
陕北小流域生物结皮空间分布影响因子的通径分析
2015-01-03 06:30:02张振师卜崇峰
水土保持研究 2015年6期
袁 方,张振师,张 朋,卜崇峰,2
(1.西北农林科技大学 水土保持研究所,陕西 杨凌712100;2.中国科学院水利部水土保持研究所,陕西 杨凌712100;3.中国电建集团西北勘测设计研究院有限公司,西安710065)
生物结皮(Biological soil crusts)是土壤颗粒与微生物、藻类、地衣以及苔藓等组成的复杂聚合体,是干旱半干旱广泛存在的活性地被物[1]。它的存在可以富集土壤养分,提高土壤抗蚀性[2],降低土壤的风蚀、水蚀量,影响水分再分配[3-4],具有重要的生态功能[5]和良好的水土保持作用。通径分析是简单相关分析的继续,是把自变量与因变量之间的相关性分解为该自变量对因变量的直接影响和通过其他相关自变量对因变量间接影响的分析方法[6]。其不仅具有回归与相关分析的作用,且进一步地将自变量与因变量的相关统计量剖分为直接通径系数和间接通径系数,揭示影响现象变异的主要因素所产生的直接和间接作用,从而区分自变量的相对重要性及其与因变量的关系,为研究和分析的问题提供更全面更完善的决策依据[7]。前人对生物结皮空间分布及其影响因子的研究大多通过野外调查,对所得各类指标进行直观的统计分析[8-9],但鲜有学者利用通径分析研究各因子对生物结皮的直接或间接影响及其作用路径。
黄土高原水蚀风蚀交错区位于黄土高原北部,属于干旱半干旱地区草原带[10],是黄土高原水土流失最为严重的地区[11],其环境条件类型复杂多变、景观破碎,水分条件比较恶劣。自退耕还林(草)工程实施以来,黄土高原地区生物结皮大面积发育[8],其在防治水土流失和促进生态演替方面作用显著。而生物结皮在自然条件下的发育十分缓慢,容易受到环境因素(土壤、水分条件、植被、海拔、坡向等[12-13])和人类干扰活动[14]的影响,且各影响因素对生物结皮的影响程度及作用存在差异,故通过生物结皮生长状况的野外调查,了解生物结皮的空间分布规律及其影响因子,明确各影响因子的具体作用路径,以期为更好地促进生物结皮的形成和发育奠定基础。
为此,本研究选取黄土高原北部水蚀风蚀交错区六道沟小流域的一处完整梁峁坡面,通过全面调查和测算分析,应用SPSS 12.0及 Excel 2007软件,采用通径分析的方法研究生物结皮空间分布与环境因子之间的关系,旨在为野外生物结皮发育的环境评价和快速培育提供理论支持。
1 材料与方法
1.1 研究区概况
研究区位于陕西省神木县以西14km处的六道沟小流域(110°21′—110°23′E,38°46′—38°51′N),海拔1 094~1 274m,流域面积6.89km2,主沟道南北走向,长4.2 km,属窟野河二级支流。地处黄土高原向毛乌素沙地、森林草原向干旱草原的过渡地带,属水蚀作用向风蚀作用过渡的典型水蚀风蚀交错带,年侵蚀模数达10 000t/km2[15]。气候属中温带半干旱气候,冬春季干旱少雨多风沙,风蚀严重,夏秋多雨且暴雨频繁,水蚀强烈。年均降雨量400mm,其中6—9月份的降雨量约占全年的70%~80%,年均气温7~9℃,盛行西北风,年平均风速为2.2m/s[16]。流域内的主要植被有柠条(Caragana korshinskiiKom.)、长芒草(StipabungeanaTrin.)、苜蓿(MedicagosativaLinn)、达 乌 里 胡 枝 子 (Lespedeza davurica(Laxm.)Schindl.)、阿尔泰狗娃花(Heteropappusaltaicus(Willd)Novopokr)和茭蒿(ArtemisiagiraldiiPamp)等。流域东侧主要为黄土覆盖,占流域面积的86.5%;西侧多为固定沙丘,占流域面积的13.5%[17]。
1.2 研究方法
本研究在流域内选定一处撂荒30a以上的典型的梁峁坡面,于2014年6—10月开展生物结皮空间分布的系统调查工作。其中于6月,7月份对生物结皮的盖度、厚度和抗剪强度进行调查。利用etrex HD型手持GPS对野外每个调查点进行定位,利用方格网法测算该点周围半径10m左右范围内生物结皮的盖度(%),厚度(mm)用游标卡尺进行原位测定,重复3~5次;抗剪强度(kPa)用袖珍剪力仪(BWT2XZJL)原位测定,重复3~5次;于8月中旬到9月上旬对植被类型和植被盖度进行调查;于10月上旬(雨季末)在调查区内选择几处典型样地,对样地内的土壤进行采样分析,取表层0—5cm土层,带回室内105℃下烘24h,测算土壤含水率(%)。利用GIS插值功能将所有野外调查点及调查数据进行制图,共得到1 342个样本的结皮盖度和厚度、地形、土壤、植被等指标。
数据的统计分析利用SPSS 12.0及 Excel 2007软件进行通径分析。通径分析是研究多个自变量与因变量间多重线性关系的一种重要的统计方法,是相关分析和回归分析的结合,将自变量与因变量的简单相关系数分解为直接通径系数和间接通径系数,分别表示某一自变量对因变量的直接影响及通过其他自变量对因变量的间接影响,决定系数表示自变量对因变量的决定程度,通过比较各变量的决定系数大小,对各变量的作用进行排序,为统计决策提供更全面更完善的依据[18];袁志发等[19]提出决策系数的概念,其反映了自变量通过其他自变量的相关网对因变量的综合决定作用,指出在复杂的信息路径中,选择什么样的路径对因变量最好。
2 结果与分析
2.1 生物结皮的空间分布概况
生物结皮盖度的空间分布具有明显的分异性和连续性。结皮的平均盖度在30%以上,主要分布在沙土区,呈成片的连续分布;而在黄土区生物结皮则呈离散的零星分布,盖度大多在20%以下,且主要分布在坡地的边缘和末端。沙土区生物结皮的空间分布具有明显的发源地,大都处在沙土和黄土交界处的低洼处。相对来说,生物结皮的厚度和抗剪强度的空间变异性却并不大。沙土区和黄土区生物结皮的平均厚度分别为12.8mm和13.1mm,平均抗剪强度分别为4.76kg/cm2,4.96kg/cm2。t检验结果表明:黄土区生物结皮的厚度及平均抗剪强度和沙土区的差异并不显著(p>0.05)。
2.2 生物结皮盖度与环境因子间的关系
黄土高原水蚀风蚀交错区复杂的环境特征影响着生物结皮的生长发育,结皮盖度是描述生物结皮发育状况的一个重要指标。为研究结皮盖度与环境因子之间的关系,根据逐步回归结果,选取沙土、沉积物运移指数、坡向、沙蒿盖度、海拔及土壤含水率作为影响因变量生物结皮盖度的自变量,进行相关及通径分析。
2.2.1 生物结皮盖度及各环境因子的相关分析对结皮盖度及沙土、沉积物运移指数、坡向、沙蒿盖度、海拔及土壤含水率采用SPSS 12.0软件进行相关分析,得出上述环境因子与结皮盖度的相关及回归系数(表1)。

表1 环境因子与结皮盖度的相关与回归分析结果
由表1可以看出:6个环境因子与结皮盖度相关系数的绝对值大小依次为:沙土>沙蒿盖度>海拔>坡向>沉积物运移指数>土壤含水率,且沙土和沙蒿盖度与结皮盖度呈正相关关系,其余均与结皮盖度呈负相关关系,除土壤含水率以外各环境因子均与结皮盖度的相关性达到极显著水平(p<0.01)。此外,各环境因子之间也存在显著的相关性,即共线性;各环境因子与结皮盖度之间标准化回归方程达到极显著水平(p<0.01),R2=0.706,这说明结皮盖度关于上述6个环境因子的通径分析是必要的。
2.2.2 生物结皮盖度及各环境因子的通径分析 表2和图1a展示了不同环境因子对结皮盖度的直接和间接影响。从表2可以看出:(1)沙土对结皮盖度的直接影响最大(0.720),土壤含水率(0.526)及沙蒿盖度(0.235)次之,即从直接影响看沙土、土壤含水率及沙蒿盖度是影响结皮盖度的主要因子。(2)沙蒿盖度通过沙土对结皮盖度的间接影响最大(0.422),土壤含水率通过沙土对结皮盖度的间接影响(-0.418)次之,且影响为负。(3)沙土通过其他环境因子对结皮盖度的间接影响为0.067(小于直接影响0.720),说明沙土是影响结皮盖度的最直接因子;沙蒿盖度通过其他环境因子对结皮盖度的间接影响为0.403,直接影响为0.235,且其仅通过沙土对结皮盖度的间接影响就已经达到0.422,即沙蒿盖度对结皮盖度的间接影响大于直接影响且以通过沙土的间接影响最为主要;土壤含水率对结皮盖度的直接影响为0.526,通过其他环境因子的间接影响为-0.612(小于0),导致土壤含水率与结皮盖度呈负相关关系(相关系数为-0.086),故实际上低估了土壤含水率对结皮盖度的直接影响。
2.2.3 各环境因子对结皮盖度的决定系数和决策系数 分析各环境因子对结皮盖度的决定系数和决策系数(表3)可知:(1)6个环境因子的直接决定系数的大小依次为沙土>土壤含水率>海拔>坡向>沉积物运移指数>沙蒿盖度,即沙土及土壤含水率对结皮盖度的直接决定作用最大;(2)共同决定系数中,沙土和土壤含水率的共同决定系数最大(-0.44),其次是沙土和沙蒿盖度(0.198)以及沙土和海拔(0.176)。(3)从决策系数来看,6个环境因子决策系数绝对值的大小依次为:沙土>土壤含水率>海拔>沙蒿盖度>坡向>沉积物运移指数,即沙土、土壤含水率、海拔和沙蒿盖度是主要的决策变量,其中除土壤含水率的决策系数为负值之外,其余均为正值。

表2 各环境因子对结皮盖度的通径系数
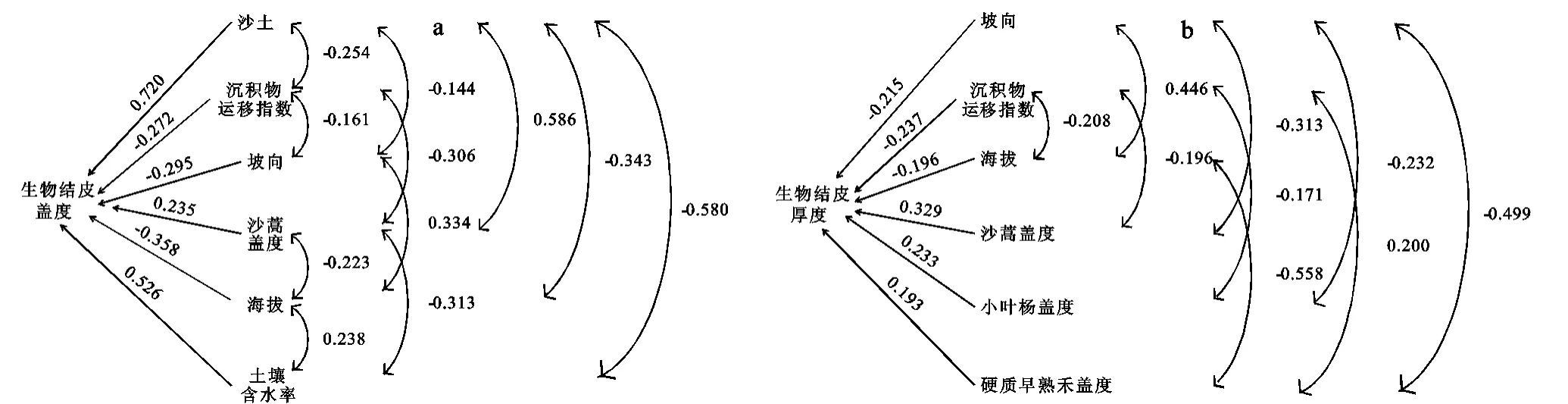
图1 生物结皮盖度、厚度与环境因子间的通径关系

表3 各环境因子对结皮盖度的决定系数和决策系数
因此,综上所述,沙土、土壤含水率、海拔及沙蒿盖度为影响结皮盖度的主要因素,但各自对生物结皮盖度的主要作用路径不同;沙土的直接作用(0.72),沙蒿盖度通过沙土的间接作用(0.422)是其主要的作用路径,两者都对结皮盖度有促进作用;土壤含水率的直接正作用(0.526)和间接负作用(-0.612)相当,因而总体贡献并不明显;海拔的直接和间接作用均为负(-0.358和-0.244),抑制作用明显。
2.3 生物结皮厚度与环境因子间的关系
结皮厚度是描述和衡量生物结皮发育状况的另外一个重要指标。为研究结皮厚度与环境因子之间的关系,根据逐步回归结果,选取坡向、沉积物运移指数、海拔、沙蒿盖度、小叶杨盖度及硬质早熟禾盖度作为影响因变量生物结皮厚度的自变量,进行相关及通径分析。
2.3.1 结皮厚度及各环境因子的相关分析 对结皮厚度及坡向、沉积物运移指数、海拔、沙蒿盖度、小叶杨盖度及硬质早熟禾盖度采用SPSS 12.0软件进行相关和回归分析,得出上述6个环境因子与结皮厚度的相关及回归系数(表4)。
由表4可以看出,6个环境因子与结皮厚度相关系数的绝对值大小依次为:坡向>沙蒿盖度>海拔>小叶杨盖度>硬质早熟禾盖度>沉淀物运移指数,且沙蒿盖度、小叶杨盖度及硬质早熟禾盖度与结皮厚度呈正相关关系,坡向、海拔及沉积物运移指数与结皮厚度均呈负相关关系,各环境因子均与结皮厚度的相关性达到极显著水平(p<0.01)。此外,各环境因子之间也存在着不同程度的显著相关性,即共线性;6个环境因子与结皮厚度之间标准化回归方程达到极显著水平(p<0.01),R2=0.552,说明结皮厚度关于上述6个环境因子的通径分析仍然是有必要的。
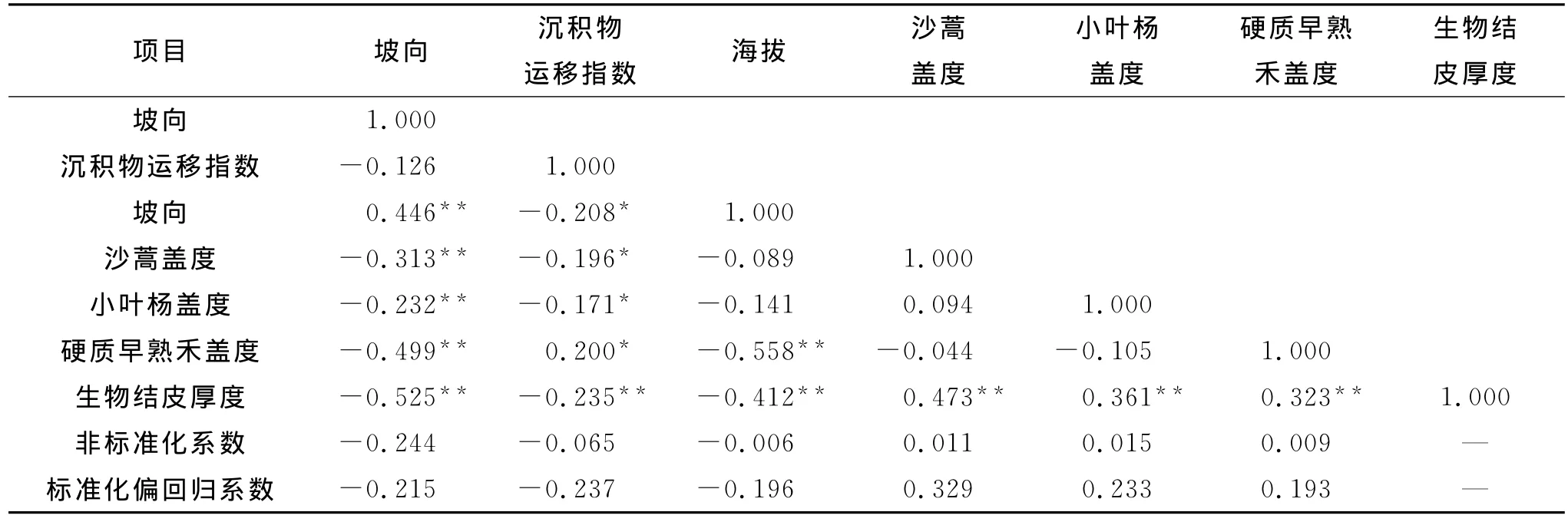
表4 环境因子与结皮厚度的相关与回归分析
2.3.2 生物结皮厚度及各环境因子的通径分析 表5和图1b展示了不同环境因子对结皮厚度的直接和间接影响。从表5可以看出:(1)直接通径系数的大小依次为沉积物运移指数(-0.237)<坡向(-0.215)<海拔(-0.196)<0<硬质早熟禾盖度(0.193)<小叶杨盖度(0.233)<沙蒿盖度(0.329),即沙蒿盖度对结皮厚度的直接影响最大,沙蒿盖度越大,结皮厚度越大。(2)硬质早熟禾盖度通过海拔对结皮厚度的间接影响(0.109)最大,海拔通过硬质早熟禾盖度对结皮厚度的间接负影响(-0.108)最大。(3)坡向通过其他环境因子对结皮厚度的间接负影响(-0.311)最大,其中以通过沙蒿盖度的间接影响(-0.103)最为突出,其导致坡向与结皮厚度之间呈负相关关系(相关系数为-0.526)。

表5 各环境因子对结皮厚度的通径系数
2.3.3 各环境因子对结皮厚度的决定系数和决策系数 分析各个环境因子对结皮厚度的决定系数和决策系数(表6)可以看出:(1)6个环境因子直接决定系数的大小依次为沙蒿盖度>沉积物运移指数>小叶杨盖度>坡向>海拔>硬质早熟禾盖度,即沙蒿盖度对结皮厚度的直接决定作用最大(0.108),沉积物运移指数和小叶杨盖度次之,分别为0.056,0.054;(2)共同决定系数中,沙蒿盖度和坡向的共同决定系数最大(0.044),硬质早熟禾盖度与坡向和海拔的共同决定作用次之(均为0.042);(3)从决策系数来看,6个环境因子决策系数的大小依次为:沙蒿盖度>坡向>海拔>小叶杨盖度>硬质早熟禾盖度>沉积物运移指数,且各因子的决策系数均为正,对结皮厚度均起促进作用,说明沙蒿盖度、坡向及海拔是比较重要的环境决策变量,沉积物运移指数的作用最小;(4)6个环境因子对结皮厚度的总决定作用达55.2%(表6),说明这些环境因子是影响结皮厚度的主要因素,对结皮盖度的影响占相对重要的地位,因此在培育生物结皮中应予注意。
综上所述,沙蒿盖度、坡向及海拔是影响生物结皮空间分布和发育的关键因素,但三者对生物结皮盖度的主要作用路径不同,沙蒿盖度的直接作用是主要的作用路径,而坡向通过沙蒿盖度、海拔通过硬质早熟禾盖度的间接作用是主要作用路径。

表6 各环境因子对结皮厚度的决定系数和决策系数
3 结论与讨论
本研究的结果表明,生物结皮在黄土区和沙土区的空间分布具有明显差异。在沙土区呈连续分布,多发源于沙土和黄土交界处的低洼带,平均盖度达30%以上;而在黄土区呈离散的零星分布,盖度大多在20%以下,这与前人的结论一致[8],其原因可能是,由于低洼带具有较好的水分条件,其有利于沙土区生物结皮的连片发育;而黄土区的植被条件较好,植被及其枯落物的覆盖使生物结皮的发育空间受到限制[20]。
各环境因子对结皮盖度都有不同程度的影响。各环境因子的决策系数除土壤含水率为负值之外,其余均为正值,而造成其决策系数为负的原因可能是:土壤含水率高的地方被更多的维管植物占据,若维管植物大量繁衍,将大大降低生物结皮的生存空间,使得结皮盖度有所减少[12]。故土壤质地(沙土)、土壤含水率、海拔及沙蒿盖度可作为影响结皮盖度的主要因子,四者是影响生物结皮空间分布和发育的关键因素,这与前人的结论一致[13]。土壤质地(沙土)是影响结皮盖度的直接因子,对其直接决定作用最大,沙蒿盖度是影响结皮盖度的间接因子,对其间接决定作用最大,两者均对结皮盖度起促进作用,土壤含水率的直接正作用和间接负作用相当,因而总体贡献并不明显;海拔的直接和间接作用均为负。主要因子对生物结皮盖度的作用路径不同,土壤质地(沙土)的直接作用、沙蒿盖度通过土壤质地(沙土)的间接作用是其主要的作用路径。
各环境因子对结皮厚度的影响程度不同。沙蒿盖度、坡向及海拔可作为影响结皮厚度的主要因子。沙蒿盖度是直接影响因子,对结皮厚度的直接决定作用最大,且沙蒿盖度越大,结皮厚度越大,原因可能是沙蒿独特的灌丛群落结构为生物结皮的定居和增殖提供了相对稳定的环境[20],有利于生物结皮的发育。坡向及海拔是影响结皮厚度的间接因子,其间接决定作用大于直接决定作用,且生物结皮厚度阴坡大于阳坡,这与前人的结论一致[9];海拔对结皮厚度起促进作用,而前人就海拔对结皮厚度影响的研究还鲜有报道。但三者对生物结皮厚度的主要作用路径不同,沙蒿盖度的直接作用、坡向通过沙蒿盖度及海拔通过硬质早熟禾盖度的间接作用是其主要的作用路径。
此外本研究中,坡向、沉积物运移指数、海拔、沙蒿盖度、小叶杨盖度及硬质早熟禾盖度对生物结皮厚度的总的决定作用仅为55.2%,其可能是因为结皮厚度本身的变化就不大,影响生物结皮厚度的因素较为复杂,且主要是发育年限[20],而本研究区域内均为撂荒30a以上的样地,故并没有考虑这个因素。除此之外,本研究仅从地形因子(坡向、沉淀物运移指数及海拔)和植被因子(沙蒿盖度、小叶杨盖度及硬质早熟禾盖度)两方面对结皮厚度进行研究,没有更多的考虑土壤因子。本研究仅从环境因子这一较为宏观的方面出发,分析其对生物结皮的影响,以获得其主要的影响因子,而对影响生物结皮的微观因子及其影响机理尚待深入研究。
[1] Greene R S B,Eldridge D J.Microbiotic soil crusts:A view of their roles in soil and ecological processes in the rangelands of Australia[J].Australian Journal of Soil Research,1994,32(3):389-415.
[2] 李聪会,朱首军,陈云明,等.黄土丘陵区生物结皮对土壤抗蚀性的影响[J].水土保持研究,2013,20(3):6-10.
[3] 李新荣,张元明,赵允格.生物土壤结皮研究:进展、前沿与展望[J].地球科学进展,2009,24(1):11-24.
[4] 熊好琴,段金跃,王妍,等.毛乌素沙地生物结皮对水分入渗和再分配的影响[J].水土保持研究,2011,18(4):82-87.
[5] Fernando T M,Cristina E,Bowker M A.Biological crusts as a model system for examining the biodiversity ecosystem function relationship in soils[J].Soil Biol.Biochem.,2009,42:1-13.
[6] 吴明山,胥辉.通径分析在林业多元线性模型共线性分析中的应用研究[J].西部林业科学,2008,37(3):83-86.
[7] 赵益新,陈巨东.通径分析模型及其在生态因子决定程度研究中的应用[J].四川师范大学学报:自然科学版,2007,30(1):120-123.
[8] 卜崇峰,张朋,叶菁,等.陕北水蚀风蚀交错区小流域苔藓结皮的空间特征及其影响因子[J].自然资源学报,2014,29(3):490-499.
[9] 焦雯珺,朱清科,张宇清,等.陕北黄土区退耕还林地生物结皮分布及其影响因子研究[J].北京林业大学学报,2007,29(1):102-107.
[10] 桑广书.黄土高原历史时期植被变化[J].干旱区资源与环境,2005,19(4):54-58.
[11] 李国英.统一思想全面推进黄河流域水土保持工作[J].中国水土保持,2003(12):9-10.
[12] 卢晓杰,张克斌,李瑞.北方农牧交错带生物结皮的主要影响因子探讨[J].水土保持研究,2007,14(6):1-4.
[13] 吴玉环,高谦,于兴华.生物土壤结皮的分布影响因子及其监测[J].生态学杂志,2003,22(3):38-42.
[14] 吕建亮,廖超英,孙长忠,等.黄土地表藻类结皮分布影响因素研究[J].西北林学院学报,2010,25(1):11-14.
[15] 成向荣,黄明斌,邵明安.神木水蚀风蚀交错带主要人工植物细根垂直分布研究[J].西北植物学报,2007,27(2):321-327.
[16] 李勉,李占斌,刘普灵,等.黄土高原水蚀风蚀交错带土壤侵蚀坡向分异特征[J].水土保持学报,2004,18(1):63-65.
[17] 贾恒义,雍绍萍,王富乾.神木地区的土壤资源[C]∥中国科学院西北水土保持研究所集刊.西安:陕西科学技术出社,1993.
[18] 张琪,丛鹏,彭励.通径分析在Excel和SPSS中的实现[J].农业网络信息,2007(3):109-110.
[19] 袁志发,周静芋,郭满才,等.决策系数—通径分析中的决策指标[J].西北农林科技大学学报:自然科学版,2001,29(5):131-133.
[20] 孟杰.黄土高原水蚀交错区生物结皮的时空发育特征研究[D].陕西杨凌:西北农林科技大学,2011.
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04 13:45:40
草业学报(2021年6期)2021-06-28 11:01:36
读者·校园版(2020年20期)2020-10-13 05:32:58
中国农村科技(2020年8期)2020-09-05 08:41:40
水土保持通报(2020年6期)2020-03-10 01:46:52
文萃报·周五版(2019年12期)2019-09-10 07:22:44
散文选刊·下半月(2018年2期)2018-02-23 21:11:50
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10 00:09:15
环境昆虫学报(2016年5期)2016-11-18 00:43:21
食品工程(2016年1期)2016-11-14 03:23:18
