
基于TRIPLEX模型的浙江省主要森林类型生物量模拟及敏感性分析
2015-01-03 01:22高智慧吴初平黄玉洁焦洁洁
浙江林业科技 2015年6期
张 骏,葛 滢,高智慧,吴初平,黄玉洁,焦洁洁,江 波,常 杰*
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江省林业技术推广总站,浙江 杭州 310020;3. 浙江大学 生命科学学院,浙江 杭州 310058)
基于TRIPLEX模型的浙江省主要森林类型生物量模拟及敏感性分析
张 骏1,2,葛 滢3,高智慧2,吴初平1,黄玉洁1,焦洁洁1,江 波1,常 杰3*
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江省林业技术推广总站,浙江 杭州 310020;3. 浙江大学 生命科学学院,浙江 杭州 310058)
TRIPLEX模型是一种新兴混合模型,提供了一种整合目前模型并能有效解决所面临问题的观念和方法。本文以浙江省21个县4种主要林型(常绿阔叶林、针阔混交林、马尾松林和杉木林)、林龄5 ~ 50 a的147个样地森林生长和产量调查实测作为模型模拟和检验的数据,用TRIPLEX模型模拟和检验了样地的林分密度、树高、胸径、凋落物库、地上和总生物量,结果表明:4种主要林型的林地生长和产量模拟值和野外实测值的相关系数极高(p < 0.001),两者之间的偏差也较小;各个林型的地上及总生物量的模拟值和实测值相关性均极高(p < 0.001),决定系数r2均以马尾松林的最高(分别为0.95和0.94),以杉木林的最低;除杉木林外,常绿阔叶林、针阔混交林和马尾松林的地上及总生物量对于温度增长均是负相关,4种主要林型的地上及总生物量对于相对湿度增长均是正相关,对于降水量变化不相关。TRIPLEX模型最小化了输入参数,对于参数要求低,但未降低预测能力,而且能够在浙江省复杂气候和土壤条件下预测地上和总生物量。
TRIPLEX;生物量;生产力;模型检验;马尾松林;杉木林;阔叶林
森林是主要的陆地碳库,全球陆地的大部分碳储存在森林生态系统中,森林植被及土壤共储存了 1 146 Pg C[1],占全球陆地总碳库(2 477 Pg C)的46%[2]。作为一个动态的碳库,森林储存碳的能力不仅取决于其面积,还取决其质量,即单位面积的森林碳储量。由于施肥、火灾、病虫害及不合理的采伐方式的影响,全球森林都存在不同程度的退化[3~4]。森林生态系统的碳循环对现在及未来的气候变化和全球碳平衡都具有重要影响,反之气候变化也对森林固碳作用产生影响[5]。
模拟森林生长和碳动态变化的基本方法包括经验、机理和混合三种森林模型,都有各自优缺点[6~12]。经验模型(森林生长和产量模型)来源于大量的野外数据,通过以样地参数、年龄、密度和基面积等为变量的回归模型来描述生长速率。经验模型的主要优势是利用特定的数学函数或曲线描绘测量数据和生长变量之间的相关,仅需要简单的输入,易于构建。它们也易和各种管理分析和森林管理相结合,有效地为森林管理计划提供定量信息。然而,经验模型不能分析如气候或环境变化下森林生长的响应[7,13]。与经验模型不同,机理模型一般描述关键生态过程或模拟依赖例如光合、呼吸、分解和营养循环交互过程的生长,提供了检验和产生选择假设的途径,有助于精确描绘在给定环境变化下这些过程如何相互作用[11~12, 14]。因此,机理模型的主要优势在于包含了生理生态原理和预测长期变化环境下的响应。过去的十年,机理模型发展取得了许多进步,它能够结合能量、碳、水和营养循环。但是在森林管理中,机理模型很少被应用[10, 14~16]。
混合模型是一种能够弥补经验模型和机理模型各自缺点的模型[10~12, 17]。TRIPLEX模型作为一种新兴混合模型,模拟森林生态系统中关键的动力学过程,包括总初级生产力(GPP)、森林生长状况、土壤水平衡以及碳和氮的分配,它提供了一种整合目前模型并能有效解决所面临问题的观念和方法。目前为止,TRIPLEX模型已经成功运用于加拿大安大略北方针叶林中的12个加拿大短叶松永久样地[18],中国浙江省常绿阔叶林、针阔混交林和马尾松林[19]和湖南省杉木林和马尾松林[20]样地。
根据浙江省2013年底森林资源年度监测,全省森林面积604.78×104hm2中共有松木林面积81.16×104hm2,杉木林面积82.09×104hm2,阔叶林面积161.86×104hm2和针阔混交林面积68.25×104hm2[21],其中杉木林面积比例达18.44%。为此,本文在浙江省常见3种林型[19]基础上增加了杉木林样地,目标是:1)通过建立4种主要林型[22]的TRIPLEX模型参数体系,评价模型应用的有效性和实用性;2)运用混合模型TRIPLEX检验了 4种主要林型的地上和总生物量,并进行了敏感性分析,为进一步应用TRIPIEX模型模拟和预测浙江省森林生物量及土壤碳库对气候变化的响应奠定基础。
1 研究方法
1.1 TRIPLEX模型简介
TRIPLEX模型类似3-PG模型[23],用于预测森林生长情况以及碳氮动力学[24],简化了需要输入的参数个数,考虑了森林生态系统中树木生长关键过程及其与碳、氮、水循环之间的重要相互作用。另外,TRIPLEX模型模拟了立地的平均特性而非单株树木的立地条件。它的设计目标是:建立对不同地理位置、土壤和气候条件下的同、异龄的针、阔叶树种的参数化模型。如图1所示,TRIPLEX模型包括4个主要的子模型:
(1)TREEDYN3.0模型[25]中的光合辐射(PAR)子模型,估算PAR、总初生产力(GPP)和地上、地下生物量;
(2)CENTURY4.0模型中的土壤碳和氮分解子模型[26],模拟土壤和凋落物中碳、氮的动态变化;
(3)3-PG模型中的森林生长和产量子模型[23],计算树木生长和生产量的动态变化,包括树高、胸径、基面积和材积;
(4)土壤水平衡子模型,模拟水动态变化。
各模块利用目标导向程序(C++)方式以类列出。模型以月单位为时间步长,以每月气象数据驱动。TRIPLEX模型的关键部分和模拟过程见图1。

图1 TRIPLEX模型流程Figure 1 The structural model of forest growth and carbon simulation from TRIPLEX
1.2 模型初始化
TRIPLEX模型的模拟需要初始值,这些值由描述森林立地和土壤的变量组成。与森林生长和产量的初始条件有关的变量有3个,即树木密度、树高及胸径。用于描述土壤状态的初始条件包括有机碳密度和水分含量,这些数据均可从野外测定结果得到[27]。
TRIPLEX模型要求输入的参数最小化,并且参数化后代表本研究中浙江省4种主要林型。用于TRIPLEX模型校准的参数大致分为3种类型:(1)必须经过已有模型设置的未知参数;(2)从立地数据库中估计出来的已知参数;(3)从已发表的文献或研究中获得的已知参数。模型校准的主要目的在于获得(1)中这些未知参数。
为了有效检验TRIPLEX模型,继续保留大多数来自先前论文[24, 28~30]的非样地参数(表 1),包括PAR、气孔导度、初始氮水平、叶中木质素含量、木质素氮比例和土壤含水率等参数。

表1 TRIPLEX模型在浙江省模拟中应用的参数Table 1 Parameters used in TRIPLEX for simulating forest ecosystems in Zhejiang
1.3 模型运行
与模拟初始条件相关的3个关键变量是树木密度、树高和胸径。一些新的初始参数列在表2,例如碳密度、比叶面积(SLA)、死亡率、叶比例、干比例、粗细根的比例根据模型缺省值进行调整,从而更好的代表浙江省森林生态系统的研究。

表2 TRIPLEX模型在浙江省不同林型的参数Table 2 Parameters used in TRIPLEX to simulate different forest types in Zhejiang
TRIPLEX模型模拟了树木的整个生长期,每个立地从起始林龄一直模拟到野外调查那年的林龄。所有的模拟在不同的气候条件下以月时间步长计算。模型计算的树木生物量各组分(包括树叶、干和根)月增长,通过立地产量变量(密度、胸径、树高)推算,最终结果按每年总计。
TRIPLEX模型在运行模拟之前,每个林型各自随机选择样地进行校正和验证。然后分别对树高、胸径、凋落物库、地上和总生物量进行模拟。其他过程参考Zhou等[28]中模型的执行。
所有初始立地生物量测量均是起始年的1月值。树木密度初始化根据树种和分布,常绿阔叶林、针阔混交林、马尾松林和杉木林分别假定为4 950,4 500、4 183和4 267株/hm2。随后,立地密度根据起始密度结合因竞争引起的自疏死亡率变化。
1.4 模型检验
为了检验TRIPLEX模型更大范围的有效性和实用性,本研究继续沿用了上面提到过的研究工作[24]所采用的标准。除去初始化的样地,将分布于浙江省21县中四种主要林型的147个标准样地进行模型检验,评价模型的有效性,样地的详细介绍和森林生长和产量调查实测过程见张骏等[22]的相关报道。
1.5 模型敏感性分析
敏感性分析是新建立模型的重要一步,它系统检验模型行为,模拟系统运行机制[24]。本研究中,敏感性分析通过对森林生长和碳动力学最重要输出变量——生物量的增长百分比决定,包括对月均温、月降水量和月均湿度与先前相比10%内的增长或降低,然后运行模型模拟结果对比分析。
2 结果
2.1 模型有效性
通过比较浙江省森林树高和胸径的模拟值与野外实测值,检验模型的有效性。TRIPLEX模型的验证结果见表3。浙江省森林生态系统的模拟值和实测值的决定系数(r2)均很高,密度的为0.92,胸径的为0.77,树高的为0.80,地上生物量的为0.92,总生物量的为0.91,只有凋落物库的较低为0.53,但也极显著相关(p < 0.001)。模拟值和实测值之间的平均预测误差(e)除了密度的为53.56外,其他变量均很小,胸径的为0.32,树高的为-0.59,凋落物库的为1.15,地上生物量的为0.47,总生物量的为-0.90。模拟值和实测值之间的偏差(预测误差除以实测值)也较小:密度的为2.7%,胸径的为3.3%,树高的为-8.6%,地上生物量的为1.0%,总生物量的为-1.5%,只有凋落物库的偏差较大,达16.8%。
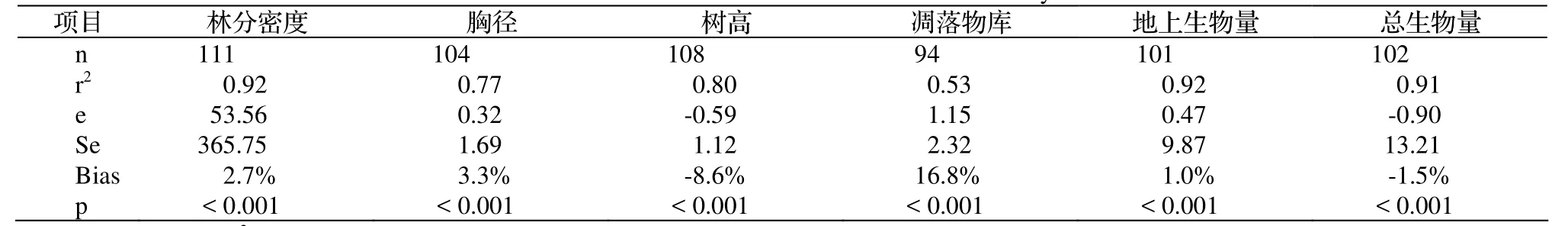
表3 TRIPLEX模型对浙江省森林生态系统密度、胸径、树高、凋落物库、地上生物量和总生物量模拟值和实测值比较的误差Table 3 Simulation errors of TRIPLEX applied to forest ecosystems in Zhejiang, comparing stand density, DBH, height, litter pool, aboveground and total biomass between simulated values and inventory ones
运用111组浙江省森林生态系统的生长和产量的实测数据,分别比较了地上和总生物量,模拟值和实测值有很高的相关性(图2)。
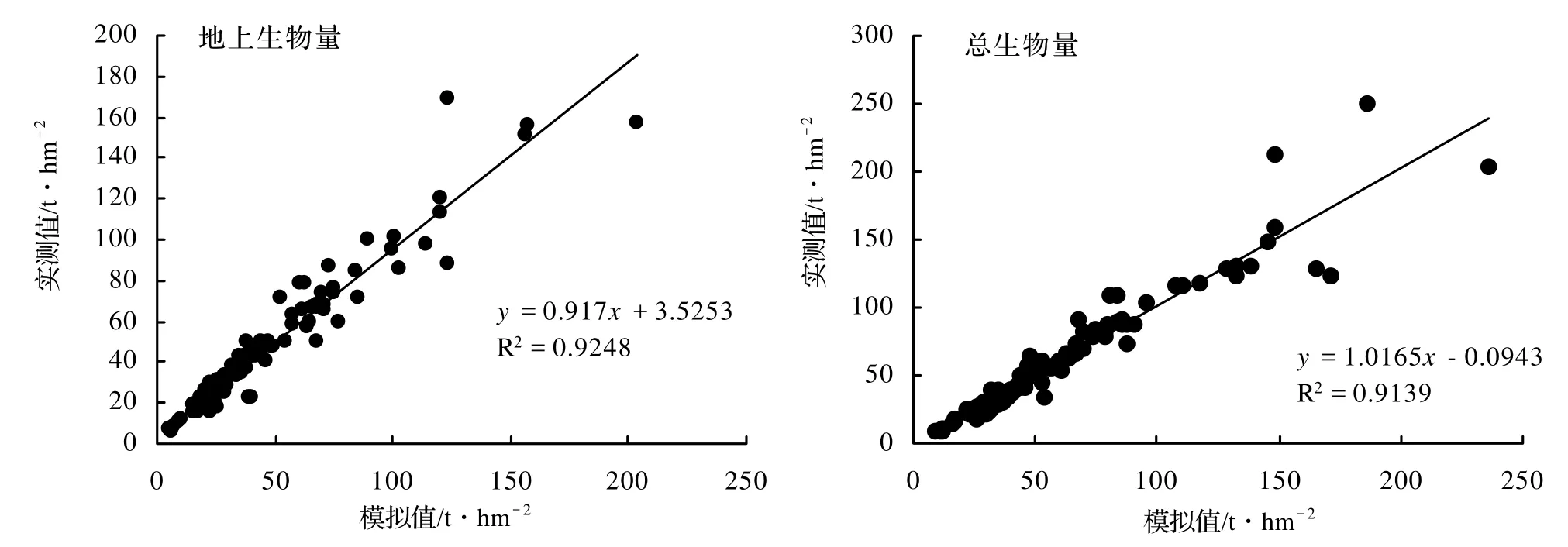
图2 浙江省森林地上和总生物量的模拟值和实测值比较Figure 2 Comparison on simulated and surveyed data of aboveground and total biomass in Zhejiang
2.2 4种主要林型的生物量模拟
表4比较了4种主要林型的地上和总生物量的模拟值和实测值的统计分析结果。各林型的模拟和实测值相关性极显著(p < 0.001):地上生物量模拟和实测值之间的相关系数是马尾松林的最高为0.95,杉木林的最低为0.63;总生物量模拟和实测值之间的相关系数也是马尾松林的最高为0.94,杉木林的最低为0.59。总生物量的预测误差大约-0.57 到-10.67 t/hm2,转换成偏差为-19.1%到0.8%。
2.3 模型生物量敏感性分析
除杉木林外,常绿阔叶林、针阔混交林和马尾松林的地上和总生物量对于温度的增长均是负相关;4种主要林型的地上和总生物量对于相对湿度的增长均是正相关;4种主要林型的地上和总生物量对于降水量的变化不相关(0.00%)。

表4 TRIPLEX模型对浙江省森林生态系统地上生物量和总生物量模拟值分别和33个常绿阔叶林(EF)、29个针阔混交林(MF)、32个马尾松林(PF)和32杉木林(CF)测量值比较的误差Table 4 Simulation err ors of TRIPLEX applied to four forest types in Zhejiang, comparing aboveground and total biomass between simulated values and inventory data from 33sample plots of evergreen broad-leaved forest, 29 of coniferous and broad-leaved mix ed forest, 32 of Pinus massoniana forest and 32 of Cunninghamia lanceolata forest

表 省4种 型代表 候变化 总生物 敏感性Table 5 Prediction sensitivitypes in Zhejiang 5 浙江 主要林 性样地气 对地上和 量的预测of aboveground and total biomass to changes in climatic of four main forest ty
3 结论与讨论
目前,大多数中国森林的生物量和NPP主要通过森林清查或一些模型获得[35~36],其中对浙江省森林生长的模拟包括CASA[37]和CEVSA[38~39]等模型的应用,我们将这些模型的预测值和清查结果进行了比较(表6)。
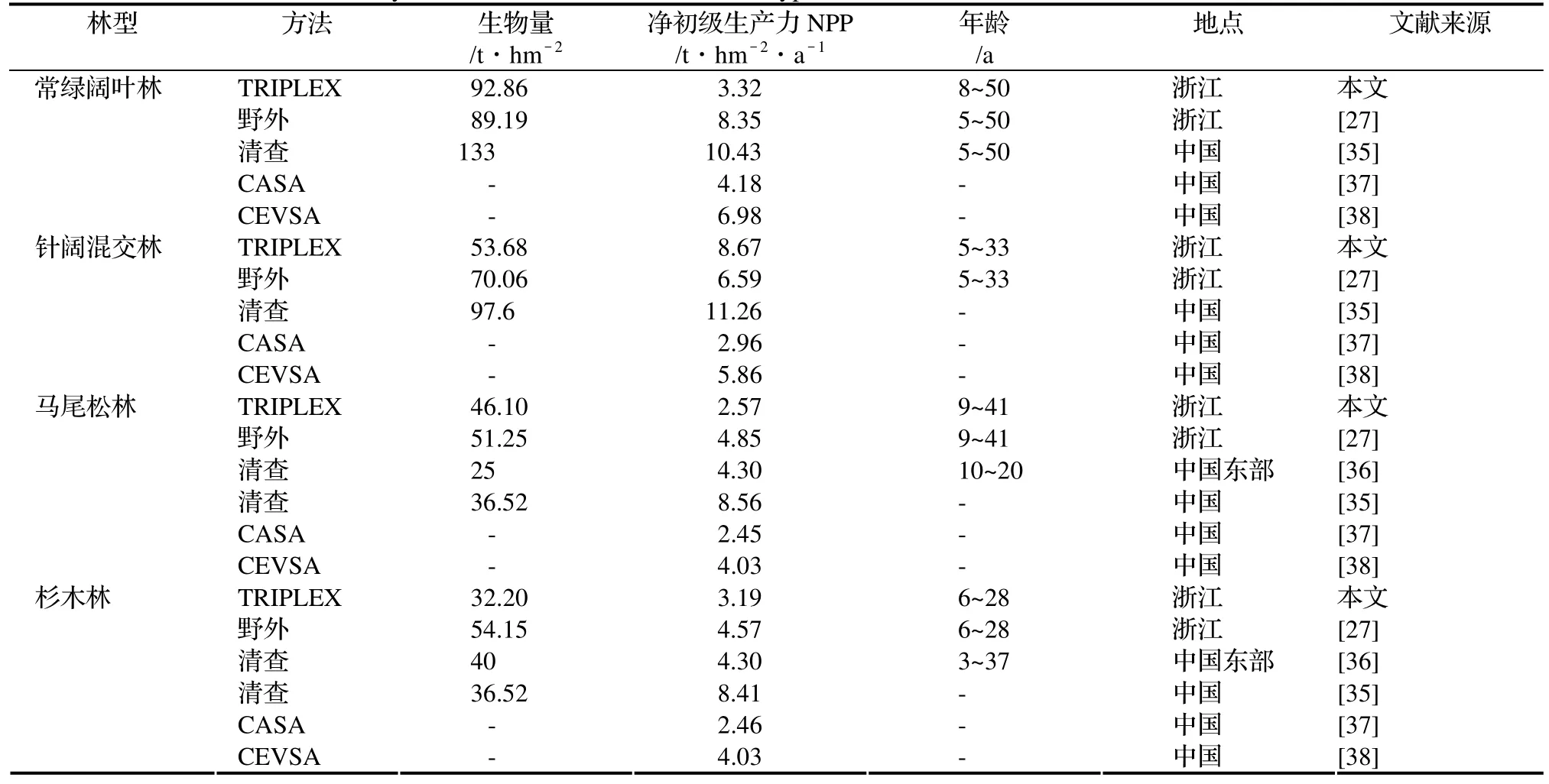
表6 4种林型的生物量和NPP实测和不同模型模拟结果比较Table 6 Surveyed data of biomass and NPP in four types of forest and simulations with different methods
与类似区域不 同方法比 较发现, T RIPLEX模型对所 有林 型的生物量 预测均 最接近野 外实测值。T RIPLEX模型模拟生物量结果:常绿阔叶林(92.86 t/hm2)和马尾松林(46.10 t/hm2)生物量在野外实测值之间,针阔混交林(53.68 t/hm2)和杉木林(32.20 t/hm2)生物量比其他实测值低。除去常绿阔叶林的NPP低于其他清查和模拟结果外,TRIPLEX模型模拟的其他 3种林型的NPP比CASA和CEVSA模拟的更接近于清查实测值:针阔混交林在Zhang 等[27]和方精云等[35]结果之间,但高于Cao[38]的预测;马尾松林和杉木林均在Cao[38],Fang等[37]和Zhang[27]结果之间,但低于方精云等[28]的结果。
TRIPLEX模型最小化了输入参数,对于参数要求低,但未降低预测能力(表6),而且能够在浙江省复杂气候和土壤条件下预测地上和总生物量。
浙江省森林生态系统的模拟和实测值的决定系数均很高,只有凋落物库的较低为 0.53,凋落物库的偏差也最大(表3),可能因为凋落物等野外数据获得困难和TRIPLEX模型时间尺度上的误差容易积累,所以和土壤碳库直接相关的凋落物预测有效性差些。虽然TRIPLEX没有考虑人类干扰、火灾以及土地利用变化,但以上结果说明TRIPLEX模型能够在浙江省森林主要林型适用。
敏感性分析对模型关键参数的有效估算和最佳模拟精度十分重要。通过敏感性分析发现,除杉木林外,其他3种林型的地上和总生物量与温度负相关,与湿度正相关,与降雨量不相关。杉木林的模拟和实测值的相关系数也是最低,主要是因为杉木林早期作为人工林培育,受干扰强度远远大于另外3种林型,故有效性和敏感性与其他3种林型不一致。随着农业、森林资源砍伐和城市化等人类干扰的加强,陆地生态系统变化加大而导致未来复杂性增加[39],将来应考虑在TREPLEX中加入生态系统干扰对于森林生长和碳动态变化影响的额外模块。
另外由于时间、精力原因,本次研究中采用的一些样地参数例如GPP到NPP转换系数、比叶面积、死亡率等仅仅参考模型缺省值进行微调,在TREPLEX模型以后的本土化中,这些样地参数需要进一步考虑根据本地区林型对各种人为干扰响应的实测数据和文献进行参数化。
[1] Dixon R K, Brown S,Houghton R A, et al. Carbon pools and fluxes of global forest ecosystems [J]. Science, 1994(263):185-190.
[2] IPCC. The Science of Climate Change [M]. Cambridge: Cambridge University Press, 2000.
[3] Smith R. Global Forest Resources Assessment 2000. Main report [M]. Rome: Food and Agriculture Organization (FAO), 2002
[4] Houghton R A. Terrestrial source and sinks of carbon inferred from terrestrial data [J]. Tellus B, 1996(48):420-432.
[5] Friedlingstein P, Dufresne J L, Cox P M, et al. How positive is the feedback between climate change and the carbon cycle [J]. Tellus, 2003 (55B):692-700.
[6 ] Landsberg J J. Physiological Ecology of Forest Production [M]. Sydney: Academic Press, 1986.
[7] Kimmins J P. Modeling the sustainability of forest production and yield for a changing and uncertain future[J]. Forestry Chronicle, 1990, 66(3):271-280.
[8] Mohren G M L, Burkhart H E, Jansen J J. Contrasts between biologically-based process models and management-oriented growth and yield models [J]. For Ecol Manage, 1994(69):1-5.
[9] Landsberg J J, Coops N C. Modeling forest productivity across large areas and long periods [J]. Nat Resoure Model, 1999(12):383-411.
[10] Mäkelä A, Landsberg J, E k A, et al. Process-based models for forest ecosystem management: current state of the art and challenges for practical implementation [J]. Tree Physiol, 2000(20):289-298.
[11] Peng C H. Understanding the role of forest simulation models in sustainable forest management [J]. Environ Impact Assess Rev, 2000(20):481 -501.
[12] Peng C H. Growth and yield models for uneven-aged stands: past, present and future [J]. For Ecol Manage, 2000(132):259-279.
[13] Shugart H H, Smith T M, Post W M. The application of individual-based simulation models for assessing the effects of global change [J]. Annu Rev Ecol Syst, 1992(23):15-38.
[14] Landsberg J J, Gower S T. Applications of Physiological Ecology to Forest Management[M]. San Diego: Academic Press, 1997.
[15] Korzukhin M D, Ter-Mikaelian M T, Wagner R G. Process versus empirical models: which approach for forest ecosystem management [J]. Can J For Res, 1996(26):879-887.
[16]Sands P J, Battaglia M, Mummery D. Application ofprocess-based models to forest management: experience with PROMOD, a simple plantation productivity model [J]. Tree Physiol, 2000(20):383-392.
[17] Johnsen K, Samuelson L, Teskey R, et al. Process models as tools in forestry research and management [J]. For Sci, 2001(47):2-8.
[18] Peng C H, Jiang H, Apps M J, et al. Effects on harvesting regimes on carbon and nitrogen dynamics of boreal forests in central Canada: a process model simulation [J]. Ecol Model, 2002(155):177-189.
[19] Zhang J, Chu Z Y, Ge Y, et al. TRIPLEX Model Testing and Application for Predicting Forest Growth and Biomass Production in the Subtropical Forest Zone of China’s Zhejiang Province [J]. Ecol Model, 2008(219):264-275.
[20] Zhao M F, Xiang W H, Deng X W, et al. Application of TRIPLEX model for predicting Cunninghamia lanceolata and Pinus massoniana forest stand production in Hunan Province, southern China[J]. Ecol Model, 2013(250):58-71.
[21] 浙江省森林资源状况及其生态功能价值(2013)[EB/OL]. http://zjrb.zjol.com.cn/html/2015-01/21/content_2845042.htm
[22] 张骏,袁位高,葛滢,等. 浙江省生态公益林碳储量和固碳现状及潜力[J]. 生态学报,2010,30(14):3 839-3 848.
[23] Landsberg J J, Waring R H. A generalized model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning [J]. Ecol Model, 1997(95):209-228.
[24] Peng C H, Liu J, Dang Q, et al. TRIPLEX: A generic hybrid model for predicting forest growth and carbon and nitrogen dynamics [J]. Ecol Model, 2002(153):109-130.
[25] Bossel, H. TREEDYN3 Forest Simulation Model [J]. Ecol Model, 1996,90(3):187-227.
[26] Parton W J, Scurlock J M, Ojima D S, et al. Observations and modelling of biomass and soil organic matter dynamics for the grassland biome worldwide [J]. Glob Biogeochem Cycle, 1993,7(4):785-809.
[27] Zhang J, Ge Y, Chang J, et al. Carbon storage by ecological service forests in Zhejiang Province, subtropical China [J]. For Ecol Manage, 2007 (245):64-75.
[2 8] Zhou X L, Peng CH, Dang Q L. Assessing the generality and accuracy of the TRIPLEX model using in situ data of boreal forests in central Canada [J]. Environ Model Software, 2004(19):35-46.
[29] Zhou X L, Peng C H, Dang Q L, et al. Predicting forest growth and yield in northeastern Ontario using the process-based carbon dynamic model of TRIPLEX 1.0 [J]. Can J For Res, 2005(35):2 268-2 280.
[30] Zhou X L, Peng C H, Dang Q L, et al. A simulation of temporal and spatial variations in carbon at landscape level: A case study for Lake Abitibi Model Forest in Ontario, Canada [J]. Mitigat Adapt Strateg Glob Change, 2007, 12(4):525-543.
[31] Kimball J S, Thornton P E, White M A, et al. Simulating forest productivity and surface–atmosphere carbon exchange in the BOREAS study region [J]. Tree Physiol, 1997(17):589-599.
[32] Ryan M G, Lavigne M B, Gower S T. Annual carbon cost of autotrophic respiration in boreal forest ecosystems in relation to species and climate [J]. J Geophys Res, 1997, 102: 28871-28883.
[33] Metherell A K, Harding L A, Cole C V, et al. CENTURY Soil Organic Matter Model Environment Technical Documentation Agroecosystem Version 4.0[J]. GPSR Technical Report No. 4, United States Department of Agriculture, Agricultural Research Service, Great Plains Systems Research Unit. http://www.nrel.colostate.edu/ projects/century/ [cited 10 May 2004], 1993.
[34] 方精云,刘国华,徐嵩龄. 我国森林植被的生物量和净生产量[J]. 生态学报,1996,16(5):497-508.
[35] Zhao M, Zhou G S. Estimation of biomass and net primary productivity of major planted forests in China based onforest inventory data[J]. For Ecol Manage, 2005(207):295-313.
[36 ] Fang J Y, Piao S L, Field C B, et al. Increasing net primary production in China from 1982 to 1999 [J]. Front Ecol Environ, 2003(1):293-297. [37] Cao M K, Woodward F I. Net primary and ecosystem production and carbon stocks of terrestrial ecosystem and their response to climatic change [J]. Glob Change Biol, 1998(4):185-198.
[38] Cao M K, Tao B, Li K R. Interannual variation in terrestrial ecosystem carbon Fluxes in China from 1981 to 1998 [J]. 植物学报(英文版), 2003,45(5):552-560.
[39] Chapin F S, Callaghan T V, Bergeron Y. Global change and the boreal forest: thresholds, shifting states or gradual change[J]. Ambio, 2004, 33 (6):361-365.
Simulation and Sensitivity for Biomass of Four Main Forest Types in Zhejiang Using TRIPLEX Model
ZHANG Jun1,2,GE Ying3,GAO Zhi-hui2,WU Chu-ping1,HUANG Yu-jie1,JIAO Jie-jie1,JIANG Bo1,CHANG Jie3
(1. Zhejiang Forestry Academy, Hangzhou 310023, China; 2 Zhejiang Forestry Extension Station, Hangzhou 310020, China; 3. College of Life Sciences, Zhejiang University, Hangzhou 310058, China)
Inventory data of growth and yield from 147 sample plots aged from 5 to 50 years of four common forest types in 21 counties of Zhejiang province was used to test the process-based model of TRIPLEX. The four main forest types are evergreen broad-leaved forest (EF), coniferous and broad-leaved mixed forest (MF), Pinus massoniana forest (PF) and Cunninghamia lanceolatae forest (CF). Simulation values by TRIPLEX model with stand density, diameter at breast height(DBH), height(H), litterfall pool, aboveground and total biomass were compared with measured ones. The results showed that significant correlation (p<0.001) between the simulated and measured values of forest stands for four main forest types in Zhejiang province was found and the coefficients of determination (r2) was 0.92 for density, 0.77 for DBH, 0.80 for H, 0.53 for litterfall pool, 0.92 foraboveground biomass and 0.91 for total biomass, with small errors of -0.32 for DBH, -0.59 for H, 1.15 for litterfall pool, -0.51 for the aboveground biomass and -2.64 for the total biomass, except for stand density (53.56), and low biases (2.7% for density, -3.3% for DBH, -8.6% for H, -1.0% for the aboveground biomass and -1.5% for the total biomass) except for litterfall pool (16.8%). The simulated and measured values had significant correlation (p<0.001) for the aboveground and total biomass for each forest type, PF had highest coefficient of determination within 0.95 and 0.94 respectively, and CF had lowest coefficient of determination. For the four main forest types, Aboveground and total biomass of tested forest types had negative correlation with temperature except CF, and positive relation with relative humidity, and no correlation with precipitation. Simulated values of biomass and NPP of four forest types in Zhejiang by TRIPLEX model were much closer to the surveyed ones than those by the CEVSA and CASA models. These results suggest that TRIPLEX model did not decrease the predictive accuracy at the low demand for parameters, which can simulate and predict biomass with complex conditions in climate and soil.
TRIPLEX; biomass; NPP; model test, Pinus massoniana forest, Cunninghamia lanceolatae, broad-leaved forest
S718.5
:A
1001-3776(2015)06-0001-08
2015-06-08;
2015-10-11
浙江省自然科学基金项目(LQ13C030001);浙江省森林生态科技创新团队(2011R50027);浙江省省级定位站管理维护及技术支撑项目
张骏(1981-),男,浙江龙游人,副研究员,博士,研究森林生态及林技推广;*通讯作者。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
环境保护与循环经济(2021年7期)2021-11-02
哈尔滨轴承(2020年1期)2020-11-03
农家科技中旬版(2020年10期)2020-07-12
农民致富之友(2020年8期)2020-05-11
农家科技中旬版(2020年2期)2020-03-18
绿色科技(2019年5期)2019-11-29
中国奶牛(2019年10期)2019-10-28
电子制作(2018年23期)2018-12-26
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
