
毛茛科雄蕊结构的分类学价值
2015-01-02 08:12:42程薪宇刘玫郭梦桥
草业学报 2015年3期
程薪宇,刘玫*,郭梦桥
(1.哈尔滨师范大学生命科学与技术学院,黑龙江省分子细胞遗传与遗传育种重点实验室,黑龙江 哈尔滨150025;2.黑龙江省科学院大庆分院,黑龙江 大庆163319)
毛茛科(Ranunculaceae)为真双子叶植物根部群[1],全世界约60属,2500种,主要分布于温带和寒温带[2-4],中国有38属,921种[3]。经典分类学研究多以植物外形特征为参考依据[5-7],而毛茛科亦是如此[8-11],如 Hutchinson[12]根据果实特征将毛茛科分为金莲花亚科(Helleboroideae)(具蓇葖果)和毛茛亚科(Ranunculoideae)(具瘦果)。但这一科后被分为4个亚科,芍药亚科(Paeonioideae)(雄蕊离心发育,有花盘,蓇葖果),金莲花亚科(雄蕊向心发育,无花盘,蓇葖,浆果或蒴果),唐松草亚科(Thalictroideae)(雄蕊向心发育,无花盘,蓇葖或蒴果),及毛茛亚科(雄蕊向心发育,无花盘,瘦果,稀浆果)[8-9]。Takhtajan[10]认为芍药亚科的一些特征(如雄蕊的发育方式及花的特征)与其他亚科区别较大,将其从毛茛科分出,另立为芍药科(Paeoniaceae)。
Decraene和Smets[13]与Endress[14]曾指出毛茛科植物雄蕊的特征具有较高的系统学价值。一些研究显示毛茛科不同类群雄蕊的排列方式不同,如驴蹄草属(Caltha)、黄连属(Coptis)及金莲花属(Trollius)的雄蕊螺旋状排列[15-16],耧斗菜属(Aquilegia)等属的雄蕊轮状排列[17-18]。此外雄蕊的发育方式也有区别,如驴蹄草属,金莲花属,侧金盏属(Adonis)及黄三七属(Souliea)雄蕊向心发育[15,19-20],耧斗菜属雄蕊离心发育[17,21]。Wang[22]注意到花丝的特征,指出铁线莲属(Clematis)对枝铁线莲组(sect.Brachiatae)的雄蕊花丝均为狭条形,被柔毛,花药通常长圆形或狭长圆形,无毛,药隔顶端通常不突出。然而,现有关于毛茛科雄蕊的研究,也仅限于对科内少数几个属雄蕊早期发育形态的观察[15-21]及成熟雄蕊外形的简要描述[11,22],未见有关花药及花丝结构的详细报道。因此,本文深入研究了毛茛科部分类群幼嫩及成熟雄蕊的外部形态及其内部结构,旨在揭示雄蕊外形及显微结构在毛茛科分类学研究中的价值,并为进一步研究毛茛科系统学提供形态学资料。
1 材料与方法
1.1 材料
研究材料为国产毛茛科18属,37种,5变种的幼嫩及成熟雄蕊。凭证标本存于哈尔滨师范大学植物标本馆(HANU)。材料名称,凭证标本及物种采集地信息见表1。
1.2 方法
雄蕊形态及其毛状体的观察:用Olympus SZX16观察20朵成熟花中雄蕊的形态,并测量其花药及花丝长度。将新鲜完整的雄蕊及撕去表皮的花丝分别置于载玻片上,各滴1~2滴50%甘油水溶液,盖上盖玻片。分别用Olympus SZX16及Olympus BX53观察雄蕊表面毛状体及花丝内薄壁组织的特征,所有观察结果用Olympus DP26成像系统照相。
雄蕊解剖结构观察:采取GMA(Glycol methacrylate,乙二醇甲基丙烯酸酯)半薄切片法观察毛茛科雄蕊内部结构,即根据Feder和 O’Brien[23]的方法将幼嫩及成熟的花用FAA(Formalin-acetic acid-alcohol mixture,福尔马林-醋酸-酒精混合液)固定24h以上,从不同的花上取幼嫩及成熟的雄蕊各3个,经50%乙醇,100%乙醇,100%异丙醇和100%正丁醇脱水处理各2次(每次4h)。随后,经GMA渗透3次(第1~2次各1d,第3次3d)后,将材料及GMA装入胶囊,置于60℃温箱24h进行聚合。用Leica Ultralcut R切片,厚度约1.5μm,同样用Feder和O’Brien[23]的方法对切片进行染色,切片干燥后用中性树胶封片。用Olympus BX53观察及O-lympus DP26照相。
2 结果与分析
本文深入研究了毛茛科一些类群(18属,37种,及5变种)的幼嫩及成熟雄蕊形态,揭示了不同物种雄蕊的形状,毛状体特征,花丝及花药的结构(图1,图2),详细特征见表2。
2.1 雄蕊外形及毛状体
毛茛科一朵花中成熟雄蕊的花丝长度差异较大,如朝鲜白头翁靠近花瓣的花丝为1.0mm,靠近雌蕊的为12.5mm(图1A),或近等长,如贝加尔唐松草的花丝长3.6~4.7mm(图1B)。不同属的花丝形状不尽相同,如耧斗菜属的呈丝状(图1D),铁线莲属为条形(图1E,图2C),唐松草属部分物种的花丝为棒状(图1H,图2D)。此外,
还有长三角形(即花丝基部条形,顶端丝状,见于乌头属和翠雀属,图1F)和短柱状(如水葫芦苗,图1G)花丝。除铁线莲属的花药长度变化较大外(约1.4~8.0mm,图1E),其他属变化较小。但花药长度在不同属中变化较大,如碱毛茛属水葫芦苗花药长约0.8mm(图1G),而金莲花属长瓣金莲花长达4.5mm。花药侧面观为∞形(仅见于乌头属,图1C)、椭圆形(如贝加尔唐松草及耧斗菜,图1H)、近圆形(如黑水银莲花及水葫芦苗,图1G)、条形(多见于铁线莲属和金莲花属,图1E)。多数物种雄蕊表面无毛(如水葫芦苗,尖萼耧斗菜),少数物种具单细胞毛状体,毛状体分为非腺毛(如齿叶铁线莲及翠雀,图1J),或乳头状腺毛(如宽苞翠雀花及翠雀,图1I)。毛状体着生于花药(如宽苞翠雀花),花丝(如齿叶铁线莲)或整个雄蕊上(如翠雀)。

表1 毛茛科物种及其凭证标本,采集地和采集时间Table 1 Taxa of Ranunculaceae,together with voucher specimen details and locality

续表1 Continued
2.2 花丝及花药结构
花丝结构:丝状,棒状及短柱状花丝横切面为圆形或近圆形(如三角叶驴蹄草,图2B,贝加尔唐松草,图2D);条形花丝较扁(如铁线莲属,图2C);长三角形(如乌头属,图1C)花丝基部较扁,顶端圆形或近圆形。花丝表皮细胞近方形(如宽叶蔓乌头,图2A)或圆形(如三角叶驴蹄草,图2B),外切向壁明显增厚(如宽叶蔓乌头,图2A)或轻微增厚(如三角叶驴蹄草,图2B)。表皮以内为多层薄壁细胞,长三角形花丝的薄壁组织无胞间隙,或胞间隙不发达(如宽叶蔓乌头,图2A);丝状、条状及短柱状花丝胞间隙不发达(如转子莲,图2C)或发达(如三角叶驴蹄草,图2B);棒状花丝胞间隙极发达(如唐松草属部分物种,图2D),且细胞排列成海绵状(如狭裂瓣蕊唐松草,图1K)。花丝中央具1条外韧维管束,木质部仅由少数导管组成(图2B)。
幼嫩花药结构:幼嫩花药具2对花粉囊,每对花粉囊由数层薄壁细胞组成的隔分开(图2E1~J)。花药横切面形状分为6种。(1,2)V形和Λ形:4个花粉囊大小相同,药隔一侧凹陷。V形的花粉囊朝向近轴面(即维管束木质部一侧)(仅乌头属,图2E1),Λ形花粉囊朝向远轴面(即维管束韧皮部一侧)(仅毛茛属,图2I1)。(3)∞形:药隔处凹陷(见于耧斗菜属和白头翁属等属,图2F1)。(4)椭圆形:药隔处无凹陷(如银莲花属和铁线莲属部分物种,图2G)。(5)方形:药隔及2花粉囊交界处凹陷(如碱毛茛属和唐松草属,图2J)。上述5种类型花药的花粉囊体积相近。(6)蝶形:花粉囊大小不同,2个小花粉囊朝向近轴面,2个大的靠近远轴面(如扁果草属,图2H)。药隔两侧的2花粉囊由数层薄壁细胞隔开(图2E1~J),这些细胞在V和Λ形花药中与花药的横切面形态相似(如乌头属及毛茛属,图2E1,I1),在∞形、椭圆形、方形花药横切面中呈直线形排列(如辣蓼铁线莲,图2G),在蝶形花药中排列成V形或直线形(如东北扁果草,图2H)。药隔均具1条维管束,维管束结构同花丝(图2E1,F1)。

图1 毛茛科雄蕊外形Fig.1 The stamen shape of Ranunculaceae
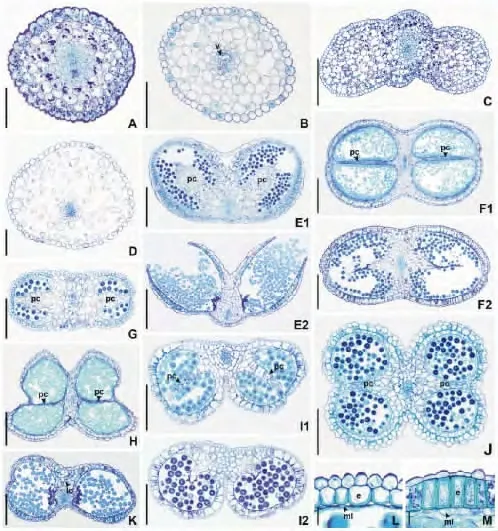
图2 毛茛科雄蕊结构Fig.2 The stamen structures of Ranunculaceae

表2 毛茛科雄蕊的形态及结构Table 2 The morphology and structure of Ranunculaceae stamens

续表2 Continued
成熟花药结构:成熟的花药壁中层及绒毡层几乎完全消失,仅具一层表皮细胞及一层木化细胞构成的药室内壁(图2L~M),木化细胞的外切向壁不木化(见于毛茛科多数属,图2L),或木化(仅见于乌头属和翠雀属,图2K)。每对花粉囊间的薄壁细胞随着雄蕊的成熟解体并消失,成为2室花药(图2E2,F2,I2,K)。成熟花药与幼嫩花药的横切面形态相似,如乌头属,毛茛属和翠雀属(图2E1,E2,2I1,I2,2K),或变化较大,如驴蹄草属和耧斗菜属幼嫩花药为∞形,成熟花药为椭圆形(图2F1,F2)。药隔附近表皮细胞通常与其内侧的细胞连接紧密(如耧斗菜属,图2F1,F2),但乌头属、银莲花属、侧金盏属和菟葵属二者分离(图2E1~E2)。药隔通常由薄壁细胞组成(如尖萼耧斗菜,图2F2),但翠雀属的药隔细胞木化(图2K)。
3 讨论
Decraene和Smets[13]指出雄蕊特征具有重要的系统学价值,且其外部形态特征可作为毛茛科部分类群组或亚组的划分依据。如Wang[24]曾依据成熟雄蕊表面是否被毛等特征将铁线莲属的菝葜叶铁线莲组(sect.Naraveliopsis)分为菝葜叶铁线莲亚组(subsect.Smilacifoliae)(雄蕊无毛),荔波铁线莲亚组(subsect.Liboenses)(雄蕊具毛)和Macgregorianae亚组(雄蕊无毛)。本研究显示,毛茛科物种花药的结构较为稳定,可作为种特征。而且幼嫩花药结构常比成熟花药结构具有更高的分类学价值,如位于不同属的侧金盏花和翠雀的成熟花药横切面虽均为∞形,但前者幼嫩花药为蝶形。这种变化是由花药发育过程中,花粉囊间的薄壁细胞逐渐变形,破裂并最终消失,以及药隔形态变化所致。
花药形态及结构可作为毛茛科多数属的分类特征,如乌头属花丝均为长三角形,花药侧面观∞形,幼嫩及成熟花药横切面V形;银莲花属花丝均为丝状,花药侧面观近圆形或椭圆形,幼嫩及成熟花药横切面椭圆形。但铁线莲属雄蕊的结构仅可作为亚组划分的依据,如 Wang[25],王文采和李良千[26]将短尾铁线莲及棉团铁线莲分别位于威灵仙组(sect.Clematis)的欧洲铁线莲亚组(subsect.Clematis)和棉团铁线莲亚组(subsect.Angustifoliae),二者花丝均为条形,但前者幼嫩花药横切面蝶形,花粉囊间薄壁细胞V形排列,成熟花药横切面∞形,后者幼嫩及成熟花药横切面为椭圆形,花粉囊间薄壁细胞排成直线形。花药结构支持 Wang[25],王文采和李良千[26]分类观点。
雄蕊特征不仅可作为属的鉴别依据,还可为系统学研究提供新的形态学证据。如水毛茛属、碱毛茛属和毛茛属的亲缘关系一直存在较大争议[2,27-30]。Benson[27]曾将前两个属作为毛茛属的亚属。另外一些学者也认为水毛茛属应作为毛茛属亚属[2,28-30]。但 Wang等[3],Takhtajan[4]依据果实及花的特征认为应保留水毛茛属、碱毛茛属和毛茛属的分类地位。本研究显示这3属的花丝虽均为短柱状,成熟花药横切面为∞形(水毛茛属及碱毛茛属)或Λ形(毛茛属),但幼嫩花药切面完全不同,水毛茛属为蝶形,碱毛茛属为方形,而毛茛属则为Λ形,支持Wang等[3]和 Takhtajan[4]的观点。唐松草属唐松草组(sect.Tripterium)所有物种及叉枝唐松草组(sect.Leptostigma)部分物种的花丝呈棒状[8],基本组织呈海绵状,其内薄壁组织的细胞排列多呈分支状,细胞间隙极大。这些特征不存在于其他物种。白新祥[31]认为基本组织细胞间具较大间隙使入射光线在花瓣内多次折射,令花瓣呈现白色。推测这也许是唐松草属部分物种棒状花丝呈现白色的原因。
本文首次揭示了毛茛科雄蕊的形态特征,幼嫩及成熟花药结构的变化,表明了雄蕊结构特别是幼嫩花药的特征可以作为属的分类依据,完善了毛茛科花的形态学研究,为进一步研究毛茛科分类及系统学提供了新的形态学基础。
Reference:
[1]The angiosperm phylogeny group.An update of the angiosperm phylogeny group classification for the orders and families of flowering.Botanical Journal of the Linnean Society,2009,161(2):105-121.
[2]Tamura M.Ranunculaceae[A].In:Kubitzki K,Rohwer J G,Bittrich V.The Families and Genera of Vascular Plants.Vol.2.Flowering Plants.Dicotyledons:Magnoliid,Hamameliid and Caryophyllid Families[M].Berlin:Springer-Verlag Berlin Heidelberg,1993:567-583.
[3]Wang W T,Fu D Z,Li L Q,etal.Ranunculaceae[A].In:Wu Z Y,Raven P H.Flora of China[M].Beijing:Science Press,St.Louis:Missouri Botanical Garden Press,2001:133-438.
[4]Takhtajan A.Flowering Plants[M].New York:Springer-Verlag New York Inc,2009:85-91.
[5]Sun T H,Liu M,Sun X Q,etal.Morphological study on the leaf structures ofPotentillain Northeastern China and its taxonomic value(Rosaceae).Acta Prataculturae Sinica,2014,23(3):75-84.
[6]Zhang X X,Liu M,Cheng X Y,etal.Comparative study of the morphological and anatomical features ofLinderniaprocumbensin different ecological environments(Linderniaceae).Acta Prataculturae Sinica,2014,23(2):235-242.
[7]Sun X Q,Liu M,Sun T H,etal.Morphological study of the leaf structures ofViolain northeastern China and discussions of their taxonomic values(Violaceae).Acta Prataculturae Sinica,2014,23(2):223-234.
[8]Delectis florae reipublicae popularis sinicae agendae academiae sinicae edita.Flora Reipublicae Popularis Sinicae(Vol.27)[M].Beijing:Science Press,1979.
[9]Delectis florae reipublicae popularis sinicae agendae academiae sinicae edita.Flora Reipublicae Popularis Sinicae(Vol.28)[M].Beijing:Science Press,1980.
[10]Takhtajan A L.Outline of the classification of flowering plants(magnoliophyta).The Botanical Review,1980,46(3):225-359.
[11]Liu L L,Yang X,Gao T P,etal.A study on the attractive function of different flora structures inTrolliusranunculoides(Ranunculaceae).Acta Prataculturae Sinica,2013,22(3):190-195.
[12]Hutchinson J.Contributions towards a phylogenetic classification of flowering plants.I.Bulletin of Miscellaneous Information(Royal Gardens,Kew),1923,(2):65-89.
[13]Decraene L P R,Smets E F.Evolution of the androecium in the Ranunculiflorae.Plant Systematics and Evolution,1995,9(Suppl.):63-70.
[14]Endress P K.The flowers in extant basal angiosperms and inferences on ancestral flowers.International Journal of Plant Sciences,2001,162(5):1111-1140.
[15]Song P,Tian X H,Ren Y.Floral morphogenesis ofCalthaandTrollius(Ranunculaceae)and its systematic significance.Acta Phytotaxonomica Sinica,2007,45(6):769-782.
[16]Gu T Q,Ren Y.Floral morphogenesis ofCoptis(Ranunculaceae).Chinese Bulletin of Botany,2007,24(1):80-86.
[17]Feng M,Fu D Z,Liang H X,etal.Floral morphogenesis ofAquilegiaL.(Ranunculaceae).Acta Botanica Sinica,1995,37(10):791-794.
[18]Tucker S C,Hodges S A.Floral ontogeny ofAquilegia,Semiaquilegia,andEnemion(Ranunculaceae).International Journal of the Plant Sciences,2005,166(4):557-574.
[19]Song C H.Morphology,structure,development and systematic ofSouliea(Ranunculaceae)[D].Xi’an:Shaanxi Normal University,2006.
[20]Ren Y,Chang H L,Tian X H,etal.Floral development in Adonideae(Ranunculaceae).Flora,2009,204(7):506-517.
[21]Kosuge K,Tamura M.Ontogenetic studies on petals of the Ranunculaceae.Journal of Japanese Botany,1989,64(3):65-74.
[22]Wang W T.A revision ofClematissect.Brachiatae(Ranunculaceae).Acta Phytotaxonomica Sinica,2004,42(4):289-332.
[23]Feder N,O’Brien T P.Plant microtechnique:some principles and new methods.American Journal of Botany,1968,55(1):123-142.
[24]Wang W T.A revision ofClematissect.Naraveliopsis(Ranunculaceae).Acta Phytotaxonomica Sinica,2006,44(6):670-699.
[25]Wang W T.A revision ofClematissect.Clematis(Ranunculaceae).Acta Phytotaxonomica Sinica,2003,41(1):1-62.
[26]Wang W C,Li L Q.A new system of classification of the genusClematis(Ranunculaceae).Acta Phytotaxonomica Sinica,2005,43(5):431-488.
[27]Benson L.The North Amerian subdivisions ofRanunculus.American Journal of Botany,1940,27(9):799-807.
[28]Dahlgren G.Karyological investigations inRanunculussubg.Batrachium(Ranunculaceae)on the Aegean islands.Plant Systematic and Evolution,1991,177(3-4):193-211.
[29]Diosdado J C,Pastor J E,Valdés B.Contributions to the karyological study of the genusRanunculusL.subgenusBatrachium(DC.)A Gray from the Iberian Peninsula.Botanical Journal of the Linnean Society,1993,112(1):75-87.
[30]Emadzade K,Lehnebach C,Lockhart P,etal.A molecular phylogeny,morphology and classification of genera of Ranunculeae(Ranunculaceae).Taxon,2010,59(3):809-828.
[31]Bai X X.Phenotype Analysis of Flower Coloration ofChrysanthemum×morifoliumRamat[D].Harbin:Northeast Forestry University,2007.
[5]孙天航,刘玫,孙雪芹,等.东北委陵菜属植物叶形态结构的研究及其分类学价值的探讨.草业学报,2014,23(3):75-84.
[6]张欣欣,刘玫,程薪宇,等.不同生境下陌上菜的形态解剖学比较.草业学报,2014,23(2):235-242.
[7]孙雪芹,刘玫,孙天航,等.东北堇菜属植物叶形态结构的研究及其分类学价值的探讨.草业学报,2014,23(2):223-234.
[8]中国科学院中国植物志编辑委员会.中国植物志(第27卷)[M].北京:科学出版社,1979.
[9]中国科学院中国植物志编辑委员会.中国植物志(第28卷)[M].北京:科学出版社,1980.
[11]刘乐乐,杨晓,高天鹏,等.毛茛状金莲花花部结构的吸引功能.草业学报,2013,22(3):190-195.
[15]宋萍,田先华,任毅.驴蹄草属和金莲花属(毛茛科)花器官的形态发生及系统学意义.植物分类学报,2007,45(6):769-782.
[16]辜天琪,任毅.黄连属(毛茛科)花的形态发生.植物学通报,2007,24(1):80-86.
[17]冯旻,傅德志,梁汉兴,等.耧斗菜属花部形态发生.植物学报,1995,37(10):791-794.
[19]宋春慧.黄三七属(毛茛科)植物的形态、结构、发育及系统位置研究[D].西安:陕西师范大学,2006.
[26]王文采,李良千.铁线莲属一新分类系统.植物分类学报,2005,43(5):431-488.
[31]白新祥.菊花花色形成的表型分析[D].哈尔滨:东北林业大学,2007.
猜你喜欢
伊犁河(2023年3期)2023-06-15 12:55:20
园林科技(2021年3期)2022-01-19 03:17:32
山西农业科学(2018年9期)2018-09-12 02:25:34
现代园艺(2017年22期)2018-01-19 05:06:56
花卉(2017年7期)2017-11-15 08:53:30
生物学教学(2017年6期)2017-02-18 10:14:33
课外语文·中(2016年12期)2017-01-03 23:43:55
红蜻蜓·低年级(2016年9期)2016-05-14 10:50:49
华南师范大学学报(自然科学版)(2014年6期)2014-08-28 00:43:38
天然产物研究与开发(2014年6期)2014-04-27 14:15:58
