
嗜热侧孢霉2441纤维素酶产酶条件优化及突变株选育
2014-12-31 12:01:04陶树兴魏园媛冯晓磊许真珍
陕西师范大学学报(自然科学版) 2014年6期
陶树兴,魏园媛,梁 健,冯晓磊,熊 平,唐 娜,许真珍
(陕西师范大学 生命科学学院,陕西 西安 710119)
纤维素是自然界中蕴藏量巨大且可再生的重要生物资源,天然纤维素以高度结晶的形式存在,得到高活力低成本的纤维素酶将其降解成单糖是纤维素资源高效利用的关键.纤维素酶是一种多组分混合酶系,按其催化功能可分外切酶、内切酶和β-葡萄糖苷酶三类,只有当三者活性比例恰当,才能表现出较高的纤维素酶活力[1-2].纤维素酶在食品、酒精、饲料和纺织等领域具有巨大的市场潜力.纤维素酶主要来源于微生物,由于丝状真菌能将纤维素酶分泌到细胞外,目前工业上生产纤维素酶的微生物大多属于真菌,如曲霉属(Aspergillus)、木霉属(Trichoderma)、青霉属(Penicillium)、枝顶孢霉属(Acremonium)等[3-6].嗜 热 侧 孢 霉 (Sporotrichum thermophile)是一类能产生纤维素酶混合酶系的嗜热微生物,骆跃军等研究发现,嗜热侧孢霉在以稻草、麸皮为碳源的培养基中发酵培养时,其纤维素酶活是曲霉的2倍,但比瑞氏木霉(Trichodermareesei)低[7].而Bhat等发现嗜热侧孢霉在50℃左右分解纤维素的速率是瑞氏木霉等常温菌株的5倍,瑞氏木霉14~18d产生的酶量,嗜热侧孢霉3~4d即可产生[8].在嗜热侧孢霉菌种选育方面,张辉、许燕等都取得了较好的结果[9-10].已有研究表明,纤维素酶的产生受葡萄糖效应阻遏[11],嗜热侧孢霉中有无此限制仍需进行探讨.另外,嗜热侧孢霉的耐高温特性赋予其所产的纤维素酶热稳定性高、培养过程中不易被杂菌污染等优点[12],但培养温度高使得工业生产耗能多,生产成本增加.本实验在研究嗜热侧孢霉纤维素酶的产酶条件及酶特性的基础上,选育抗葡萄糖效应突变株和中温型突变株,对提高纤维素酶的生成量、降低培养温度以降低生产成本以及扩大该菌株应用范围等方面都有重要的实践意义.
1 材料和方法
1.1 实验菌株
嗜热侧孢霉(Sporotrichumthermophile)2441,由中国工业微生物菌种保藏管理中心提供.
1.2 培养基
1.2.1 斜面种子培养基 采用PDA培养基[13].
1.2.2 产酶基础培养基 微晶纤维素10g,麸皮3.0g,葡萄糖1.0g,酵母膏1.0g,NaNO32.0g,K2HPO41.0g,KCl 0.5g,MgSO4·7H2O 0.5g,FeSO40.01g,MnSO40.005g,ZnSO40.002g,去离子水1 000mL,pH 6.0,113℃灭菌20min.
1.2.3 初筛培养基
(1)七叶苷筛选培养基[14].
(2)微晶纤维素刚果红筛选培养基(双层平板法) 底层培养基:酵母膏1.0g,NaNO32.0g,K2HPO41.0g,KCl 0.5g,MgSO4·7H2O 0.5g,FeSO40.01g,MnSO40.005g,ZnSO40.002g,脱氧胆酸钠2.0g,琼脂20g,去离子水1 000mL,pH 6.0,113℃灭菌20min.上层培养基:在底层培养基配方中加入湿磨的微晶纤维素2.0g(按干重计算),刚果红0.25g.
1.3 粗酶液制备
将嗜热侧孢霉2441接种于PDA斜面培养基,45℃培养5d,每支斜面用5mL无菌水洗下孢子,用移液枪吹打,经无菌脱脂棉过滤制得孢子悬液并调整其浓度为108个/mL.以4%的接种量将孢子悬液接入装有50mL产酶培养基的250mL三角瓶中,45℃,160r/min振荡培养5d(通过预备试验确定),将发酵液于10 000r/min离心10min,取上清液即为粗酶液.
1.4 纤维素酶活力测定
以1%的微晶纤维素(160r/min湿磨48h)悬液、CMC-Na溶液、水杨苷溶液为底物,60℃反应30 min(依据反应初速度确定),DNS法测定酶水解底物产生的还原糖量计算微晶纤维素酶活力、CMCNa酶活力和β-葡萄糖苷酶活力,酶活力单位(U)定义为每分钟水解底物产生1μmoL葡萄糖所需的酶量[15-19].
1.5 嗜热侧孢霉2441纤维素酶产酶条件优化
按L9(34)正交表安排试验,以微晶纤维素酶活力为考核指标,探讨产酶培养基中碳源、氮源、无机盐、微量元素种类及含量的最优组合和最优培养条件,每个试验重复3次.
1.6 酶特性研究
1.6.1 酶作用最适温度 分别在50、55、60、65、70、75℃下测定纤维素酶活力,每个处理重复3次.
1.6.2 酶作用最适pH 分别配制pH为4.0、4.5、5.0、5.5的乙酸-乙酸钠缓冲液和pH 为6.0、6.5、7.0、7.5、8.0的磷酸缓冲液,并用相应pH 的缓冲液配制底物溶液,在最适作用温度下测定纤维素酶活力,每个处理重复3次.
1.6.3 酶的热稳定性 将粗酶液分别于40、50、60、70、80、90℃下保温120min,取各温度保温后的酶液于最适作用温度和最适作用pH系统中,测定剩余酶活力,以未处理的酶液在最适温度和pH下测定的酶活力为100%作为对照,每个处理重复3次.
1.6.4 酶对pH的稳定性 将粗酶液与pH值为4.0~8.0的缓冲液等体积混合,在4℃冰箱中放置24h后于最适温度、最适pH下测定剩余酶活力,以未处理的酶液在最适温度和pH下测定的酶活力为100%作为对照,每个处理重复3次.
1.7 突变株选育
1.7.1 NTG诱变和抗葡萄糖效应突变株的选育取适当萌发的孢子悬液1.5mL,加入0.5mL NTG溶液(4mg/mL),28℃恒温振荡处理5、10、15、20、25、30、35、40、45、50min后,离心,用生理盐水洗涤2次除去NTG溶液,适当稀释,涂布于含5%葡萄糖的七叶苷筛选平板,45℃培养5~7d.在70%~80%致死率范围内(NTG处理20min),挑取棕色较深且显色圈与菌落直径比值较大的菌株移接斜面,45℃培养5d,转接于优化后且加有高浓度葡萄糖(5g/L)的产酶培养基,45 ℃、170r/min振荡培养5d,分别测定微晶纤维素酶、CMC-Na酶和β-葡萄糖苷酶活力,选择酶活提高幅度最大的突变株连续转接5代,测定第1、3、5代菌株的三种纤维素酶活,检测其遗传稳定性.
1.7.2 UV诱变和中温型突变株的选育 取适当萌发的孢子悬液5mL于放有大头针的无菌平皿中,将平皿置于15W紫外灯下30cm处的磁力搅拌器上,分别照射10、20、40、60、90、120、180、240s,在红光下适当稀释,涂布于微晶纤维素刚果红筛选平板,将平板用黑布包严置32℃培养5~7d,在70%~80%致死率范围内(UV照射60s),挑取长势良好且透明圈与菌落直径比值较大的菌株移接斜面,32℃培养5d,转接于优化后的产酶培养基,32℃、170r/min振荡培养5d,分别测定微晶纤维素酶、CMC-Na酶和β-葡萄糖苷酶活力,选择酶活提高幅度最大的突变株连续转接5代,测定第1、3、5代菌株的三种纤维素酶活,检测其遗传稳定性.
2 结果分析
2.1 嗜热侧孢霉2441纤维素酶产酶条件优化
2.1.1 碳源对嗜热侧孢霉2441纤维素酶活的影响采用L9(34)正交表考查微晶纤维素含量(A1=10.0 g/L,A2=15.0g/L,A3=20.0g/L)、麸皮含量(B1=0.0g/L,B2=1.5g/L,B3=3.0g/L)和葡萄糖含量(C1=0.0g/L,C2=2.0g/L,C3=5.0g/L)对嗜热侧孢霉2441微晶纤维素酶活的影响,其中4号试验(A2B1C2)酶活最高,为49.843±0.664U/mL.
方差分析结果表明,微晶纤维素(F=4.276,P>0.05)和麸皮(F=1.260,P>0.05)含量对酶活的影响差异不显著,葡萄糖含量差异显著(F=38.768,P<0.05).由极差分析可以看出,3种碳源影响的主次顺序是C>A>B,葡萄糖对微晶纤维素酶的产生影响最大;最佳组合为A2B2C2,即微晶纤维素15.0g/L,麸皮1.5g/L,葡萄糖2.0g/L.适量的葡萄糖有利于嗜热侧孢霉的生长和产酶,而葡萄糖浓度偏高时,纤维素酶活下降,这说明在嗜热侧孢霉中也存在有葡萄糖阻遏效应,这为选育抗葡萄糖效应突变株提供了依据.
2.1.2 氮源对嗜热侧孢霉2441纤维素酶活的影响在碳源优化的基础上,采用L9(34)正交表考查(NH4)2SO4含量(A1=0,A2=1.0g/L,A3=2.0 g/L)、NaNO3含量(B1=0,B2=1.0g/L,B3=2.0 g/L)和酵母膏含量(C1=0,C2=0.5g/L,C3=1.0 g/L)对嗜热侧孢霉2441微晶纤维素酶活的影响,其中7号试验(A3B1C3)酶活最高,为53.896±0.461U/mL.
方差分析结果表明,(NH4)2SO4(F=46.576,P<0.05)含量对酶活的影响差异显著,酵母膏(F=114.969,P<0.01)含量差异极显著,硝酸钠含量差异不显著(F=19.191,P>0.05).由极差分析可以看出,三种氮源影响的主次顺序是C>A>B,最佳组合为A3B1C3,即(NH4)2SO42.0g/L,酵母膏1.0 g/L,不加硝酸钠.
2.1.3 无机盐对嗜热侧孢霉2441纤维素酶活的影响 在碳源和氮源优化的基础上,采用L9(34)正交表探讨K2HPO4含量(A1=0,A2=1.0g/L,A3=2.0g/L)、KCl含量(B1=0,B2=0.5g/L,B3=1.0 g/L)和 MgSO4·7H2O含量(C1=0,C2=0.5g/L,C3=1.0g/L)对嗜热侧孢霉2441微晶纤维素酶活的影响,其中6号试验(A2B3C1)酶活最高,为53.897±0.835U/mL.
方差分析结果表明,K2HPO4(F=19.102,P<0.05)和KCl(F=25.741,P<0.05)含量对酶活的影响差异显著,MgSO4·7H2O含量差异不显著(F=2.456,P>0.05).由极差分析可以看出,三种无机盐影响的主次顺序是B>A>C,最佳组合为A2B3C3,即 K2HPO42.0g/L,KCl 1.0g/L,MgSO4·7H2O 1.0g/L.
2.1.4 微量元素对嗜热侧孢霉2441纤维素酶活的影响 在碳源、氮源、无机盐种类和含量优化的基础上,采用L9(34)正交表探讨微量元素FeSO4含量(A1=0,A2=0.010g/L,A3=0.020g/L)、MnSO4含量(B1=0,B2=0.005g/L,B3=0.010g/L)和ZnSO4含量(C1=0,C2=0.002g/L,C3=0.004 g/L)对嗜热侧孢霉2441微晶纤维素酶活的影响,其中5号试验(A2B2C3)酶活最高,为55.704±0.623 U/mL.
方差分析结果表明,FeSO4(F=21.648,P<0.05)含量对酶活的影响差异显著,MnSO4(F=3.496,P>0.05)和ZnSO4(F=7.124,P>0.05)差异不显著.由极差分析可以看出,三种微量元素影响的主次顺序是A>C>B,最佳组合为A2B2C2,即FeSO40.01g/L,MnSO40.005g/L,ZnSO40.002 g/L.
2.1.5 培养条件对嗜热侧孢霉2441纤维素酶活的影响 在培养基组成优化的基础上,采用L9(34)正交表考察培养基初始pH(A1=5.0,A2=5.5,A3=6.0)、培养温度(B1=45 ℃,B2=50 ℃,B3=55℃,)、振荡频率(C1=160r/min,C2=170r/min,C3=180r/min)对嗜热侧孢霉2441微晶纤维素酶活的影响,通过预备试验确定培养时间为5d,其中4号试验(A2B1C2)酶活最高,为56.362±0.442 U/mL.
方差分析结果表明,培养温度(F=32.995,P<0.05)和培养基初始pH(F=19.707,P<0.05)对酶活的影响差异显著,振荡频率(F=9.073,P>0.05)对酶活的影响差异不显著.由极差分析可以看出,各培养条件影响的主次顺序是B>A>C,最佳组合为A1B1C2,即培养基初始pH 为5.0,培养温度为45℃,振荡频率为170r/min.
2.1.6 验证试验 采用优化前后的培养基及培养条件分别培养嗜热侧孢霉2441,重复3次,5d后测定微晶纤维素酶活、CMC-Na酶活以及β-葡萄糖苷酶活.方差分析结果表明,产酶条件优化后,三种纤维素酶活力极显著提高(P<0.01),其中微晶纤维素酶活达到56.774U/mL,与优化前45.684U/mL相比,酶活提高 24.275%;CMC-Na酶活达到68.705U/mL,与优化前58.052U/mL相比,酶活提高18.35%;β-葡萄糖苷酶活达到59.473U/mL,与优化前48.176U/mL相比,酶活提高23.449%.
2.2 嗜热侧孢霉2441纤维素酶的特性
嗜热侧孢霉2441纤维素酶作用的最适温度、最适pH、酶的热稳定性和酶对pH的稳定性如图1所示.微晶纤维素酶、CMC-Na酶和β-葡萄糖苷酶作用的最适温度分别为60、65和70℃,最适pH均为5.5,温度高于70℃及碱性条件下,酶活力显著下降.

图1 嗜热侧孢霉2441纤维素酶的特性:酶作用的最适温度(a)、酶作用的最适pH(b)、酶的热稳定性(c)、酶对pH的稳定性(d)Fig.1 The characteristics of cellulase produced by Sporotrichum thermophile 2441:the change of activity with temperature(a),the change of activity with pH(b),the change of relative activity with temperature(c)and the change of relative activity with pH(d)
2.3 嗜热侧孢霉2441抗葡萄糖效应突变株的选育
七叶苷初筛平板上显色圈与菌落直径比值较大的10个菌株,微晶纤维素酶、CMC-Na酶和β-葡萄糖苷酶活及方差分析结果见表1.由表1可见,在高浓度葡萄糖存在的条件下,N20-01三种纤维素酶活力最高,与出发菌株2441相比,微晶纤维素酶活提高了13.203%,CMC-Na酶活提高了47.176%,β-葡萄糖苷酶活提高了119.517%.将N20-01转接5代,测定第1、3、5代3种纤维素酶的酶活,方差分析表明酶活均无显著差异,说明N20-01遗传特性稳定.
2.4 嗜热侧孢霉2441中温型突变株的选育
选择微晶纤维素刚果红初筛平板上透明圈与菌落直径比值较大的菌株,接入优化后的产酶培养基于32℃培养5d,其中10个菌株的微晶纤维素酶、CMC-Na酶和β-葡萄糖苷酶活及方差分析结果见表2.表中对照组嗜热侧孢霉2441在45℃培养5d.由表2可见,U40-03的三种纤维素酶活力最高,与其余突变株之间的差异极显著.U40-03与对照组相比,微晶纤维素酶活提高了11.411%,CMC-Na酶活较出发株提高了29.484%,β-葡萄糖苷酶活提高了54.759%.将 U40-03转接5代,测定第1、3、5代3种纤维素酶的酶活,方差分析表明酶活均无显著差异,说明U40-03遗传特性稳定.

表1 嗜热侧孢霉抗葡萄糖效应突变株复筛结果Tab.1 The secondary screening results of glucose resistant mutants of Sporotrichum thermophile
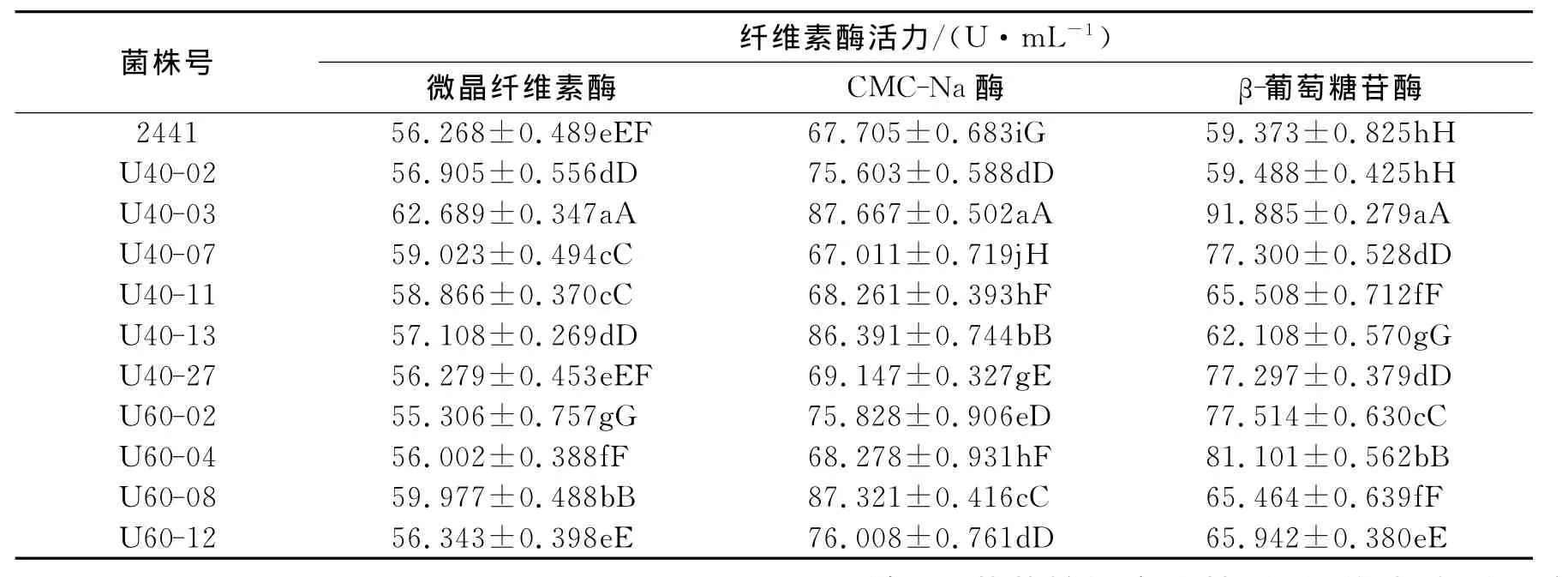
表2 嗜热侧孢霉中温型突变株复筛结果Tab.2 The secondary screening results of mesophilic mutants of Sporotrichum thermophile
3 讨论
在产酶培养基优化正交试验中,当培养基中葡萄糖添加量为2g/L时,嗜热侧孢霉2441纤维素酶活较高,但葡萄糖浓度升高到5g/L时,酶活反而下降,这说明嗜热侧孢霉2441也存在类似斜卧青霉等其他纤维素酶产生菌中的葡萄糖阻遏效应[11].在筛选抗葡萄糖效应突变株时,曾用微晶纤维素-刚果红-葡萄糖平板进行初筛,结果透明圈与菌落直径比值大小与酶活的相关性较差;采用微晶纤维素-刚果红-2-脱氧-D-葡萄糖平板进行初筛,虽然筛选到抗2-脱氧-D-葡萄糖的突变株,但纤维素酶活不高;改用七叶苷-柠檬酸铁-葡萄糖平板进行初筛,结果显色圈与菌落直径比值大小与β-葡萄糖苷酶活力有明显的正相关性,选育的抗葡萄糖效应突变株N20-01,β-葡萄糖苷酶活提高了119.517%.
嗜热侧孢霉2441虽是一株嗜热菌,但实验发现它在28~40℃也可以生长,28℃培养5d时,只长出白色菌丝体,不产生孢子.本研究得到的中温型突变株U40-03,在28℃培养5d时能产生孢子,进一步研究发现,U40-03的纤维素酶的最适产酶温度下降至32℃,且微晶纤维素酶活、CMC-Na酶活和β-葡萄糖苷酶活均有显著提高.在纤维素酶工业化生产中,菌株培养温度的降低可减少工业生产中由于升温、保温所需的能源,降低生产成本;不同季节使用不同菌株进行生产,可减少环境温度对菌株的限制,一年四季都可生产;将同一菌种的中温型菌株与高温型菌株混合使用,在培养前期中温型菌株大量生长并产热,可为中期高温型菌株的生长创造条件,两种类型的菌株协调作用,可降低能源消耗.目前,嗜热侧孢霉已广泛用作生产秸秆腐熟菌剂的菌株,在有机肥堆肥时,中温型和高温型菌株混合使用,升温加快,有利于物料腐熟;在秸秆还田中,使用中温型突变株能更好的适应土壤环境.另外,该菌株纤维素酶的最适产酶温度降低,也可应用于食品加工领域.
本试验只对出发株进行了一轮诱变,如进行多轮诱变或将不同方法及不同策略选育的突变株进行多亲本递进式原生质体融合,有可能得到酶活进一步提高的菌株.突变菌株产酶条件和酶特性有无变化正在进行研究.
4 结论
本研究得出以下结论:(1)通过正交试验优化后的培养基组成为:微晶纤维素15.0g/L、麸皮1.5 g/L、酵母膏1.0g/L、葡萄糖2.0g/L、(NH4)2SO42.0g/L、KCl 1.0g/L、K2HPO41.0g/L、FeSO40.01g/L、MgSO41.0g/L、MnSO40.05g/L、ZnSO40.002g/L;产酶最适培养条件为:培养基起始pH 5.0,培养温度45℃、摇床转速170r/min、培养时间为5d;培养基和培养条件优化后,嗜热侧孢霉2441的微晶纤维素酶活提高了24.275%;CMC-Na酶活提高了18.35%;β-葡萄糖苷酶活提高了23.449%.(2)嗜热侧孢霉2441产生的微晶纤维素酶、CMC-Na酶和β-葡萄糖苷酶,最适作用温度分别为60℃、65℃、70℃,最适作用pH均为5.5,温度高于70℃及碱性条件下,酶活力下降明显.(3)嗜热侧孢霉2441纤维素酶生成存在葡萄糖阻遏效应,选育到抗葡萄糖效应突变株N20-01,微晶纤维素酶活提高了13.203%,CMC-Na酶活提高了47.176%,β-葡萄糖苷酶活提高了119.517%.(4)选育到中温型突变株 U40-03,微晶纤维素酶活提高了11.411%,CMC-Na酶活提高了29.484%,β-葡萄糖苷酶活提高了54.759%.
[1]曲音波.木质纤维素降解酶与生物炼制[M].北京:化学工业出版社,2011:94-96,211-217.
[2]吴琳,景晓辉,黄俊生,等.产纤维素酶菌株的分离·筛选及酶活性测定[J].安徽农业科学,2009,37(17):7855-7857,7859.
[3]杜娟,曲音波,林觐勤,等.灰绿曲霉高产纤维素酶突变株的选育[J].厦门大学学报:自然科学版,2006,45(S1):23-26.
[4]He Jun,Yu Bing,Zhang Keying,et al.Strain improvement ofTrichodermareeseiRut C-30for increased cellulase production[J].Indian Journal of Microbiology,2009,49:188-195.
[5]Cheng R,Song X,Qin Y,et al.Genome shuffling improves production of cellulase byPenicilliumdecumbensJU-A10[J].Journal of Applied Microbiology,2009(10):1365-2672.
[6]Fang X,Yano S,Inoue H,et al.Lactose enhances cellulose production by the filamentous fungusAcremonium Cellulolyticus[J].Journal of Bioscience and Binengineering,2008(107):256-261.
[7]骆跃军,陈育如,李雪梅.纤维降解高温微生物的产酶特性研究[J].林产化学与工业,2005,25(S1):55-58.
[8]Bhat K M,Maheshwari R.S.thermophile:growth,cellulose degradation,and cellulase activity[J].Applied and Environmental Microbiology,1987,53:2175-2182.
[9]张辉,朱奇,杨启银,等.嗜热纤维素分解菌TH3-9诱变选育及产酶条件初步研究[J].安徽农业科学,2006,34(11):2318-2320.
[10]许燕.高内切葡聚糖酶活嗜热侧袍霉突变菌株的筛选[D].福州:福建农林大学生命科学学院,2008:48-52.
[11]曲音波,高培基,王祖农.青霉的纤维素酶抗降解物阻遏突变株的选育[J].真菌学报,1984,3(4):238.
[12]Argyrios Margaritis,Edward Creese.Thermal stability characeristics of cellulose enzymes produced bySporotrichumthermophile[J].Biotechnology Letters,1981,9(3):471-476.
[13]沈萍,陈向东.微生物学实验[M].4版.北京:高等教育出版社,2007:242.
[14]袁晓华.葡萄糖苷酶产生菌的筛选-培养条件优化及葡萄糖苷酶应用研究[D].济南:山东大学生命科学学院,2009:13-18.
[15]Ghose T K.Measurement of cellulose activities[J].Pure and Applied Chemistry,1987,59:257-268.
[16]Horikoshi K,Nakao M,Kurono Y,et al.Cellulase of an alkalophilic Bacillus strain isolated from soil[J].Canadian Journal of Microbiology,1984,30:774-779.
[17]Christakopoulos P,Goodenough P W,Kekos D.Purification and characterization of an extracellularβ-glucosidase with transglycosylation and exo-glucosidase activites fromFusariumoxysporum[J].European Journal of Biochemistry,1994,224:378-385.
[18]董志扬,祝令香,于巍,等.纤维素酶高产菌株的诱变选育及产酶条件研究[J].核农学报,2001,15 (1):26-31.
[19]王洪媛,范丙全.三株高效秸秆纤维素降解真菌的筛选及其降解效果[J].微生物学通报,2010,50(7):870-875.
猜你喜欢
土壤学报(2022年1期)2022-03-08 08:52:56
建材发展导向(2021年11期)2021-07-28 06:57:06
食用菌(2019年4期)2019-08-27 06:34:58
食品与生物技术学报(2017年2期)2017-04-09 11:43:29
山东工业技术(2016年15期)2016-12-01 05:31:50
天然产物研究与开发(2016年6期)2016-06-05 10:29:27
中国酿造(2016年12期)2016-03-01 03:08:19
超硬材料工程(2016年1期)2016-02-28 22:20:07
应用海洋学学报(2015年3期)2015-11-22 07:39:30
四川师范大学学报(自然科学版)(2015年1期)2015-02-28 14:07:30
