗病品种推广、区域抗病品种布局上,应密切监测不同区域稻瘟病菌群体的致病型组成及其毒力结构变化,防止优势致病型的过速更替,延长抗病品种种植年限,预防稻瘟病的爆发流行。
[1] 张学博.应用抗病品种防治稻瘟病的研究 [J].福建农学院学报,1981,10(2):23-29.
Zhang X B.Studies on the use of resistant varieties in controlling rice blast disease [J].Journal of Fujian Agricultural College,1981,10(2):23-29.(in Chinese)
[2] 梁 斌,余腾琼,徐福荣,等.云南省3个地方稻种的抗稻瘟病性遗传分析 [J].中国农业科学,2002,35(7):784-788.
Liang B,Yu T Q,Xu F R,et al.Genetic analysis of three Yunnan rice varieties for resistance to rice blast [J].Scientia Agricultura Sinica,2002,35(7):784-788.(in Chinese)
[3] 张长伟,郑家奎,蒋开峰,等.杂交水稻叶瘟抗性杂种优势的遗传分析 [J].植物病理学报,2000,30(1):7-12.
Zhang C W,Zheng J K,Jiang K F,et al.Genetic analysis for heterosis of resistance to leaf blast in hybrid rice [J].Acta Phytopathologica Sinica,2000,30(1):7-12.(in Chinese)
[4] 黄 富,谢 戎,刘成元,等.亲本抗瘟性对杂交水稻组合抗瘟性的影响 [J].杂交水稻,2007,22(2):64-68.
Huang F,Xie R,Liu C Y,et al.Effects of blast resistance of parents on that of their F1hybrids in rice [J].Hybrid Rice,2007,22(2):64-68.(in Chinese)
[5] 杜宜新,李 科,阮宏椿,等.稻瘟病菌对稻瘟灵、异稻瘟净和三环唑的敏感性研究 [J].植物保护学报,2011,38(5):455-460.
Du Y X,Li K,Ruan H C,et al.Sensitivities ofMagnaporthegrisesto isprothiolane,iprobenfos and tricyclazole [J].Journal of Plant Protection,2011,38(5):455-460.(in Chinese)
[6] 杜宜新,阮宏椿,王茂明,等.福建省水稻主栽品种对稻瘟病的抗性评价 [J].中国农学通报,2010,26(7):217-221.
Du Y X,Ruan H C,Wang M M,et al.Resistance evaluation of leading rice varieties to rice blast disease in Fujian province [J].Chinese Agricultural Science Bulletin,2010,26(7):217-221.(in Chinese)
[7] 张 舒,陈其志,吕 亮,等.自然诱发条件下湖北省水稻主栽品种对稻瘟病、纹枯病的抗性鉴定 [J].华中农业大学学报,2006,25(3):236-240.
Zhang S,Chen Q Z,Lü L,et al.Assessment of the variety resistance toPyriculariagriseaandRhizoctoniasolaniinduced under the natural condition in Hubei province [J].Journal of Huazhong Agricultural University,2006,25(3):236-240.(in Chinese)
[8] 杨秀娟,朱春雨,阮宏椿,等.福建省稻瘟病菌毒性类型及部分水稻品种(组合)抗病性 [J].福建农林大学学报:自然科学版,2008,37(3):244-247.
Yang X J,Zhu C Y,Ruan H C,et al.Pathogenic types ofMagnaporthegriseaBarr and the resistance of some rice cultivars to the pathogens in Fujian province [J].Journal of Fujian Agriculture and Forestry University:Natural Science Edition,2008,37(3):244-247.(in Chinese)
[9] 兰 波,杨迎青,徐沛东,等.水稻主要抗瘟基因品系对江西省稻瘟病菌分离株系的抗性分析 [J].植物保护学报,2014,41(2):163-168.
Lan B,Yang Y Q,Xu P D,et al.Analysis of the resistance of rice majorPi-genes to the isolates in Jiangxi province [J].Journal of Plant Protection,2014,41(2):163-168.(in Chinese)
[10] 周江鸿,王久林,蒋琬如,等.我国稻瘟病菌毒力基因的组成及其地理分布 [J].作物学报,2003,29(5):646-651.
Zhou J H,Wang J L,Jiang W R,et al.Virulence genes diversity and geographic distribution ofPyriculariagriseain China [J].Acta Agronomica Sinica,2003,29(5):646-651.(in Chinese)
[11] Gilomour J.Octal notation for designating physiologic races of plant pathogen [J].Nature,1973,242:620.
[12] 凌忠专,T New,王久林,等.中国水稻近等基因系的育成及其稻瘟病菌生理小种鉴别能力 [J].中国农业科学,2000,33(4):1-8.
Ling Z Z,New T,Wang J L,et al.Development of Chinese near-isogenic lines of rice and their differentiating ability to pathogenic race ofPyriculariagrisea[J].Scientia Agricultura Sinica,2000,33(4):1-8.(in Chinese)
[13] Flor H H.Current status of the genes for gene concept [J].Ann Rev Phytopath,1971,9:275-296.
[14] Silue D,Tharreau D,Notteghem J L.Evidence of a gene-for-gene relationship in theOryzasativa-Magnaporthegriseapathosystem [J].Phytopathology,1992,82:577-580.
[15] 韦新宇,许旭明,张 锐,等.籼粳交新种质康丰A对稻瘟病抗性的遗传 [J].植物遗传资源学报,2014,15(5):1133-1137.
Wei X Y,Xu X M,Zhang R,et al.Inheritance of blast resistance in new germplasm kangfeng A from indica-japonica crosses [J].Journal of Plant Genetic Resources,2014,15(5):1133-1137.(in Chinese)
[16] 杨 静,何霞红,王云月,等.水稻遗传多样性田间稻瘟病菌生理小种研究 [J].植物病理学报,2004,34(5):468-471.
Yang J,He X H,Wang Y Y,et al.Study on constitution of physiological race ofMagnaporthegriseain rice mixture field [J].Acta Phytopathologica Sinica,2004,34(5):468-471.(in Chinese)
[17] 雷财林,王久林,程治军,等.水稻抗稻瘟病单基因鉴别体系研究的概况与展望 [J].作物杂志,2014(2):5-8.
Lei C L,Wang J L,Cheng Z J,et al.Advances in development of rice monogenic differentials for blast resistance [J].Crops,2014(2):5-8.(in Chinese)
Resistances of seven rice breeding parents toMagnaporthegriseain Fujian
DU Yi-xin,SHI Niu-niu,RAUN Hong-chun,GAN Lin,YANG Xiu-juan,CHEN Fu-ru
(InstituteofPlantProtection,FujianAcademyofAgriculturalSciences,Fuzhou,Fujian350013,China)
【Objective】 This study evaluated the resistances of seven rice breeding parents (12S,M76A,Minxiang A,Mingxing A,Yanyuan A,Fufeng 5A and Fuxiang 1A) toMagnaporthegriseato provide references for breeding and distribution of resistant rice cultivars.【Method】 The resistances of seven tested rice breeding parents to rice leaf blast and rice panicle blast were also determined under nature induction condition.The resistances of seven tested rice breeding parents to rice seedling blast and their resistance frequencies to four main pathotypes ofM.griseawere assayed by artificial inoculation in greenhouse.【Result】 The resistance frequencies of M76A,Mingxing A,Yanyuan A,Fufeng 5A and Fuxiang 1A toM.griseawere all above 75%.M76A,Fufeng 5A and Fuxiang 1A also showed high resistance to rice leaf blast and rice panicle blast.Mingxing A and Yanyuan A were susceptible to both rice leaf blast and rice panicle blast,and 12S and Minxiang A were susceptible to rice seedling blast,rice leaf blast and rice panicle blast.Mingxing A,Yanyuan A and Fufeng 5A possessed high resistance frequencies to four strains (I34.1,I24.1,I36.1 and I35.1) of main pathotypes in Fujian,while M76A and Fuxiang 1A showed low resistance frequencies to strains I24.1 and I36.1,respectively.12S showed low resistance frequencies to strains I34.1 and I36.1 and Minxiang A showed low resistance frequencies to strains I34.1, I36.1 and I35.1.【Conclusion】 The resistances of M76A,Fufeng 5A and Fuxiang 1A toM.griseain Fujian were strong and they could be used as candidate parents for rice breeding.Mingxing A,Yanyuan A,12S and Minxiang A should be hybridized with restorer lines that had strong resistance toM.grisea.
Magnaporthegrisea;resistance frequency;Pi-gene;breeding parent
时间:2015-11-11 16:16
10.13207/j.cnki.jnwafu.2015.12.016
2015-06-05
公益性行业(农业)科技专项(201203014);福建省省属公益类科研院所基本科研专项(2015R1024-1);福建省科技厅项目(2009N2005)
杜宜新(1979-),男,山东济宁人,硕士,助理研究员,主要从事植物真菌病害及其防治研究。 E-mail:yixindu@163.com
陈福如(1961-),男,福建莆田人,研究员,主要从事植物真菌病害研究。E-mail:chenfuruzb@163.com
S511.037;S435.111.4+1
A
1671-9387(2015)12-0107-05
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20151111.1616.032.html
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
热带作物学报(2022年7期)2022-08-06
西南农业学报(2022年5期)2022-06-06
作物学报(2022年6期)2022-04-08
作物学报(2022年4期)2022-02-10
中国糖料(2021年3期)2021-07-13
农民文摘(2019年7期)2019-01-14
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
中国蔬菜(2015年9期)2015-12-21
