
秦岭地区‘凤丹’牡丹居群果期相关性状的表型多样性研究
2015-01-02 10:19:50李林昊张延龙牛立新张晓骁任利益
西北林学院学报 2015年4期
李林昊,张延龙,牛立新,张晓骁,任利益
(西北农林科技大学 风景园林艺术学院,陕西 杨陵712100)
表型多样性主要研究居群在其分布区内各种环境下的表型变异,是遗传多样性与环境多样性的综合体现[1]。表型性状变化常常作为遗传变异的表征,因此如何根据表现型差异来反映基因型差异就成为形态学标记检测遗传多样性的关键[2]。通过有效的采样、合理的数学统计方法,采用遗传上较稳定的、不易受环境影响的性状,可以揭示群体的遗传规律、变异大小[3]。
牡丹(Paeonia suffruticosa)是芍药科(Paeoniaceae)芍药属(Paeonia)牡丹组(Sect.Mouton)的多年生落叶小灌木,是原产中国的一种名贵花卉[4],同时中国也是其栽培及遗传多样性的中心[5]。‘凤丹’牡丹是牡丹野生种杨山牡丹(Paeonia ostii)的栽培变种,主要应用于牡丹皮和籽油的生产,是一种很好的药食兼备植物,同时也是我国江南牡丹品种群的源种之一[6]。秦岭地区气候和地形地貌复杂多变,是我国多种植物的起源地,这其中就包括杨山牡丹[7]。虽然对该地区野生杨山牡丹的资源分布有报道[8-9],但迄今为止缺乏有关‘凤丹’牡丹资源变异情况的研究。
本研究选择陕西境内秦岭分布的8个具有代表性的‘凤丹’牡丹居群(含2个杨山牡丹居群)为对象,对其表型特征进行分析,旨在了解‘凤丹’牡丹的表型变异情况,揭示其变异程度、变异格局,为选择优良变异、合理开发利用资源供参考依据。
1 材料与方法
1.1 居群选择与样本采集
2013年7月下旬至8月上旬在陕西省境内秦岭‘凤丹’牡丹分布区内,抽取略阳(LY)、凤县(FX)、眉县(MX)、留坝(LB)、山阳(SY)、商州(SZ)、白河(BH)、商南(SN)等市县境内的8个居群,采样地地理生态因子、样点特征分别见表1、表2。入选居群必须保存完好,具备自然更新能力,植株无严重缺陷,无明显病虫害,且应为主要灌木种。在每个结实单株上采集不同方位的3片完整、成熟叶片及3个成熟但果荚未开裂果实。
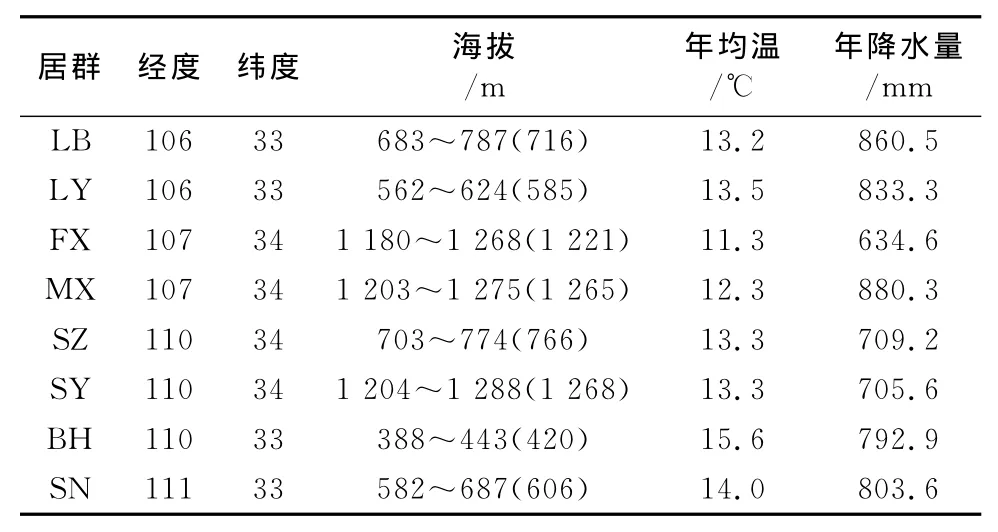
表1 ‘凤丹’牡丹8个居群的地理生态因子Table 1 Geo-ecological factors of P.ostii‘Feng Dan’in 8populations
1.2 性状选取测定与统计分析
该研究所选性状包括坐果量、果荚数、单果内种子数、果实横径、果实厚径、单果荚长度、单果荚宽度、种子径长、叶片长、叶片宽、叶柄长、二回叶长、二回叶宽、小叶数等14个数量性状,以及株形、分枝类型、叶柄颜色、叶形、果实颜色、果实是否有毛、果实饱满程度、种子饱满程度等8个质量性状。果实横径为果实平面最长处长度,果实厚径为果实立面最厚处长度,单果荚宽度为果荚最宽处长度,种子径长为种子直径平均长度,叶片长、宽均为二回羽状复叶的长、宽。叶片长、叶片宽、二回叶长、二回叶宽用卷尺测量,精度为0.1cm,其他需要测量的表型数量性状用游标卡尺测量,精度为0.1mm。每个居群随机测10个植株的性状数值,重复3次。对各性状值用SPSSv 18.0软件进行相关分析,其他统计分析按照常规方法并利用Excel 2003等提供的有关程序进行。
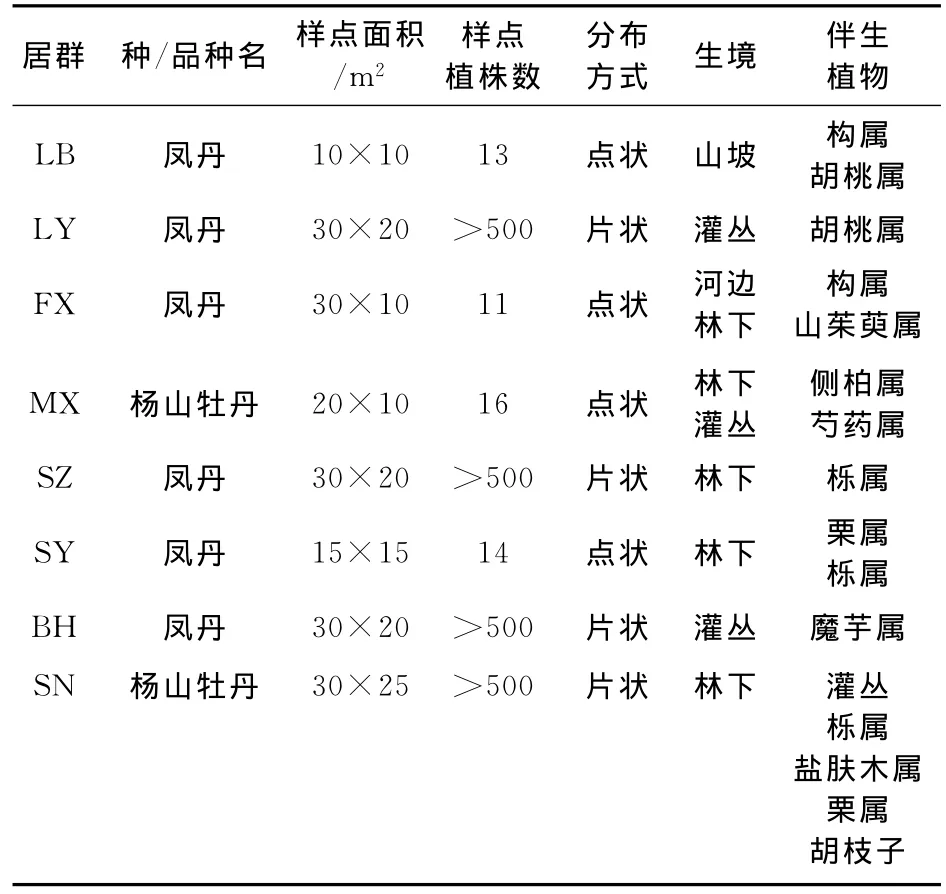
表2 ‘凤丹’牡丹8个居群的样点特征Table 2 Sample features of P.ostii‘Feng Dan’in 8populations
2 结果与分析
2.1 质量性状的变异
对不同居群的主要质量性状观察表明(表3),‘凤丹’牡丹8个居群的质量性状在居群内与居群间都存在着很明显的变异,其株形在居群间与居群内均存在变异,而分枝类型、新枝颜色、叶形、果实颜色、果实是否有毛、果实饱满程度、种子饱满程度等仅在居群间存在明显变异,在种群内则趋于一致性。其中,MX居群和SY居群内及居群间株形差别较大,有直立形、半开展形,这可能是由于种群密度不同所导致,密度大时牡丹生长空间有限,导致植株直立生长,密度小时,生长空间充足,植株则枝条开展;分枝类型均为单枝型;新枝颜色有绿色和绿色带紫红色条纹;叶形有大形长叶和中形叶;果实颜色由绿色到褐绿色过渡 ,这可能是由于不同种群受当地物候条件影响而导致果实成熟期不同;果实上均被短绒毛;果实饱满程度不一,除了SY居群果实有部分干瘪以外,其他居群果实均饱满,这可能与花期气候有关系;不同居群的种子均较饱满,SY居群尽管有些果实不饱满,但所结种子仍较饱满。
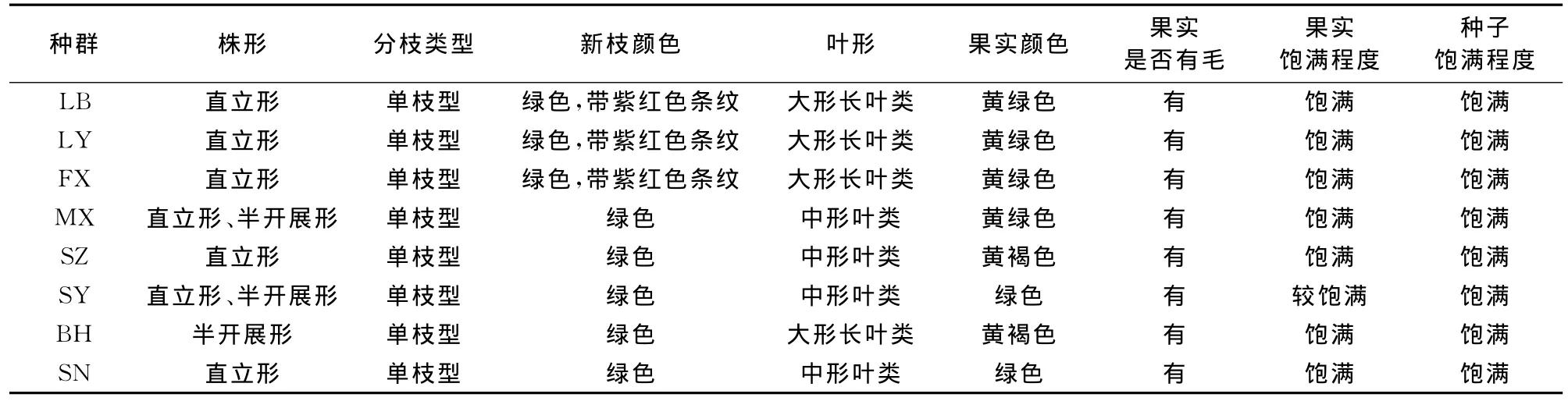
表3 ‘凤丹’牡丹8个天然种群的质量性状Table 3 Qualitative traits of‘Feng Dan’in 8populations
2.2 数量性状的变异
试验调查的杨山牡丹在居群内和居群间均存在显著或者极显著差异,说明这些性状在居群间与居群内均存在明显变异,而且变异不同。
2.2.1 均值比较 对8个居群内与居群间14个数量性状大小进行了测定比较(表4)。就果实性状而言,坐果量以LY居群的均值最大(15.0),SN居群的均值最小(1.7);果荚数在所有居群中差异不显著;单果内种子数以BH居群的均值最大(70.3),FX居群的均值最小(17.7),两者之间存在显著性差异;果实横径以SY居群的均值最大(118.3 mm),FX居群和MX居群的均值较小(84.7mm和86.5mm),且他们之间存在显著性差异,其余居群均值居中且其间差异不大;果实厚径以FX居群MX居群的均值较大(30.0mm和29.6mm),MX居群和SN居群的均值较小(16.9mm和16.5 mm),BH居群、LY居群、SZ居群、SY居群的均值居中且其间差异不大;单果荚长度在所有种群中均差异不大;单果荚宽度以BH居群的均值最大(19.7 mm),SZ居群的均值最小(14.5mm);种子径长在所有种群中均差异不大。就叶片性状而言,叶片长、叶片宽均以LY居群的均值较大(71.8cm和34.8 cm),SZ居群的均值最小(37.8cm和25.9cm),其他居群均值居中且其间差异不显著;叶柄长以BH居群的均值最大(226.2mm),SZ居群的均值最小(140.0mm),其他居群均值居中但差异不显著;二回叶长、二回叶宽分别以LB居群和SN居群的均值较大(2 6.7cm和1 5.7cm),但均以MX居群的均值最小,其他居群差异不显著;小叶数以LB居群和SN居群的均值最大(15.0),SZ居群的均值最小(10.0),其他居群居中且差异不大。

表4 ‘凤丹’牡丹8个居群数量性状的比较(平均值±标准误)Table 4 Comparison on quantitative traits of P.ostii‘Feng Dan’in 8populations(Mean±SE)
2.2.2 变异系数比较 变异系数反映性状在种群内与种群间的变异情况,变异系数越大,性状的离散程度越大,个体间的差异越大,性状越不稳定。‘凤丹’牡丹8个居群数量性状的变异系数见表5。从表5可以看出8个居群内数量性状的平均变异系数有一定的差异。结果显示:坐果量>单果内种子数>单果荚长度>果实厚径>二回叶宽>果荚数>叶柄长>叶片宽>二回叶长=果实横径>小叶数>叶片长>单果荚宽度>种子径长,说明不同数量性状的离散程度不同,稳定性也不同,并以种子径长最稳定,坐果量最不稳定。这些不同数量性状间变异系数的差异,可能是因为‘凤丹’牡丹在长期的自然选择条件下,不同数量性状对环境具有不同适应能力的结果。同时,‘凤丹’牡丹各个居群数量性状的平均变异系数表现为:BH居群>FX居群>SY居群>SZ居群>MX居群>LY居群>SN居群>LB居群,说明不同种群间的性状离散程度与稳定性不同,并以BH居群的性状最不稳定,LB居群的性状最稳定。
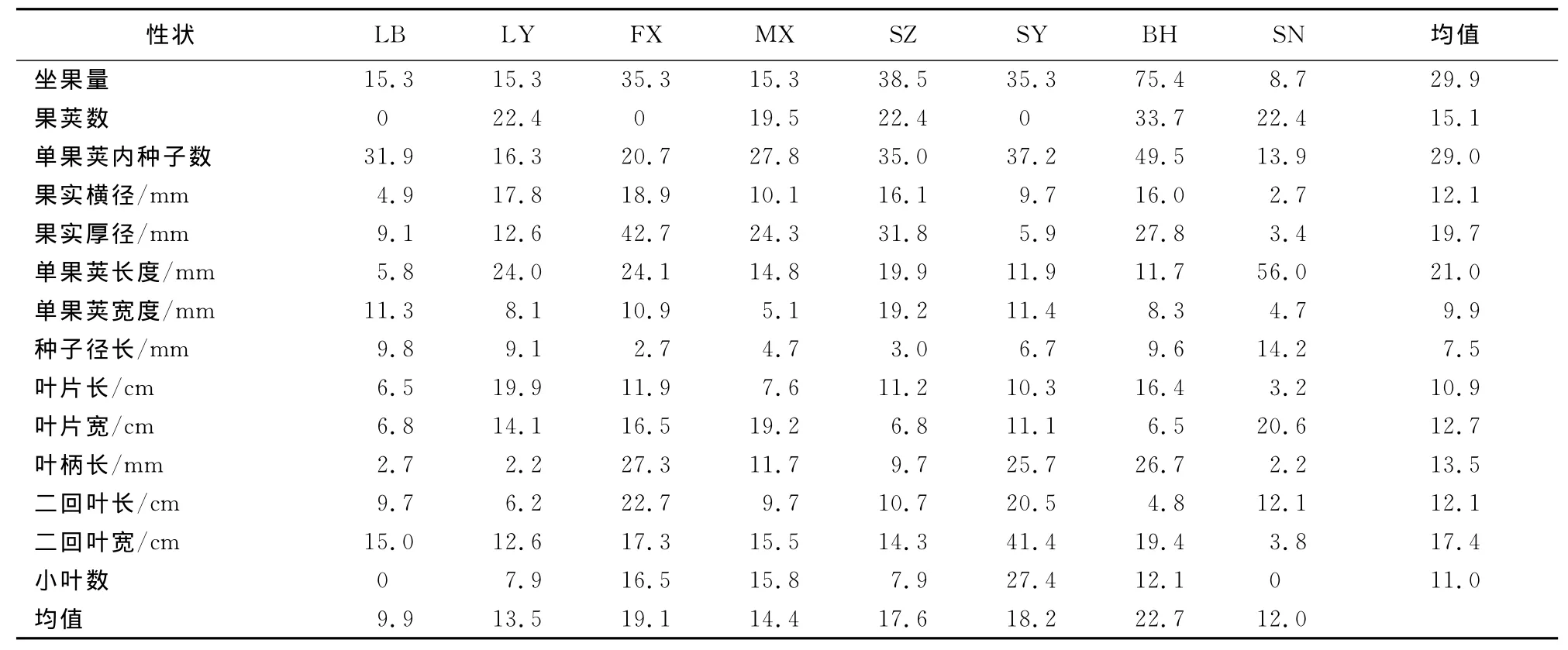
表5 ‘凤丹’牡丹8个种群数量性状的变异系数Table 5 Coefficients of variation of quantitative traits of 8populations in‘Feng Dan’
2.3 表型性状与地理因子的关系
对‘凤丹’牡丹不同居群14个数量性状与采样地的海拔、纬度、年均温及年降水量进行了相关分析和检验(表6)。结果表明,纬度、年均温与果荚数、单果内种子数、果实横径关系密切,且与单果内种子数达到极显著相关水平(-0.950**和0.861**);海拔与果荚数、单果内种子数、果实厚径关系密切,但均未达到显著相关水平;年降水量仅与坐果量关系密切,与其他性状只具有一般的相关性。可见,‘凤丹’牡丹居群数量性状变异受分布地的纬度和年均温影响较大,并以单果内种子数对纬度和年均温反应最敏感,与年均温呈极显著的正相关,与纬度则呈极显著的负相关;果荚数对经度反应最敏感,而年降水量则对各性状影响最小。
3 结论与讨论
从质量性状上看,‘凤丹’牡丹株形在居群内与居群间均存在变异外,其他表型性状只在居群间存在变异,且居群间的变异明显多于居群内。从数量性状上看,‘凤丹’牡丹居种群内与居群间均有显著变异,表型性状的离散程度高,居群间比居群内变异稍丰富,如8个居群的13个数量性状均值在种群间差异全部达到了极显著水平。种群间与种群内的变异同等重要[10],因为分布在居群间的变异反映了地理和生殖隔离上的差异,而居群内的变异是构成居群间变异的基础,也是居群内多样性的重要组成部分[11]。因此,‘凤丹’牡丹表型性状在居群间与居群内丰富的变异,都是其种质资源表型多样性的来源。

表6 ‘凤丹’牡丹表型性状与生态因子的相关性分析Table 6 Correlation of phenotypic characteristics and ecological factors of‘Feng Dan’
环境因素多样性是影响植物变异的重要因子之一,分布区的环境条件越复杂,则种群的表型变异越大[12-13]。试验所选择的8个居群,地理跨度较大,海拔、气压、年降水量及土壤条件都有差异,而且其分布呈间断的不连续,使得‘凤丹’牡丹的表型变异较大。本研究中‘凤丹’牡丹的单果内种子数对纬度和年均温的极显著相关性充分说明这点。
‘凤丹’牡丹作为当今油用牡丹产业发展的主要推广品种,其籽油中含有大量对人体健康有益的不饱和脂肪酸及微量元素[14-15],同时其高产量和强适应能力是其他牡丹品种所不能比的。通过对陕西省秦岭地区不同‘凤丹’牡丹居群果期相关性状的表型多样性分析,探讨环境因素对果实的影响,这对陕西油用牡丹产业的推广及发展具有重要参考价值。
[1] 杨生超,徐邵忠,文国松,等.灯盏花种质资源群体表型多样性研究[J].西北植物学报,2008,28(8):1573-1579.YANG S C,XU S Z,WEN G S,et al.Phenotypic diversity of populations in germplasm resources of Erigeron breviscapus[J].Acta Botanica Boreali-Occidentalia Sinica,2008,28(8):1573-1579.(in Chinese)
[2] 司国臣,张延龙,赵冰,等.太白杜鹃天然居群的表型多样性[J].西北植物学报,2012 ,32(8):1560-1566.SI G C ,ZHANG Y L ,ZHAO B ,et al.Phenotypic variation of natural populations in Rhododendron purdomii in Qinling Mountains[J].Acta Botanica Boreali-Occidentalia Sinica,2012,32(8):1560-1566.(in Chinese)
[3] 马玉敏,陈学森,何天明,等.中国板栗3个野生居群部分表型性状的遗传多样性[J].园艺学报,2008,35(12):1717-1726.MA Y M,CHEN X S,HE T M,et al.Genetic diversity of morphological traits in wild populations of Castanea mollissima Blume[J].Acta Horticulturae Sinica,2008,35(12):1717-1726.(in Chinese)
[4] 洪德元,潘开元.芍药属牡丹组的分类历史和分类处理[J].植物分类学报,1999,37(4):351-368.HONG D Y,PAN K Y.Taxonomical history and revision of Paeonia Sect.Moutan(Paeoniaceae)[J].Acta Phytotaxonomica Sinica,1999,37(4):351-368.(in Chinese)
[5] 蓝保卿,李嘉珏,段全绪.中国牡丹全书[M].北京:中国科学技术出版社,2002:1-4.
[6] 李嘉珏,张西方,赵孝庆.中国牡丹[M].北京:中国大百科全书出版社,2011:70-77.
[7] 李嘉珏.中国牡丹品种图志:西北·西南·江南卷[M].北京:中国林业出版社,2006:157-163.
[8] 洪涛,张家勋,李嘉珏,等.中国野生牡丹研究(一)芍药属牡丹组新分类群植物研究[J].植物研究,1992,12(3):223-234.HONG T,ZHANG J X,LI J J,et al.Study on the Chinese wild woody peonies(Ⅰ)new taxa of paeonia L.Sect.moutan DC [J].Bulletin of Botanical Research,1992,12(3):223-234.(in Chinese)
[9] 王佳.杨山牡丹遗传多样性与江南牡丹品种资源研究[D].北京:北京林业大学,2009:22.
[10] 辜云杰,罗建勋,吴远伟,等.川西云杉天然种群表型多样性[J].植物生态学报,2009,33(2),291-301.GU Y J,LUO J X,WU Y W,et al.Phenotypic diversity in natural populations of Picea balfouriana in Sichuan,China[J].Chinese Journal of Plant Ecology,2009,33(2),291-301.(in Chinese)
[11] 佘诚棋,杨万霞,方升佐,等.青钱柳天然群体种子性状表型多样性[J].应用生态学报,2009,20(10):2351-2356.SHE C Q,YANG W X,FANG S Z,et al.Phenotypic diversity of natural Cyclocaryapaliurus populations seed traits[J].Chinese Journal of Applied Ecology,2009,20(10):2351-2356.(in Chinese)
[12] 肖海峻,徐柱,翟利剑,等.鹅观草居群表型多样性研究[J].西北植物学报,2007,27(11):2222-2227.XIAO H J,XU Z,ZHAI L J,et al.Phenotypic diversity of Roegneria kamoji Ohwi population[J].Acta Botanica Boreali-Occidentalia Sinica,2007,27(11):2222-2227.(in Chinese)
[13] 李宗艳,张海燕.黄牡丹表型变异及多样性研究[J].西北林学院学报,2011,26(4):117-122.LI Z Y,ZHANG H Y.Morphological variation and diversity in populations of Paeonia lutea[J].Journal of Northwest Forestry University,2011,26(4):117-122.(in Chinese)
[14] 翟文婷,朱献标,李艳丽,等.牡丹籽油成分分析及其抗氧化活性研究[J].烟台大学学报:自然科学与工程版,2013,26(2):147-150.ZHAI W T,ZHU X B,LI Y L,et al.Chemical constituents and antioxidant activity of peony seed oil[J].Journal of Yantai University:Natural Science and Engineering Edition,2013,26(2):147-150.(in Chinese)
[15] 周海梅,马锦琦,苗春雨,等.牡丹籽油的理化指标和脂肪酸成分分析[J].中国油脂,2009,34(7):72-74.
猜你喜欢
特产研究(2022年6期)2023-01-17 05:06:02
现代农业科技(2022年18期)2022-09-27 12:05:38
东北农业大学学报(2022年7期)2022-09-23 02:40:40
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
植物资源与环境学报(2020年4期)2020-08-10 05:23:14
河北果树(2020年2期)2020-05-25 06:58:20
草地学报(2018年5期)2018-11-07 02:25:00
农家之友(2018年7期)2018-07-19 08:13:42
天然产物研究与开发(2018年3期)2018-05-07 06:38:52
