
极限糊精对小麦淀粉回生的影响
2014-12-25 02:28徐学明金征宇范雪荣
食品与生物技术学报 2014年11期
徐 进, 徐学明, 金征宇, 王 强, 范雪荣
(1.江南大学 纺织服装学院,江苏 无锡 214122;2.江南大学 食品学院,江苏 无锡 214122)
淀粉回生是糊化后的淀粉分子链经氢键缔合形成的双螺旋结构趋于有序化形成晶体的过程,它是米饭、面包、糕点等淀粉基食品在贮藏过程中变硬、干缩、掉渣等问题的主因[1]。因此,淀粉回生导致食品可接受程度降低和巨大的经济损失,是制约我国主食工业化的主要问题之一。
利用淀粉酶来抑制淀粉回生,一直是研究淀粉抗回生应用的热点。但从文献报道α-淀粉酶抑制面包老化的机理来看,该机理目前还存在一些争议。Miyazak等[2]报道在制作面包过程中加入小相对分子质量的糊精可抑制淀粉的老化,但小相对分子质量糊精的添加与面包变硬没有潜在的关系。Hug-Iten等[3]对实验结果的分析则认为:α-淀粉酶抑制面包老化的效果是由于淀粉的部分水解导致结构变化所引起。 然而,León等[4]研究认为,α-淀粉酶延缓面包老化并不是水解改变其结构,而是水解产物(糊精)起到延缓面包老化的作用,因为León等在加入糖化酶(glucoamylase)除去糊精后,面包老化延缓的效果就随之消失。Akers和Hoseney的研究结果[5]也证实,α-淀粉酶抑制面包变硬效果与α-淀粉酶水解淀粉所产生的麦芽糊精相对分子质量大小和比例都密切相关。此外,Witczak等[6]研究结果表明,高DE值(尤其是DE 18.0和DE 21.8)的麦芽糊精能够有效降低淀粉老化速率。因此,α-淀粉酶抑制淀粉回生研究结果的不一致说明目前对α-淀粉酶抑制面包老化的机理尚未研究透彻。
虽然文献报道α-淀粉酶水解淀粉生成的糊精延缓淀粉的回生,但并未详细研究是直链还是支链的糊精片段延缓淀粉回生。前期研究结果发现,直链糊精片段可以影响淀粉回生程度[7],但分支结构的糊精片段是否影响淀粉回生尚待考察。作者通过添加极限糊精到小麦淀粉中,考察分支结构的糊精片段对小麦淀粉回生的影响,并结合分子动力学模拟的方法为α淀粉酶抑制淀粉回生机理提供了理论依据。
1 材料与方法
1.1 材料与试剂
1.1.1 材料 小麦淀粉:购于天津顶峰淀粉开发有限公司。
1.1.2 试剂 α-淀粉酶(来源于地衣芽孢杆菌)、β-淀粉酶(来源于大麦):购于美国Sigma-Aldrich公司;乙酸、乙酸钠、无水乙醇等试剂:均为市售国产试剂。
1.2 仪器与设备
SII X-DSC7000型差示扫描量热仪:日本SII NanoTechnology公司产品;D8 ADVANCE广角X射线衍射仪:德国Bruker公司产品;Optima L-80XP型超速离心机:美国贝克曼公司产品;Hyperchem8.0软件:美国Hypercube公司产品。
1.3 实验方法
1.3.1 制备极限糊精及其SE-HPLC分析相对分子质量分布 取100 g小麦淀粉,加蒸馏水配置成5%的淀粉乳。完全糊化后,用0.02 mol/L、pH 5.0的醋酸缓冲液将淀粉糊调节至α-淀粉酶最适作用pH值(pH 6.0),加入α-淀粉酶,然后置于60℃的恒温水槽中酶解24 h后,再将α-极限糊精溶液的pH值调节至β-淀粉酶最适作用范围(pH 5.5),然后加入β-淀粉酶,置于55℃的恒温水槽中酶解48 h,最终将反应液煮沸30 min灭酶活。极限糊精溶液再加入无水乙醇使乙醇浓度分别达到为75%(F75)和88%(F88),进行分级沉淀,采用超速离心的方法分步收集沉淀,并将沉降物冷冻干燥,粉碎,过200目筛。上述制备的极限糊精取0.1 g溶解于5 mL溶液蒸馏水中,将其稀释到100 mL后再在45℃保温2 h。所制备的溶液经0.22 μm的滤膜过滤,取滤液10 μL在 SE-HPLC上使用 Shodex OHpark SB-804 HQ(8.0 mm× 300 mm,i.d.6 μm)柱分析。
1.3.2 WXRD测定淀粉回生 将1 g极限糊精(F75)溶解于20 mL蒸馏水中,再将9 g小麦淀粉加到极限糊精溶液中。用磁力搅拌器搅拌均匀后移入玻璃管中,将其密封后沸水浴糊化1 h,样品放置冷却至室温,放入4℃冰箱保存120 h。极限糊精(F88)的回生样品也按照上述方法制备。采用D8 ADVANCE广角X射线衍射仪对上述所制备的回生样品进行WXRD测定[8]。
1.3.3 DSC测定淀粉回生 分别称量3 mg极限糊精与小麦淀粉的物理混合物 (质量比为5∶95或 10∶90)于铝盒中,用微量进样器添加6 μL蒸馏水后密封,平衡24 h。平衡的样品以10℃/min的速率从20℃升温至90℃,样品冷却至室温,放入4℃冰箱分别保存 24、72、120、360、720 h 后,以 10 ℃/min 的速率从20℃升温至90℃,记录样品回生焓变(ΔH)。测定时以空坩埚为参比,氮气流速为20mL/min。
1.3.4 分子动力学模拟 采用Hyperchem 8.0软件模拟极限糊精与淀粉片段之间的相互作用。根据极限糊精和淀粉片段的分子尺寸设定模拟的边界条件为 40 Å×30 Å×50 Å。 其中淀粉分子片段由 15 个葡萄糖残基以左螺旋的方式连接组成。由于模拟软件的边界参数最大可设置为5 nm,因此淀粉片段的分支点(α,1-6键连接)并未设定在本模拟实验中。所有建立的模型在分子动力学模拟前均使用Amber力场进行构象优化计算。分子动力学模拟DSC测试变温过程:能量优化的模型首先在1 ps内加热90℃后(糊化),并在90℃平衡2 ps,再从90℃降温到4℃(回生)。
2 结果与讨论
2.1 极限糊精的相对分子质量表征
小麦淀粉通过α-淀粉酶和β-淀粉酶复合水解得到极限糊精溶液,采用乙醇分步沉淀的方法得到两个组分(F75和 F88),组分 F75和 F88的 SE-HPLC 见图1。从图1可知,采用乙醇分步沉淀法已将极限糊精分为不同平均相对分子质量大小的极限糊精。极限糊精的保留时间(tR)随乙醇浓度的增大而增大,说明F75组分的相对分子质量较F88大。通过标准相对分子质量校正可知F75和F88的平均相对分子质量分别为 2 861.2(18)和 1 455.4(9)。

图1 SE-HPLC测定极限糊精的高效液相色谱图Fig.1 SE-HPLC chromatographic molecular weight profile of limit dextrin
2.2 X射线衍射分析
X射线衍射图谱见图2。可以看出:所有的样品在2θ角=17°时均呈现出回生淀粉的特征峰(B型衍射峰),其反映的是六角形晶胞β-直链淀粉100个晶面之间的距离[9],亦说明小麦淀粉在糊化后失去其天然存在的A型结晶。此外,在图2中观察到所有的小麦淀粉样品2θ为19.7°都有一个衍射峰,这个衍射峰归属于直链淀粉与脂质形成复合物的V型结晶特征峰[10],该峰的出现说明本实验中淀粉样品中残存脂质。小麦淀粉中加入10%的极限糊精,回生样品的相对结晶度均小于不加极限糊精的小麦淀粉回生的相对结晶度。这个现象可以用Gidley和Bulpin[11]研究成果解释:DP<9的淀粉链片段在浓度为35%和50%的溶液中,4℃的条件下可至少稳定2个月。换言之:小相对分子质量的糊精片段不会发生回生,从而导致极限糊精抑制小麦淀粉回生。

图2 小麦淀粉回生的广角X射线衍射图谱Fig.2 WXRD pattern of the retrograded wheat starch
2.3 DSC分析
融化回生的直链淀粉或直链淀粉与脂复合物需要较高的温度(≥90℃)[12],而在本实验中测定淀粉回生焓值时所关注的温度则是低于90℃的范围。此外,小麦淀粉样品在4℃保存时间长达720 h,所以测定的融化重结晶焓值变化(ΔH)对应的是支链淀粉的回生焓值。糊化的小麦淀粉在4℃存贮24~720 h后,再次使用DSC程序升温扫描样品测试其ΔH,见表1。随着小麦淀粉的回生,回生热流峰出现在55.97℃(±1.28℃),这与前期研究的结果相符[13],在此温度附近的ΔH是由回生的支链淀粉解链所形成。随着存贮时间的延长,回生焓值逐渐增大。与未添加极限糊精的小麦淀粉对照样相比,添加极限糊精的小麦淀粉样回生ΔH显著降低。在相同的条件下,添加F88的小麦淀粉回生ΔH小于添加F75的小麦淀粉回生ΔH。这很可能是由于F88的平均相对分子质量较F75小的缘故,因为分子质量越小的组分在低温下其流动性更强。总之,上述结果说明添加极限糊精可降低小麦淀粉回生程度。Würsch等[14]研究结果表明,DP 6-9的糊精片段能抑制淀粉回生。因此,表1中小麦淀粉回生ΔH的变化说明了极限糊精很可能与小麦淀粉发生相互作用,起到抑制小麦淀粉回生的作用。

表1 小麦淀粉重结晶焓变和Avrami重结晶动力学参数Table 1 Kinetic parameters of wheat starch retrogradation
Avrami方程指数(n)反映的是成核过程的晶体形态,而速率常数(k)反映的是结晶过程的速度,因此Avrami方程(公式1)常用于淀粉回生动力学研究[15]。

从表1可以发现,随小麦淀粉中加入不同种类和比例的极限糊精,小麦淀粉的结晶速率常数k值随之改变。在本研究中,小麦淀粉样品的Avrami指数(n)说明所有的淀粉样品回生的成核模式都为瞬时一维生长。
2.4 分子动力学模拟分析
WXRD和DSC数据表明,F88极限糊精组分抑制小麦淀粉回生的效果较F75极限糊精组分显著,且SE-HPLC显示F88极限糊精组分的平均相对分子质量为1 455.4(9)。因此,选择DP为9的极限糊精与淀粉片段 (DP15)作为分子模拟的对象(图3a为DP 9的极限糊精在真空中的几何优化构象)。 经过升温(2 ps)和降温过程(3 ps)的分子动力学模拟,得到极限糊精与淀粉片段相互作用的最低能量构象(图3b)。此外,回生的淀粉片段在同样的升温和降温模拟后得到(图3c)。图4a是极限糊精与淀粉片段的加水混合体系首先经构象优化,在1 ps中模拟加热到90℃及维持在90℃条件下动力学模拟1 ps,该动力学模拟过程相当于淀粉的糊化过程。而极限糊精与淀粉片段的回生过程则是将糊化的淀粉片段体系进行降温模拟(从90℃降温到4℃),让体系达到一个低能量的动态平衡(见图4b)。由图4b可知降温模拟1 ps后整个极限糊精与淀粉片段加水混合体系达到动态平衡状态(见图3b)。

图3 DP 9极限糊精的几何优化构象(a)、极限糊精与淀粉片段相互作用的分子动力学模拟构象图(b)及回生的淀粉片段模型(c)Fig.3 Geometry optimization conformations of the limit dextrin (a);One of the possible conformations corresponding to interaction of starch fractions with the limit dsextrin(b);c,Conformation corresponding totheretrogradatedstarchfraction(c)
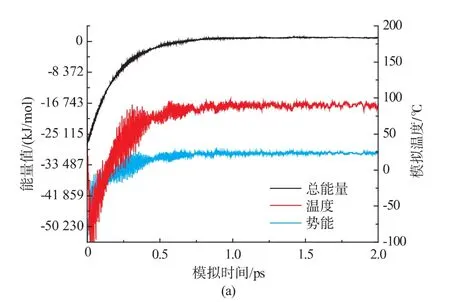
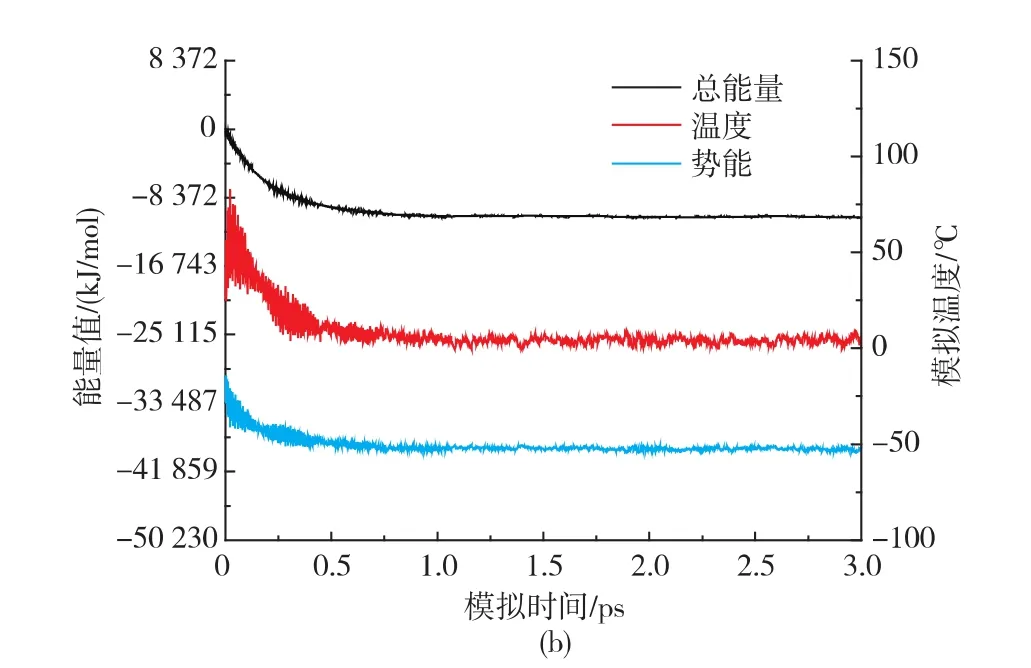
图4 预先优化的构象模拟加热到90℃并在90℃平衡1 ps的总能量、势能和模拟温度的曲线(a)及从90℃到4℃降温模拟3 ps的总能量、势能和模拟温度的曲线(b)Fig.4 Curvesoftotalenergy,potentialenergy,and simulation temperature during the heating simulation performed to 90℃in 1 ps and the simulation at 90 ℃ in 1 ps,based on the preoptimized conformation (a) and Curves of total energy,potential energy,and simulation temperature during the cooling simulation performed from 90 ℃ to 4 ℃ in 3 ps(b)
由表2可知,经分子动力学模拟后,对照淀粉片段与添加极限糊精的淀粉片段相比,对照淀粉片段回生后体积(ΔMV)和表面积(ΔSA)有很大的减小,说明糊化的淀粉片段从无序态转变为有序的双螺旋结构(见图3c)。这个实验结果可以用Tian等[16]研究结果解释,在低温下聚合物会转变成为有序构象。Hyperchem 8.0软件的单点能工具模块计算得到的主要非键合相互作用(氢键能、范德华能和势能)结果见表2(表2中“-”号表示作用力的方向)。其中,氢键能(ΔEHB)的变化最大,据此可判断氢键的相互作用是导致淀粉片段体积和表面积变化的主要作用力,这与Sasaki和Matsuki的报道相一致[17]。从表2中也可看出,范德华作用(ΔEVdW)是导致淀粉片段体积和表面积变化的另外一个重要因素,这是因为范德华作用力是聚合物重结晶的重要驱动力之一[18]。然而势能(ΔEEF)同氢键作用或范德华作用相比,势能变化在淀粉片段结构有序化过程中作用有限。因此,由分子动力学模拟结果可以得出结论:DP 9的极限糊精可以抑制淀粉片段的结构有序化,这与本研究WXRD和DSC的测试结果一致。

表2 淀粉片段的结构性质变化及非键合相互作用参数Table 2 Changes of structure properties and non-bonded interaction energy of starch fractions
3 结语
本研究通过WXRD和DSC技术手段证实极限糊精显著影响小麦淀粉回生的速率和程度。而Avrami方程指数说明小麦淀粉回生成核维数为一维成核。分子动力学模拟结果除印证WXRD和DSC实验结果之外,还说明极限糊精抑制小麦淀粉回生主要通过极限糊精与淀粉片段之间的氢键和范德华作用阻碍淀粉片段的有序化进程。作者旨在了解极限糊精影响淀粉回生,在此基础上为α淀粉酶抑制淀粉回生的机理提供一定的参考依据。
[1]唐敏敏,洪雁,顾正彪,等.黄原胶对大米淀粉长期回生的影响[J].食品与生物技术学报,2013,32(7):692-697.TANG Minmin,HONG Yan,GU Zhengbiao,et al.Effect of xanthan on long-term retrogradation of rice starch[J].Journal of Food Science and Biotechnology,2013,32(7):692-697.(in Chinese)
[2]Miyazaki M,Maeda T,Morita N.Effect of various dextrin substitutions for wheat flour on dough properties and bread qualities[J].Food Research International,2004,37(1):59-65.
[3]Hug-Iten S,Escher F,Conde-Petit B.Structural properties of starch in bread and bread model systems:influence of an antistaling α-amylase[J].Cereal Chemistry,2001,78(4):421-428.
[4]León A E,Durán E,Benedito de Barber C.Utilization of enzyme mixtures to retard bread crumb firming[J].Journal of Agricultural and Food Chemistry,2002,50(6):1416-1419.
[5]Akers A A,R C Hoseney.Water-soluble dDextrins from α-amylase-treated bread and their relationship to bread firming[J].Cereal Chemistry,1994,71(3):223-226.
[6]Witczak M,Korus J,Ziobro R,et al.The effects of maltodextrins on gluten-free dough and quality of bread[J].Journal of Food Engineering,2010,96(2):258-265.
[7]Xu J,Zhao W,Ning Y,et al.Comparative study of spring dextrin impact on amylose retrogradation[J].Journal of Agricultural and Food Chemistry,2012,60(19):4970-4976.
[8]曹旭,王金鹏,陈光耀,等.4-α-糖基转移酶对大米淀粉消化特性的影响[J].食品与生物技术学报,2011,30(2):194-199.CAO Xu,WANG Jinpeng,CHEN Guangyao,et al.Effect of 4-α-glucanotransferase modification on the digestibility of rice starch[J].Journal of Food Science and Biotechnology,2011,30(2):194-199.(in Chinese)
[9]Nishiyama Y,Putaux J l,Montesanti N,et al.B→A allomorphic transition in native starch and amylose spherocrystals monitored by in situ synchrotron X-ray diffraction[J].Biomacromolecules,2010,11(1):76-87.
[10]Lalush I,Bar H,Zakaria I,et al.Utilization of amylose-lipid complexes as molecular nanocapsules for conjugated linoleic acid[J].Biomacromolecules,2005,6(1):121-130.
[11]Gidley M J,Bulpin P V.Crystallisation of malto-oligosaccharides as models of the crystalline forms of starch:minimum chainlength requirement for the formation of double helices[J].Carbohydrate Research,1987,161(2):291-300.
[12]Mestres C,Ribeyre F,Pons B,et al.Sensory texture of cooked rice is rather linked to chemical than to physical characteristics of raw grain[J].Journal of Cereal Science,2011,53(1):81-89.
[13]Ottenhof M A,Hill S E,Farhat I A.Comparative study of the retrogradation of intermediate water content waxy maize,wheat,and potato starches[J].Journal of Agricultural and Food Chemistry,2005,53(3):631-638.
[14]Würsch P,Gumy D.Inhibition of amylopectin retrogradation by partial β-amylolysis[J].Carbohydrate Research,1994,256(1):129-137.
[15]Beck M,Jekle M,Becker T.Starch re-crystallization kinetics as a function of various cations[J].Starch-Starke,2011,63:792-800.
[16]Tian Y,Li Y,Jin Z,et al.A novel molecular simulation method for evaluating the endothermic transition of amylose recrystallite[J].Eropean Fod Rsearch and Tchnology,2009,229(6):853-858.
[17]Sasaki T,Matsuki J.Effect of wheat starch structure on swelling power[J].Cereal Chemistry,1998,75(4):525-529.
[18]Chung H J,Q Liu.Impact of molecular structure of amylopectin and amylose on amylose chain association during cooling[J].Carbohydrate Polymers,2009,77(4):807-815.
猜你喜欢
农业技术与装备(2022年6期)2022-08-17
中国粮油学报(2019年4期)2019-07-12
科学之谜(2018年3期)2018-04-09
新型建筑材料(2018年2期)2018-03-09
食品工业科技(2017年18期)2017-10-16
西安工程大学学报(2016年6期)2017-01-15
中国粮油学报(2016年1期)2016-02-06
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年15期)2014-03-11
