
红雉凤仙花结籽的花粉限制
2014-12-02 04:17李艳红
四川林业科技 2014年4期
李艳红,王 琼
(西华师范大学生命科学学院,四川南充 637000)
生殖力是种群兴衰的主要标志(王亚琴等,2007)。在自然状态下,开花植物的生殖力常常受到花粉限制,Knight认为,“当植物产生的果实或者种子比接受到足够的花粉少得多的现象说明该植物受到花粉限制”(Knight et al.,2005);Cruden也注意到,异交植物的花粉/胚珠比率(P/O比)远远大于自交者,他认为P/O比是由“最小花粉-胚珠比率”所决定的:低于这个最小比率,种子生产就会受到花粉量制约(张大勇等,2001)。因此,花粉限制常常被理解为花柱上接受到的花粉粒数量不足(数量限制)。但是,花粉限制又常常与质量联系在一起(Herlihy et al.,2004),不同来源的花粉其质量各异,萌发率和花粉管生长情况也不同——低质量的花粉可能减少种子的产量和质量(Charlesworth et al.,1987)。自交不亲和物种的自花花粉受精会降低异花花粉受精的成功率(Ramsey et al.,2000;Kawagoe et al.,2005),因为自花的花粉管可能使胚珠在子房中失活,或者通过自交的不亲和性使得植株根本无法受精(Sage et al.,1994);对于自交亲和的物种,当柱头上存在自交花粉时,常常导致激烈的花粉竞争:子房可能对自交花粉管具有抑制作用、自交胚珠可能更容易败育等(黄双全等,1998),这种由于自交引起的近交衰退对后代适合度的影响体现在植物生活史的各个阶段,如自交的种子产生的后代在发芽、存活、生长等阶段都不及杂交后代(何亚平等,2003)。在植物的有性生殖过程中,无论是由于传粉媒介的作用还是因为自交,花朵都可能接收到自身花粉,从而导致这样的质量限制经常发生。另外,从较远植株上或其它物种接收的远交亲和的花粉又可能因为远交衰退而降低种子产量(Waser et al.,1992)。因此,这种由于不同来源的花粉所造成的花粉质量限制是影响植物有性生殖成功的又一重要因素。
花粉限制常常导致植物较低的结籽率。Husband和Schemske通过实验发现,与异花授粉相比,自花授粉的62个自交亲和物种其种子产量平均减少了20%,最大的减少了近87%(Lloyd et al.,1992;Husband et al.,1996),这说明花粉限制严重降低了植物的有性生殖成功。为了缓解这种现象,植物常常采取了相应的策略来避免或减轻花粉限制,如增大花部展示(花朵大小、花冠颜色和气味等)或者提供相应的报酬(花粉、花蜜等)来吸引传粉者以增加授粉几率(张大勇,2004),同时植物还通过对花部结构的精心设置,如花柱异长、雌雄蕊异位或雌雄异熟等在一定程度上都有效地降低了自交几率(张红玉,2005),从而提高后代适合度。因此,花粉限制还与传粉者的传粉效率有关。
结籽率是衡量开花植物生殖能力强弱的重要标准之一。生长在峨眉山常绿阔叶林下的红雉凤仙花是一种1 a生草本,其植株秀丽、花姿优美,是潜在的优秀花卉资源之一。但是,自2010年以来野生分布的种群数量锐减,且自然状态下结籽率不到50%。对红雉凤仙花主要传粉昆虫的传粉效率进行检测,以分析红雉凤仙花结籽是否受到花粉数量限制;进行人工补充授粉,以检测红雉凤仙花结籽是否受花粉来源限制。总之,关于红雉凤仙花生殖生态学的研究,有助于找出其数量锐减的原因,阐明生殖受限的机理,为制定相应的保护措施提供理论依据。
1 材料与方法
1.1 研究地概况
峨眉山位于四川盆地西南边缘(E103°15'~103°28',N29°31'~29°38'),动植物资源极其丰富,其复杂的地形和地理环境造成了整个山区云雾多、日照少、雨量充沛的气候特点;山麓气候湿润,平均气温17.2℃,最冷月 (1月)平均气温7℃ ,最热月(7月)平均温度 26.3℃;年均降雨量为1 555.3 mm,其中5月~9月份的降雨量占全年的70%~80%(朱晓帆等,1997)。实验样地位于峨眉山万年寺附近海拔约1 000 m的黑水村,近年来随着全球气候变暖,这里的生态环境发生了巨大变化:山涧小溪水位降低,动植物数量锐减,虫害严重,给当地的生产生活带来极大影响,同时也对生长在这一带的珍稀植物造成严重威胁。
1.2 研究方法
由于花冠形态常常与传粉效率有关,在盛花期随机挑选30株生长健康的红雉凤仙花植株,每一株随机选择3朵生长健康的花测定其花冠形态指标;同时统计所选每朵花的胚珠数量。各形态指标标准如下(图1):
花冠深度(FL):从唇瓣上最后一条纹到花冠口的长度;
花冠宽(FW):即花冠口两个翼瓣小裂片之间的距离;
翼瓣长(VL):花冠口到翼瓣尖端的距离;
花冠总长(FL+SL):花冠深度与距长之和;
距长(SL):从唇瓣最后一条纹到距末端的距离;
距圈直径(SCD):蜜距弯曲一圈的直径;
距弯曲度:即红雉凤仙花距的弯曲程度,参照王琼的方法(王琼等,2013);
有蜜距长:当三条熊蜂访过之后,花朵含有花蜜的距的长度。

图1 花冠形态指标(王琼等,2013)Fig.1 The standard of morphological index in the flower
同时测定红雉凤仙花主要传粉昆虫—三条熊蜂(Bombus trifasciatus Smith)的传粉效率,另外选择30株生长健康的植株,分别测量单只三条熊蜂单次访花移出的花粉数量和落置在柱头上的花粉数量。
重新选择30株生长健康的植株,进行人工补充授粉实验,取相邻近似大小的植株(30株)为参照,果实即将成熟时即采回统计结籽率。
1.3 数据分析
所有数据采用 Excel进行输入和预处理;用Spss17.0软件计算平均值,用Spearman进行相关性分析,用One-way ANOVA进行单因素方差分析,用Duncan(D)进行多重比较。
2 结果分析
2.1 花冠形态与传粉效率
通过对花粉的运动去向进行统计,发现红雉凤仙花的主要传粉者—三条熊蜂单次访花时带出的花粉数量远远大于带到柱头上的花粉数,而落置到柱头上的花粉数又远远大于子房内的胚珠数(表1),说明红雉凤仙花的传粉昆虫三条熊蜂的传粉效率较高,其柱头上落置的花粉数量充足。

表1 三条熊蜂访花时移出和花粉落置的花粉数(平均数±标准差)
检测花冠形态与移出花粉数量的相关性,发现三条熊蜂单次访花时带出的花粉数量与花冠的距长、距弯曲度、有蜜距长存在一定的负相关,与距圈直径存在一定正相关,但相关性均不显著(表2)。

表2 花冠形态与移出花粉数的关系
分析花冠形态与落置花粉数量的相关性,发现三条熊蜂单次访花时带到柱头的花粉数量与花冠的距弯曲度、距圈直径、有蜜距长存在一定负相关,与距长存在一定正相关,但相关性也不显著(表3)。

表3 花冠形态与落置花粉数的关系
2.3 传粉效率与结籽率
同时,分别检测三条熊蜂单次访花移出和落置的花粉数与结籽率的相关性,发现红雉凤仙花结籽率与移出和落置的花粉数量相关性均不显著(表4)。

表4 落置花粉数与结籽率
2.4 人工补充授粉对结籽率的影响
在同一植株和不同植株进行比较,发现不同授粉方式对红雉凤仙花的结籽率造成不同影响{图2,(F=1.029,Sig.=0.315)}。与自然传粉相比,异株异花授粉的结籽率提高了约10%,而同株异花授粉与对照之间差异不显著。说明花粉来源对红雉凤仙花结籽率确实存在一定影响。
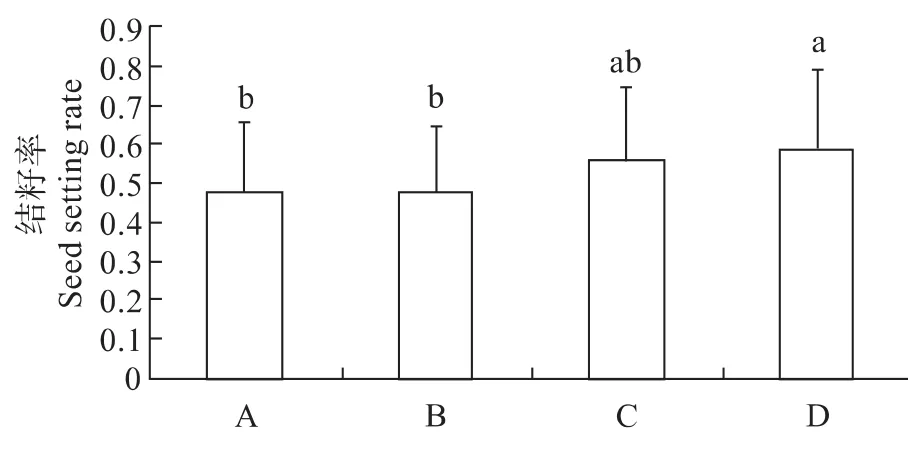
图2 人工授粉的结籽率Fig.2 The seed setting rate of supplemental pollination
3 讨论
开花植物有性生殖的成功很大程度上依赖于为其授粉的传粉者的花粉转移效率,其中行动灵巧的昆虫类占有举足轻重的地位。除了花朵的颜色、气味、大小等对传粉者具有吸引作用的因素之外(唐璐璐等,2007),花冠的形态、报酬物等可能通过影响传粉者传粉效率而影响植物的生殖成功。不同的植物常常拥有不同形态的花朵,针对形态各异的花朵昆虫的访花行为和访花效率也不尽相同,Young通过研究传粉者对I.capensis距形态的选择,发现蜜距的形态对I.capensis雌性生殖成功和雄性生殖成功均有显著影响:当蜜距弯曲度较大时,熊蜂的访花时间也较长,其相应的结籽率也相对较大;同时,与蜜距弯曲度较小的花朵相比,弯曲度更大的蜜距内,熊蜂常常留下更少的花蜜(Young,2008),因此,蜜距的形态可能通过影响传粉者的访花行为间接影响访花效率——传粉者在拥有更多报酬物的花朵上可能停留时间更长,从而使柱头授粉更充分。通过分析红雉凤仙花主要传粉者的访花效率与花冠形态的关系,发现三条熊蜂(Bombus trifasciatus Smith)单次访花时移出和落置的花粉数与花冠的距长、距弯曲度、距圈直径以及有蜜距长均不存在显著的相关性,说明三条熊蜂访花时对花冠形态特征选择性不明显,这可能是因为它更多地倾向于对其他因素的选择,如花的颜色、花蜜的体积或含糖量的高低等。由于花蜜可以对传粉者访花过程中消耗的能量进行补充,因此拥有较多花蜜的花朵可能更容易受到传粉者的青睐(王洁,2012)。传粉者在选择花朵时常常避免访问刚刚被其他传粉者访问过的花,对于刚被访过的花,传粉者仅凭留在花冠上的其它传粉者留下的气味即能辨别(Makoto,1998)。进行传粉效率实验时发现三条熊蜂喜爱访问雄花期的花,因为雄花期花朵除了能够提供丰富的花蜜之外,还有许多花粉可供搜集,而雌花期花朵则仅有花蜜;另外,除了报酬本身多少之外,处于雌花期的花花瓣颜色明显变淡,熊蜂从颜色的艳丽程度也能辨别哪些是具有更丰富报酬的花朵。对于刚被访问过的花,三条熊蜂仅在花冠口停留一下,随即退出花冠开始寻找下一个目标,因此,除了花冠本身的资源,访花者的访花习性及它们之间的相互作用也可能影响传粉效率。
作为中高海拔地区一种主要的传粉昆虫,三条熊蜂具有较强的热调节能力,能在逆境中飞行,花粉传递效率高等特点(何亚平等,2005),这些均使得三条熊蜂具有较强的适应恶劣环境的能力,特别是在应对湿度较大的阴雨天气时比其他昆虫更胜一筹。作为一种自交亲和的植物,红雉凤仙花选择了三条熊蜂为其传粉,在一定程度上解决了高山环境中气候多变,生态因子不稳定,温度低且昼夜波动大等不利因素(Young et al.,2007)。通过对访花昆虫进行观察,发现红雉凤仙花的访花者有熊蜂、蝴蝶和摇蚊等,其中最有效的传粉者为三条熊蜂。与其他传粉蜂类相比,观察到熊蜂的主要特点是个体大,携粉多,但没有专门装载花粉的器官,到其将搜集到的花粉主要以花粉团的形式粘附在后腿上,飞行时势必造成花粉的大量浪费,因此,这是否是导致红雉凤仙花的传粉受限的原因呢?在盛花期,我们通过对熊蜂的访花效率进行评估,发现熊蜂具有较高的花粉移出和落置率,特别是带到柱头上的花粉数量远远大于子房内的胚珠数,因此,红雉凤仙花在开花高峰期不存在花粉数量的限制,这可能是因为盛花期种群的开花样式较大,对传粉者的吸引相对增加(唐璐璐等,2007;肖宜安等,2004),从而提高了熊蜂数量和访花频率。但是,通过分析熊蜂移出花粉数量及落置花粉数量分别对结籽的效应发现,红雉凤仙花的结籽率与熊蜂单次访花时移出和落置的花粉数相关性不明显,因此,只能说明红雉凤仙花较低的结籽率可能与落置在柱头上的花粉质量有关。
不同来源的花粉对结籽的效应不同(施祥等,2010)。一般,异交可以提高后代的适合度,从而增加种群的竞争力;而自交则可以通过降低植物的生殖成功最终导致整个种群平均适合度的下降(郭华等,2003;杨琨,2012)。相应的,为了避免自交,植物往往发展了相应的策略,如两性花雌雄功能在空间或时间上的分离——雌雄异位或雌雄异熟,被广泛地认为能部分有效地阻止自花传粉(任明讯等,2012)。但是,这却不可避免花朵之间的同株异花传粉,同株异花传粉本身不是一种适应,而是植物为了提高异交花粉输出量而产生的一个无法避免的后果(张大勇等,2001),然而,这对种群的生殖成功却可能产生完全不一样的后果。通过观察发现,红雉凤仙花具有从基部到顶部的开花顺序,这使得其主要访问者三条熊蜂访花的访花顺序通常也是从基部到顶部;又由于红雉凤仙花雄性先熟的特点,使得三条熊蜂从基部到顶部访花完毕后离开时带走的正好是植株上部处于雄花期的花粉(张大勇,2004)。在盛花期,当一个植株上有较多花时,最下部的花已经通过了雄性阶段而正处于雌性阶段,三条熊蜂来访时携带的外来花粉正好可以给它们授粉。因此,红雉凤仙花实际上只有很少部分花朵接收到了其它植株的花粉,而绝大部分种子的产生均源于同株异花授粉,即自交。自交花粉和异交花粉常常存在着竞争,如子房对自交花粉管的抑制、自交胚珠的败育等都会对种子的产生造成影响(Husband et al,1996;Mitchell et al,1993)。通过人工补充授粉,发现同株异花授粉植株的结籽率明显高于对照组,说明在自然传粉状况下,落置在红雉凤仙花柱头上的花粉数量虽多,但可能只有很少一部分萌发或穿过柱头到达子房与胚珠结合进而发育成种子。因此,花粉来源是导致红雉凤仙花结子率低的重要原因之一。
峨眉山上夏秋季节降雨量充沛,这是红雉凤仙花短期内迅速生长的根本保证,但是大量的雨水对其有性生殖造成较大影响,特别是花期与雨季的重叠可能限制昆虫的种类及活动能力(杨期和等,2011)。一般情况下,1 a生草本植物一年仅有1次开花结实的机会,为了顺利完成有性繁殖,红雉凤仙花选择了自交亲和的交配系统,允许花朵通过同株异花授粉来增加种子的产量,以对异交繁殖风险进行补充。同时,红雉凤仙花最有效的传粉者—三条熊蜂虽然具有较高的访花效率,但是其较少的数量以及较低的访花频次也不得不使红雉凤仙花选择自交亲和的繁育系统来保证每个子房都能有胚珠成功受精,从而提高生殖成功的几率。因此,较大比例的同株异花授粉可能是红雉凤仙花选择有性生殖成功的一种保障机制。
本实验表明,盛花期红雉凤仙花大量自交导致的花粉质量限制而不是数量限制可能是其结籽率低的主要原因之一。但是,在其开花初期和后期,由于花朵数量稀少导致传粉诱物不足而引起的传粉者数量较少,可能使红雉凤仙花存在不同程度的数量限制。除了花粉限制,红雉凤仙花的结籽可能还受到其他因素的影响,比如资源限制等。
[1]王亚琴,李菁,陈军.贵州梵净山尖叶拟船叶藓生物量的生殖配置[J].浙江林学院学报,2007,24(2):145~149.
[2]Knight T M,Steets J A,Vamosi J C,et al.Pollen limitation of plant reproduction:Pattern and process[J].Annual Review of Ecology Evolution Systematics,2005,36:467~497.
[3]张大勇,姜新华.植物交配系统的进化、资源分配对策与遗传多样性[J].植物生态学报,2001,25(2):130~143.
[4]Herlihy C R,Eckert C G.Experimental dissection of inbreeding and its adaptive significance in a flowering plant,Aquilegia Canadensis(Ranunculaceae)[J].Evolution,2004,58(26):93~2703.
[5]Charlesworth B,Charlesworth D.Inbreeding depression and its evolutionary consequences[J].Annual Review of Ecology Evolution and Systematics,1987,18:237~268.
[6]Ramsey M.,Vaughton G.Pollen quality limits seed set in Burchardia umbellate(Colchicaceae)[J].American Journal of Botany,2000,87:845~852.
[7]Kawagoe T,Suzuki N.Self-pollen on a stigma interferes with outcrossed seed production in a self-incompatible monoecious plant,Akebia quinata(LardizabaIaceae)[J].Functional Ecology,2005,19:49~54.
[8]Sage T L,Berlin R I,Williams E G.Ovarian and other late-acting self-incompatability systems[J].1994.Pages 116~140 in Williams E G,Clarke A E,Knox R B,editors.Genetic control of selfincompatability and reproductive development in flowering plants[C].Kluwer Academic Publishers,Dordrecht,The Netherlands.
[9]黄双全,郭友好,陈家宽.渐危植物鹅掌楸的授粉率及花粉管生长[J].植物分类学报,1998,36(4):310~316.
[10]何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151~163.
[11]Waser N M,Price M V.Outcrossing distance effects in Delphinium nelsonii:pollen loads,pollen tubes,and seed set[J].Ecology,1991,72:171~179.
[12]Lloyd D G,Schoen D J.Self-and crossfertilization in plants.I.Functional dimensions[J].International Journal of Plant Sciences,1992,153:358~369.
[13]Husband B C,Schemske D W.Evolution of the magnitude and timing of inbreeding depression in plants[J].Evolution,1996,50:54~70.
[14]张大勇.植物生活史进化与繁殖生态学[M].北京:科学出版社,2004,97~159.
[15]张红玉.虫媒植物与传粉昆虫的协同进化(二)—虫媒花的性状对昆虫传粉的适应[J].四川林业科技,2005,26(6):22~27.
[16]朱晓帆,蒋文举,朱联锡等.峨眉山环境现状研究[J].四川环境,1997,16(2):9~17.
[17]Wang Q,Li Y X,Pu X Z et al.Pollinators and nectar robbers cause directional selection for large spur circle in Impatiens oxyanthera(Balsaminaceae)[J].Plant Systematics and Evolution,2013,299(7):1263~1274.
[18]唐璐璐,韩冰.开花式样对传粉者行为及花粉散布的影响[J].生物多样性,2007,15(6):680~686.
[19]HelenJ.Young.Sclection on spur shape in Impatiens capensis[J].Oecologia,2008,156:535~543.
[20]王洁.凹叶厚朴繁育系统研究及其濒危的生殖生物学原因分析[D].北京林业科学研究院,2012.
[21]Makoto Kato.Bumblebee visits to Impatiens spp.:pattern and efficiency[J].O ecologia,1988,76:364~370.
[22]何亚平,费世民,刘建全.高山植物繁育系统研究进展初探[J].四川林业科技,2005,26(8):43~49.
[23]Helen J.Young,Dan W.dunning,Kyle W.von Hasseln.Foraging behavior affects pollen removal and deposition in Impatiens capensis(Balsaminaceae)[J].American Journal of Botany,2007,94(7):1267~1271.
[24]肖宜安,何平,李晓红.濒危植物长柄双花木的花部综合特征与繁育系统[J].植物生态学,2004,28(3):333~340.
[25]Shi X,Wang J C,Zhang D Y.Pollen source and resource limitation to fruit production in the rare species Eremosparton songoricum(Fabaceae)[J].Nordic Journal of Botany,2010,28:1~7.
[26]郭华,王孝安,肖娅萍.植物交配系统及其在植物保护中的应用[J].西北植物学,2003,23(5):852~859.
[27]杨琨.植物繁育系统中传粉机制的多样性及交配系统研究评析[J].西安文理学院学报,2012,15(3):41~48.
[28]任明讯,姜新华,张大勇.植物繁殖生态学的若干重要问题[J].生物多样性,2012,20(3):241~249.
[29]袁存权.刺槐有性生殖过程及交配系统研究[D].北京林业大学,2013.
[30]Randall J Mitchell,Rebecca E Irwin,Rebecca J Flanagan,Jeffrey D Karron.Ecology and evolution of plant-pollinator interactions[J].Annals of Botany,2009,103:1355~1363.
[31]杨期和,杨和生,李姣清.植物自花授粉的类型及其适应性进化[J].嘉应学院学报,2011,29(8):55~64.
猜你喜欢
今日农业(2021年6期)2021-06-09
基层中医药(2020年10期)2020-11-27
课外生活(小学1-3年级)(2020年2期)2020-03-09
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
中国绿色画报(2016年9期)2017-05-31
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
中国民族医药杂志(2016年2期)2016-05-14
作品(2015年5期)2015-11-18
智慧与创想(2013年7期)2013-11-18
