
米蛾卵饲养东亚小花蝽的关键点研究
2014-12-09 09:12杨丽文张志勇
环境昆虫学报 2014年6期
杨丽文,王 甦,张志勇,张 帆*
(1.北京农学院 植物科学技术学院 北京 102206;2.北京市农林科学院植物保护环境保护研究所,北京 100097)
东亚小花蝽Orius sauteri (Poppius)属半翅目Hemiptera 花蝽科Anthocoridae,是农、林小型害虫的优势天敌,其成虫和若虫均可捕食叶螨、蚜虫、粉虱、蓟马等小型节肢动物及多种鳞翅目害虫的卵及初其孵幼虫(Riudavets,1995)。东亚小花蝽主要分布在东亚及俄罗斯的远东地区 (Wang et al.,2002),在该地区的蔬菜、果树、花卉以及农田生态系统中发挥着重要的作用 (Yano,1996)。在日本,东亚小花蝽已经成功用于农田生态系统,如温室中茄子上的南黄蓟马Thrips palmi Karny 等的控制(Kawai,1995;Hemerik and Yano,2010)。目前欧美一些天敌公司已经实现了对美洲小花蝽O.insidiosus (Say),O.laevigatus (Fieber),O.majusculus (Reuter)等小花蝽的商品化生产,并广泛应用于农业生产,尤其对温室蓟马起到了很好的控制效果(Van et al.,1997)。另外,2013年夏季作者在北京温室茄子大棚释放东亚小花蝽,其对二斑叶螨、朱砂叶螨、西花蓟马和粉虱也均表现出良好的控害效果(数据未发表)。因此,东亚小花蝽是一种极具应用价值的杂食性天敌昆虫。
为了满足东亚小花蝽的生物防治需求,多年来国内外相继开展了众多东亚小花蝽的人工饲养应用基础研究,如东亚小花蝽的生物学特性、生长发育、生活规律等。为了实现东亚小花蝽的规模化饲养,科研工作者针对小花蝽的替代饲料进行了大量研究:前人曾利用害螨、蚜虫、蓟马等昆虫和螨类等动物源饲料,以及植物花粉、嫩玉米粒、啤酒酵母液等植物源饲料和微胶囊等人工饲料饲养东亚小花蝽(周伟儒和王韧,1989;Kohno and Kashio,1998),但均具有一定的局限性和消极影响,未能达到规模化饲养的目的。鳞翅目昆虫的卵是东亚小花蝽喜欢取食的饲料之一(Yano,2002)。在欧洲、印度等地科学工作者以鳞翅目昆虫地中海粉斑螟Ephestia kuehniella 卵和印度谷斑螟Plodia interpunctella 卵作为替代饲料成功实现了小花蝽的规模化饲养 (周伟儒和王韧,1989;Cocuzza,1997;Yano et al.,2002;Oliveira et al.,2004;Bonte and De Clercq,2010)。但这两种鳞翅目昆虫非我国原产,并且在我国引种定殖扩繁受环境条件和寄主植物的局限性,很难形成规模。目前国内尚没有成熟的规模化饲养东亚小花蝽的技术手段,尚不能提供批量产品用于田间害虫的控制,无法满足我国设施农业和绿色农业的发展需要。
1 材料与方法
1.1 供试虫源
东亚小花蝽:成虫采集自北京市农林科学院玉米试验田内,后在室内养虫笼(50 cm×50 cm×50 cm)内建立种群。每天供以足量的新鲜米蛾卵为饲料,白芸豆叶片(带完整叶柄,叶柄处包裹湿润的脱脂棉保鲜)为产卵基质。东亚小花蝽发育至成虫期后于叶片上添加10%蜂蜜水作为补充饲料。定期移出已产卵的芸豆叶片,放入上述相同的养虫笼内孵化并按上述方法饲养。饲养三代以上供试。
米蛾:在室内 (温度:25℃-28℃,湿度:60%-80%),利用玉米粉:豆粉:麦麸按重量为7∶2∶1 的比例配置饲料进行人工饲养,幼虫饲养在455 mm×325 mm×40 mm 的养虫盒内,待幼虫羽化后,收集成虫于100 mm×50 mm 的纱网小袋内待其产卵,每天收集新鲜米蛾卵,并制作成10 mm×10 mm 大小的卵卡(每张卡10 粒卵),后紫外灯灭活供试。
1.2 供试器具及试验条件
养虫笼:吉林新天地生物有限责任公司生产;
人工气候培养箱:SANYO,MLR-351H;
东亚小花蝽的饲养条件:T=25±1℃;L∶D=16∶8;RH=75±5 %。除个别说明,该试验均在上述条件下进行。
1.3 试验方法
1.3.1 米蛾卵饲喂东亚小花蝽的相关参数研究
取同一时间段 (5 h 内)东亚小花蝽卵共108 粒,于上述相同设置的培养箱内持续饲养,记录其孵化时间和孵化数量,孵化出的一龄若虫在培养皿(直径9 cm)内单头饲养,培养皿内放置润湿的脱脂棉提供水分,80 目纱网封口,每天提供足量新鲜米蛾卵,每天观察3 次 (6∶ 00,14∶00,22∶00)记录其若虫各龄期的蜕皮时间、羽化时间及存活数量。试验成虫的寿命研究对象为交配过的雌成虫(雌成虫是种群的繁殖的关键因子,且雄虫寿命相对较短)。试验至东亚小花蝽全部死亡。
大陆村依托香米种植优势,打造了全国最新颖的大面积3D稻田艺术,丰富多样的艺术图案吸引了全国各地的游客驻足;大陆村的玉米迷宫、创意石头画、农创市集和乡村美食街等,用绿色植物制造游客项目,以绿色环境打造休憩模式,给游客们带来了新奇的游乐体验;大陆村接连推出的“找稻”新大陆、“欢乐嘉年华”生态旅游主题活动月、稻田文化艺术节等旅游节庆活动,以及“奇趣宾阳”山地自行车赛、2018“活力宾阳”田园迷你铁人三项赛等旅游推介活动,不断提升了古辣镇大陆村的稻田艺术知名度和古辣香米品牌影响力。
1.3.2 东亚小花蝽对米蛾卵的取食量试验
取同一时间段(5 h 内)孵化的东亚小花蝽若虫23 头,分别置于直径2.2 cm 高1.0 cm 的培养皿内单头饲养,并置于同上设置的人工培养箱内,每天每头提供足量(20 粒,约0.75 g)的新鲜米蛾卵卡(10 粒/卡)和润湿的脱脂棉球提供水分,每天定时记录其蜕皮及取食量(显微镜观察:米蛾卵卵壳塌陷干瘪视为被取食)。若虫羽化后雌成虫和雄成虫配对24 h 后分离单头饲养雌成虫,并提供白芸豆具叶柄叶片作为其产卵基质,每天更换产卵基质,每天为雌成虫提供足量米蛾卵20 粒,每天定时记录雌成虫对米蛾卵的取食量。该试验中以雌成虫个体为研究对象(雌成虫是种群的繁殖的关键因子,其取食量直接关系到卵的营养供给是否充足)。
1.3.3 东亚小花蝽最佳饲养密度的研究
取同一时间段(12 h)羽化的东亚小花蝽进行雌雄配对,在直径10 cm 高10 cm 的塑料养虫盒内进行。设置四个饲养密度梯度 5 对/盒,10 对/盒,20 对/盒,30 对/盒。在雌虫产卵期每天定时更换新鲜产卵基质,计数已经产卵的产卵基质上卵的数量,每个处理重复5 次。
1.3.4 产卵基质的筛选
取同一时间段(12 h)羽化未交配的东亚小花蝽雌雄各个体15 头,随机进行雌雄配对,分别在15 个直径10 cm,高10 cm 的饲养盒内成对饲养,80 目纱网封口,置于培养箱内。我们以下列预处理的植物组织作为产卵基质的选择试验对象:白芸豆嫩茎1 个(发芽的芸豆种子主茎长至6 cm左右,截取5 cm 用做产卵基质)、白芸豆叶片1 个(选择真叶,叶柄约5 cm)、发芽的蚕豆种子2 粒(根部约2.5 cm)、夏至草嫩茎1 个(约5 cm),以上供试产卵基质均用湿润的脱脂棉包裹基部保湿,于东亚小花蝽产卵期放入饲养容器内。每天提供足量的新鲜米蛾卵(30 粒/盒),并更换产卵基质;在显微镜下记数已取出产卵基质上的着卵量,然后将其置于相同设置的培养箱内饲养,定期给保湿脱脂棉加水以保证产卵基质的新鲜,记录其孵化率及各产卵基质的保湿特点。试验至成虫停止产卵结束。重复5 次。
1.4 数据统计与分析
东亚小花蝽的产卵量、发育历期、其对米蛾卵的取食量等的平均数均为Mean±SE;不同产卵基质上产卵量的比较及最佳饲养密度筛选试验中不同密度下单雌平均产卵量的比较采用单因素方差分析 (P=0.05),上述数据处理均采用SPSS 18.0进行分析(Allen and Bennett,2010)。
2 结果与分析
2.1 米蛾卵饲喂对东亚小花蝽发育的影响
表1 的参数可以看出,米蛾卵饲养的东亚小花蝽的平均发育历期为28.9 天;种群指数为22.18;幼期累计存活率为64.81%,其中,低龄若虫的存活率低于高龄若虫,存活率从低到高依次 为:1 龄若虫 (82.83%)、2 龄若虫(91.46%)、3 龄若虫 (96%)、4 龄若虫(98.61%)、5 龄若虫(98.59%)。其中1 龄和2龄若虫的存活率较低可能是因为1、2 龄若虫体型微小,行动力弱,容易在培养皿内形成的水膜中溺亡。因此要特别注意低龄若虫饲养的湿度控制,提高若虫成活率。
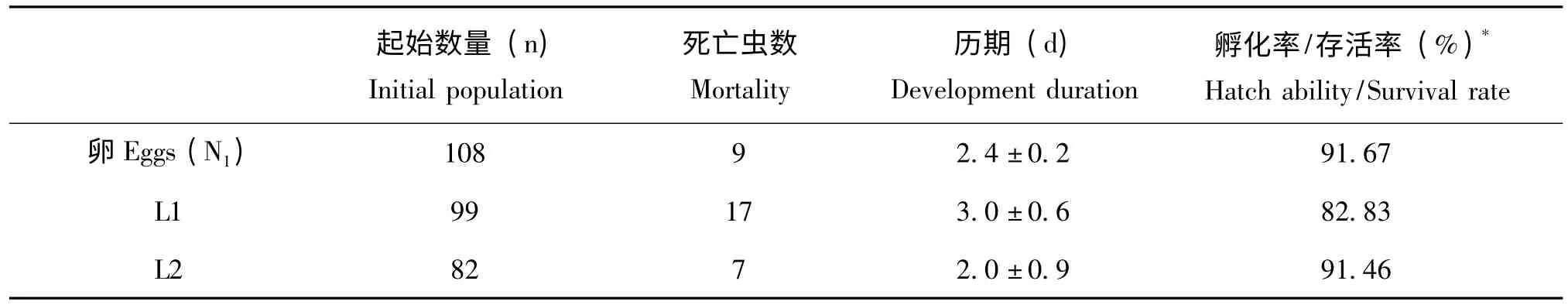
表1 米蛾卵饲喂东亚小花蝽的相关参数Table1 The parameters of Orius sauteri fed on eggs of Corcyra cephalonica
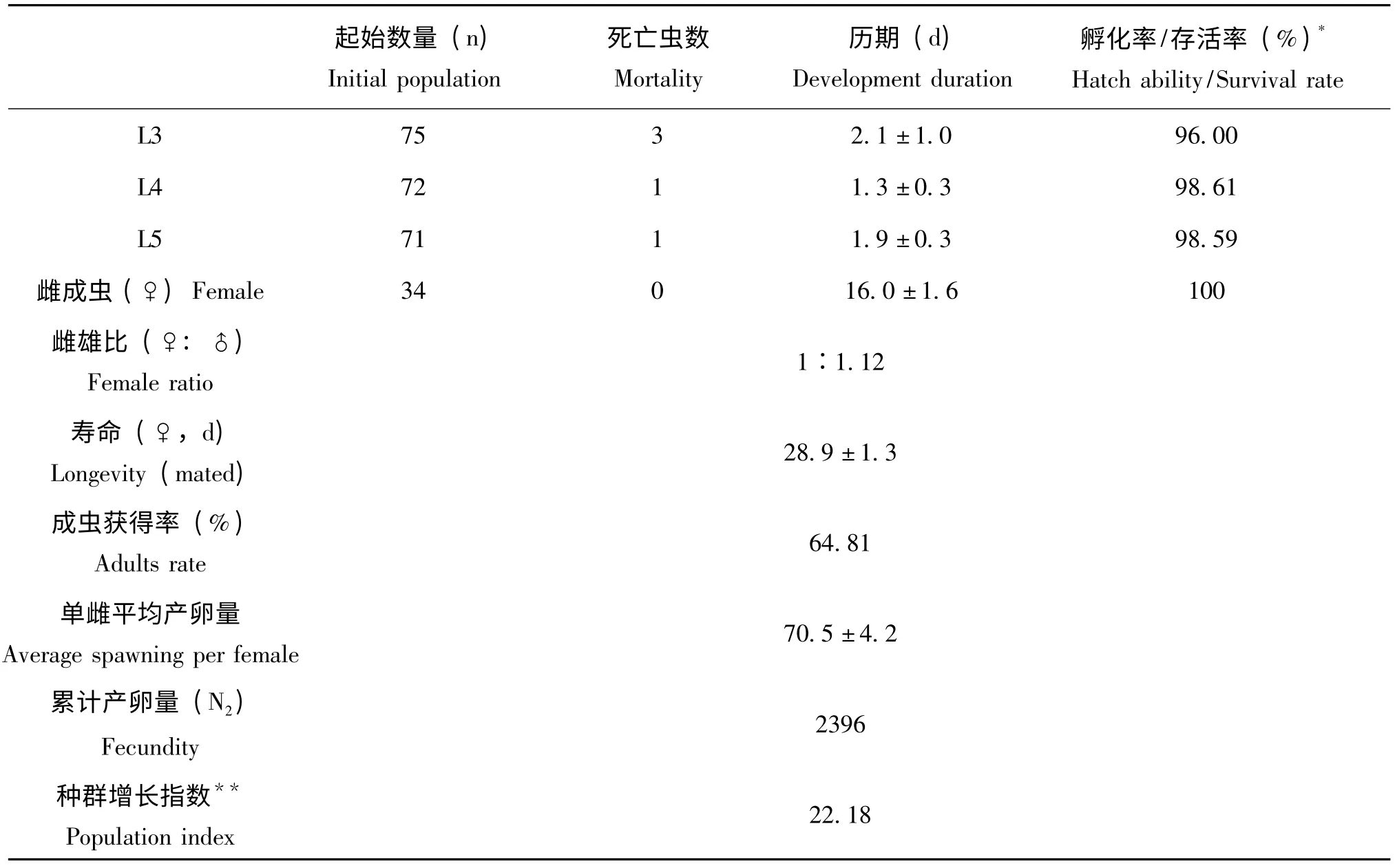
(续上表)
2.2 东亚小花蝽对米蛾卵的取食量
研究发现在食物充足的情况下,雌性东亚小花蝽一生可取食米蛾卵136.9 粒。东亚小花蝽对米蛾卵的取食量随着龄期的增加而增加(表2),其中1 龄和2 龄东亚小花蝽日均取食量相近,分别为2.5 粒/d 和2.7 粒/d;3 龄和4 龄若虫日均取食量分别是5.0 粒/d 和5.9 粒/d;在若虫期,5 龄若虫的日均取食量最多为8.0 粒/d;雌成虫的日均取食量为8.4 粒/d。

表2 东亚小花蝽对米蛾卵的取食量Table 2 The Orius sauteri consumption of Corcyra cephalonica eggs
2.3 东亚小花蝽的饲养密度
表3 结果说明,饲养密度为 5 头/盒、10 头/盒、15 对/盒三个处理中,东亚小花蝽的单雌平均产卵量分别为:68.8 粒、69.7 粒和70.2粒、,三者间没有显著性差异,但是均显著高于20头/盒(52.3 粒)和25 头/盒 (38.4 粒)处理(P<0.05)。
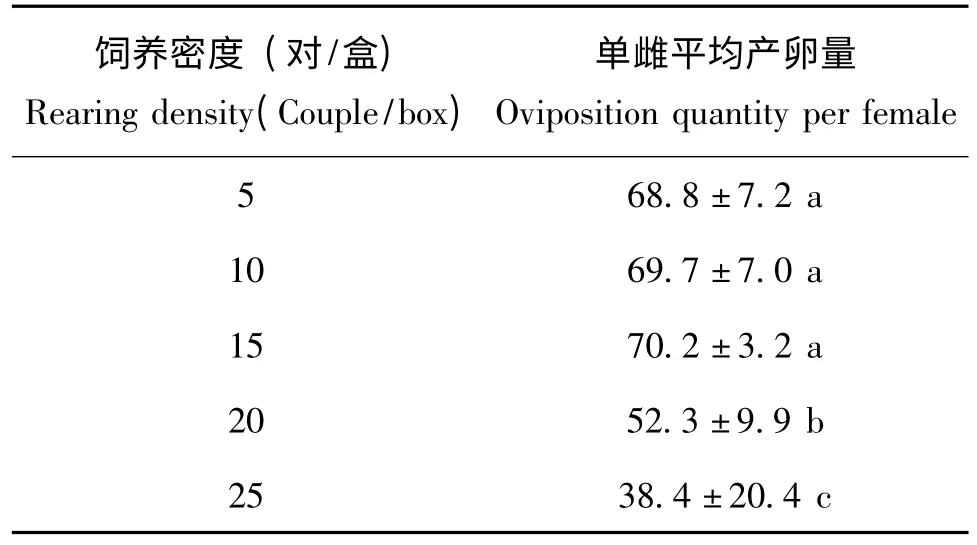
表3 不同饲养密度下东亚小花蝽的单雌平均产卵量Table 3 The average fecundity of Orius sauteri per female in differente rearing density
2.4 产卵基质的筛选
定量和定性的试验观察结果(表4)可见,在供试的四种产卵基质中,白芸豆叶片(具叶柄)因其着卵量显著高于其他三种产卵基质,说明四种产卵基质共存时,东亚小花蝽更喜欢在白芸豆叶片上产卵,且卵的孵化率达98.1%,同时其具有易于保鲜、室内培育方便、可周年供应等优点,因此较其他三种产卵基质,白芸豆具叶柄叶片更适合作为东亚小花蝽规模化饲养的产卵基质。白芸豆嫩茎虽然着卵量不多,但是保湿效果好,卵的孵化率很高,可以和白芸豆叶片混合使用,这样也提高了白芸豆的利用率。试验发现发芽的蚕豆种子其着卵量显著低于白芸豆叶片的着卵量(P<0.05),且由于其保鲜效果不易掌握,造成其孵化率仅为67.3%;同样夏至草叶片保鲜效果不佳,易萎蔫,且仅春夏季可在野外采集到,实验室培育困难,因此后二者不适宜作为规模化饲养东亚小花蝽的产卵基质。

表4 东亚小花蝽产卵基质的筛选Table 4 Optimization of oviposition substrates for Orius sauteri
3 结论与讨论
通过实验研究天敌昆虫规模化饲养的关键技术有助于指导天敌昆虫的规模化饲养。成虫获得率、发育历期、单雌产卵量和雌成虫寿命等均是评价饲养效果的重要指标,它们可以有效的确定天敌的饲养周期和饲养成本。
成虫获得率是评定天敌饲养效率的重要指标之一。周伟儒和王韧(1989)曾利用米蛾卵和水在27℃和RH 70%的条件下饲养东亚小花蝽,其成虫获得率为64.6%,本研究的结果(64.81%)与其相似。成虫获得率的高低取决于其若虫各龄期的存活率的高低,作者在试验中观察发现主要是1 龄和2 龄若虫的死亡率较高,分析原因有二:一是低龄若虫个体微小,容易溺亡在饲养器皿内壁形成的水膜上;二是在饲养过程中由于若虫生长发育的不均一性,也会出现较大个体攻击较小个体致死的现象。因此,增加东亚小花蝽低龄幼虫的成活率对于提高人工饲养繁殖效率具有至关重要的作用。为了增加东亚小花蝽各龄期的存活率,保证其生长的适宜环境是必要的。温度和湿度又是影响昆虫生长发育速率的重要环境因素。于毅等(1998)的研究发现东亚小花蝽在20-28℃的范围内,随着温度的升高其发育速率加快,发育历期缩短。本研究中设置温度为25± 1℃,RH 75±5%,东亚小花蝽若虫发育历期为12.9 d,略长于周伟儒和王韧(1989)在27℃,RH70%的条件下同样以米蛾卵饲喂的结果(10.08 d),与前人研究结果一致。同样,不同饲料处理对于小花蝽的生长发育也有一定的影响。本研究结果同Jeffrey 等 (1998)利用地中海粉斑螟Ephestia kuehniella 卵饲养的东亚小花蝽的结果(12.0 d)相近,却短于Yano 等(2002)14.0 d 的发育历期;Jeffrey 等(1998)供试的产卵基质与本研究相同均为采用白芸豆叶片,而Yano 等(2002)则利用发芽的蚕豆种子作为产卵基质。本研究中的雌成虫寿命 (16.0 d)略长于Yano 等 (2002)(13.7 d)和Jeffrey 等(1998) (13.9 d)的研究结果。基于以上结果分析,东亚小花蝽的生长发育速率和发育历期可能受温度、湿度、饲料、产卵基质等多种因素共同作用的影响,因此在东亚小花蝽的规模化饲养研究中,综合考虑多种因素的影响有助于筛选出适合繁殖东亚小花蝽的最佳条件。
东亚小花蝽的产卵量同样受多种因素的影响。Yano (2002)等的研究报道,东亚小花蝽的单雌平均产卵量为103.9 粒,远高于本研究结果的70.47 粒与Jeffrey 等(1998)的68.4 粒;东亚小花蝽的产卵量在本研究中利用米蛾卵和Jeffrey 等(1998)利用地中海粉斑螟卵两种不同替代寄主间差异不大,但均明显低于Yano 等(2002)研究结果。是供试的昆虫种群不同、产卵基质影响、抑或是试验条件差异而致?仍需要进一步研究和分析。
在天敌人工大量饲养中,精准掌握其不同龄期的日取食量是非常重要的。对于利用米蛾卵作为替代寄主的东亚小花蝽的取食量研究尚未见报道。本研究的各龄期取食量及其历期等结果可用于指导制定东亚小花蝽及其寄主的繁殖计划,精准控制米蛾卵的日供给量,可有效避免饲料的浪费,降低饲养成本。
在长期的进化过程中,昆虫为了获得适宜的生长发育及繁殖条件,存在种内竞争,昆虫种内斗争是其对外界压力的一种适应性表现(Klomp 1964)。在天敌昆虫(特别是捕食性天敌)大量人工繁殖中,饲养的空间密度是影响繁殖效率的重要因素之一,也是控制产量和质量的关键环节。研究发现动物间的自相残杀多由于食物不足、个体种群的密度变化及龄期变化等因素 (Fox,1975);宋丽文等(2010)的研究发现不同饲养密度对蠋蝽Arma chinensis 若虫各龄期发育历期的影响无规律,不同饲养密度对蠋蝽生殖力有不同程度的影响,密度过高或过低都显著降低其生殖力。但是,关于东亚小花蝽的饲养密度的研究尚未见相关报道。作者在研究中观察到东亚小花蝽个体间存在自残现象。本研究结果表明每对50 cm3的密度饲养东亚小花蝽成虫时,其种间个体对空间和食物的竞争最小,其单雌产卵量也最高。这对于确定规模化饲养密度,提高空间和食物的利用率,降低饲养成本具有重要意义。因此,确定东亚小花蝽的最佳饲养密度也是解决规模化饲养东亚小花蝽的关键点之一。
小花蝽具有将卵产于植物组织中的习性,因此产卵基质的筛选也是规模化饲养东亚小花蝽的关键点之一。对此国内外工作者已经做了很多相关研究,例如Richards 和Schmidt (1996)利用四季豆Phaseolus vulgaris L.茎、四季豆荚、土豆嫩芽作为美洲小花蝽的产卵基质,研究发现美洲小花蝽更偏好在四季豆茎上产卵;在相对湿度为100%的条件下,美洲小花蝽卵Oriush insidiosus 在四季豆荚上卵的孵化率最高为59%,其显著高于相同条件下土豆嫩芽 (39%)和四季豆茎(29%)。研究发现四季豆荚在高湿状态下容易腐烂变质,降低了小花蝽卵的孵化率(Richards and Schmidt,1996)。周伟儒等(2001)利用黄豆芽、寿星花嫩茎及燕子掌叶片作为东亚小花蝽的产卵基质的研究时发现黄豆芽易萎蔫干枯,不能保证小花蝽卵和低龄幼虫对植物源饲料的需求,且干枯黄豆芽不便于清理;燕子掌叶片和寿星花上小花蝽卵的孵化率分别为70%和75%,本研究中利用白芸豆具叶柄叶片上东亚小花蝽卵的孵化率可高达91.67%,而具叶柄的白芸豆叶片的保湿时间可持续一周以上,完全可以保证东亚小花蝽卵的孵化和幼虫对水分及植物源食物的需求,可作为东亚小花蝽规模化生产的适宜产卵基质。
一直以来,食品安全和生态安全备受关注,对害虫生物防治技术和产品的需求也日趋强烈。东亚小花蝽作为一种优质的捕食性天敌昆虫,其产品具有广阔的应用前景。但其规模化繁殖技术仍需进一步研发和完善,以期在我国害虫的生物防治中发挥重要作用。
References)
Allen PJ,Bennett K.PASW Statistics by SPSS:A Practical Guide:Version 18.0 [M].Cengage Learning South Melbourne,Australia.2010.
Bonte M,De Clercq P.Influence of diet on the predation rate of Orius laevigatus on Frankliniella occidentalis [J].BioControl,2010,55 (5):625-629.
Cocuzza G,Clercq P,Veire M,et al.Reproduction of Orius laevigatus and Orius albidipennis on pollen and Ephestia kuehniella eggs[J].Entomologia Experimentalis et Applicata,1997,82 (1):101-104.
Du XG,Yan YH.Effect of the cover crops on the pest mite Panonychus ulmi and its predator Orius sauteri [J].Chinese Journal of Biological Control,1994,10 (3):114-117.[杜相革,严毓骅.苹果园混合覆盖植物对害螨和东亚小花蝽的影响[J].生物防治通报,1994,10 (3):114-117]
Fox LR.Cannibalism in natural populations[J].Annual Review of Ecology and Systematics,1975,87-106.
Hemerik L,Yano E.A simulation model for the functional response of Orius sauteri on eggplant leaves with Thrips palmi:implications for biological control[J].Proc.Neth.Entomol.Soc.Meet,2010,21:61-74.
Honda JY,Nakashima Y,Hirose Y.Development,reproduction and longevity of Orius minutus and Orius sauteri (Heteroptera:Anthocoridae) when reared on Ephestia kuehniella eggs [J].Applied Entomology and Zoology,1998,33 (3):449-453.
Kawai A.Control of Thrips palmi Karny (Thysanoptera:Thripidae)by Orius spp.(Heteroptera:Anthocoridae)on greenhouse eggplant[J].Applied Entomology and Zoology,1995,30:1-7.
Kohno K,Kashio T.Development and prey consumption of Orius sauteri(Poppius)and O.minutus (L.) (Heteroptera:Anthocoridae)fed on Frankliniella occidentalis (Pergande)(Thysanoptera:Thripidae)[J].Applied Entomology and Zoology,1998,33:227-230.
Lattin JD.Bionomics of the Anthocoridae [J].Annual Review of Entomology,1999,44 (1):207-231.
Oliveira H,De Clercq P,Zanuncio,et al.Nymphal development and feeding preference of Podisus maculiventris (Heteroptera:Pentatomidae) on eggs of Ephestia kuehniella (Lepidoptera:Pyralidae) parasitised or not by Trichogramma brassicae(Hymenoptera:Trichogrammatidae)[J].Brazilian Journal of Biology,2004,64 (3A):459-463.
Riudavets J.Predators of Frankliniella occidentalis (Perg.)and Thrips tabaci Lind.:a review [J].Wageningen Agricultural University Papers,1995,95 (1):43-87.
Richards P,Schmidt J.The suitability of some natural and artificial substrates as oviposition sites for the insidious flower bug,Oriusinsidiosus[J].Entomologia Experimentalis et Applicata,1996,80(2):325-333.
Song LW,Tao WQ,Guan L,et al.Influence of host plants and rearing density on growth,development and fecundity of Arma chinensis[J].Scientia Silvae Sinicae,2010,(46)3:105-110 [宋丽文,陶万强,关玲,等.不同宿主植物和饲养密度对蠋蝽生长发育和生殖力的影响[J].林业科学,2010,46 (3):105-110]
Van Lenteren JC,Roskam MM,Timmer R.Commercial mass production and pricing of organisms for biological control of pests in Europe[J].Biological Control,1997,10 (2):143-149.
Wang YH,Wu H,Bu WJ,et al.Geographic distribution of the genus Orius Wolff (Heteroptera:Anthocoridae)[J].Journal of Zhejiang Forestry College,2002,20 (4):389-393.
Wu KJ,Chen YP,Li MH.Life tables for experimental populations of the cotton bollworm,Heliothis Armigera (Hubner),at different temperatures[J].Acta Entomologica Sinica,1978,21 (4):385-392.[吴坤君,陈玉平,李明辉,不同温度下的棉铃虫试验种群生命表[J].昆虫学报,1978,21 (4):385-392]
Xu XN,Wang ND.Techniques for production and application of natural enemies in abroad [J].Chinese Journal of Biological Control,2008,24 (1):24-29.[徐学农,王恩东.国外昆虫天敌商品化生产技术及应用[J].中国生物防治,2008,24 (1):24-29]
Yano E.Biology of Orius sauteri (Poppius)and its potential as a biocontrol agent for Thrips palmi[J].IOBC/wprs Bull.,1996,19:203-206.
Yano E,Watanabe K,Yara K.Life history parameters of Orius sauteri(Poppius) (Het.,Anthocoridae)reared on Ephestia kuehniella eggs and the minimum amount of the diet for rearing individuals[J].Journal of Applied Entomology,2002,126 (7-8):389-394.
Yu Y,Yan YH,Hu XS.Effect of nutrition and other ecological factors on the development of Orius sauteri [J].Chinese Journal of Biological Control,1998,14 (1):4-6.[于毅,严毓骅,胡想顺.营养和生态因子对东亚小花蝽生长发育的影响[J].中国生物防治,1998,14 (1):4-6]
Yu Y,Zhang AS,Yan YH.Studies on the occurrence and dispersion of Orius sauteri in relation to ground cover vegetation in apple orchard and adjacent farms[J].Chinese Journal of Biological Control,1998,14 (4):148-151.[于毅,张安盛,严毓骅.东亚小花蝽的发生和扩散与苹果园和邻近农田植被的关系[J].中国生物防治,1998,14 (4):148-151]
Zhou WR,Wang R.Rearing of Orius sauteri (Hem.:Anthocoridae)with natural and artificial diets[J].Chinese Journal of Biological Control,1989,5 (1):9-12.[周伟儒,王韧.用天然和人工饲料饲养小花蝽的研究[J].中国生物防治,1989,5 (1):9-12]
Zhou WR,Wang R,Qiu SB.Use of soybean as the oviposition material in mass rearing of Orius sauteri (Het:Anthocoridae)[J].Chinese Journal of Biological Control,1991,7 (1):7-9.[周伟儒,王韧,邱世邦.用黄豆芽作产卵植物繁殖东亚小花蝽[J].中国生物防治通报,1991,7 (1):7-9]
猜你喜欢
农业工程学报(2022年10期)2022-08-22
天津农业科学(2022年5期)2022-05-31
小学生学习指导(当代教科研)(2021年6期)2021-05-23
黄河之声(2021年2期)2021-03-29
学生天地(2020年11期)2020-08-25
外国问题研究(2018年2期)2018-07-04
焊接(2017年11期)2018-01-04
初中生世界·七年级(2017年7期)2017-09-04
教学与研究(2016年1期)2016-02-28
长江大学学报(自科版)(2015年21期)2015-12-04
