
蜂虻寄生习性研究进展(双翅目:蜂虻科)
2014-11-25 02:59李文亮王国全
环境昆虫学报 2014年3期
姚 刚,李文亮,王国全,杨 定
(1.杭州市林业科学研究院,杭州 310016;2.河南科技大学林学院,洛阳 471003;3.广西大学农学院,南宁 530004;4.中国农业大学农业与生物技术学院,北京 100193)
蜂虻 (Bee fly)为双翅目Diptera 蜂虻科Bombyliidae 俗称,全世界已知16 亚科221 属5000余种,是双翅目昆虫中种类最多的类群之一。中国目前已知5 亚科28个属233 种。蜂虻体型变化大,体长2-20 mm,少数种类可达40 mm;其体色多样,通常被各种颜色的毛和鳞片,少数种类体光裸无毛,喙通常很长,翅通常有各种形状的斑;因部分种类模拟熊蜂、胡蜂或姬蜂的形态而得名“蜂虻”,为著名的拟态昆虫。蜂虻科成虫访花,幼虫寄生或捕食直翅目、鞘翅目、鳞翅目和其他双翅目的幼虫或卵,是重要的天敌昆虫,因此,蜂虻可以作为传粉昆虫和天敌资源昆虫加以利用,具有十分重要的经济价值。
蜂虻为复变态 (hypermetaboly,hypermetamorphosis)昆虫,不同龄期幼虫存在明显不同寄生生活方式。
蜂虻雌性成虫产卵量很大。Bohart 等(1960)和Marston (1964)报道Heterostylum robustum 和Anthrax limulatus Say 每天大约可以产卵1000个,Gerling and Hermann (1976)报道Xenox tigrinus(De Geer)可以产卵2000-3000个,Yeates (1997)报道Comptosia biguttata Yeates (Lomatiinae)每天能产卵约800个。

图1 蜂虻的生活史Fig.1 Generalized life-history of bee fly
1 研究简史
对蜂虻寄生的研究始于十九世纪末,Riley(1877,1878)首先报道了蜂虻取食蝗虫卵,这引起大家对蜂虻幼虫的生活习性的关注。二十世纪初期开始,陆续报道了蜂虻的寄生习性,到二十世纪中期,发表了大量关于蜂虻幼虫的生物学研究成果。这一时期有三个代表性的工作:一是Bohart 等 (1960)系统报道了在美国西北部Heterostylum robustum 寄生Nomia melanderi 在幼虫阶段的生物学习性,在蜂虻亚科Bombyliinae 中具有代表性。二是Du Merle (1964,1979a,b,c,1981)发表了一系列文章,完整地报道了内寄生蜂虻Villa brunnea 幼期生物学习性。Villa 属处于蜂虻科中种类最丰富的炭蜂虻亚科Anthracinae 中。三是Greathead (1958)详细研究非洲东部Systoechus Somali的生活史,研究发现其幼虫取食沙漠飞蝗Schistocerca gregaria的卵。Hull (1973)和Yeates(1997)分别总结了蜂虻科昆虫寄生习性。
2 寄生类型
蜂虻幼虫不同亚科不同属之间的寄生方式和类型存在一定的差异,根据羽化位置可归结为三种形式:外寄生、内寄生幼虫和内寄生蛹 (图2)。外寄生即在寄主表皮寄生,此为大多数蜂虻的寄生方式,内寄生幼虫指蜂虻在寄主处于幼虫阶段从其体内羽化出来,内寄生蛹是指蜂虻在寄主处于蛹期从其体内羽化出来。外寄生完成羽化之后可以在干瘪的寄主幼虫表皮上找到一个口钩大小的孔,一个脱落的蜂虻幼虫的蜕和一个成形的蜂虻蛹(图2,A)。(蜂虻的蛹在头、胸和腹部有特别的鬃和毛,因此很好辨认 (Hull,1973));幼虫可从头部表皮骨化形状,以及具有显著突出的上唇等特征进行辨认(Bohart et al.,1960)。炭蜂虻亚科Anthracinae 寄生时,如其寄主正经历幼虫到蛹期的蜕皮时期,在这种情况下,蜂虻寄生羽化后的蛹壳和末龄幼虫的蜕保存在寄主茧或者巢穴中(图2,C)。有证据表明,相对于炭蜂虻亚科Anthracinae的内寄生习性,弧蜂虻亚科Toxophorinae的蜂虻在寄主幼虫形成茧但还未化蛹之前从寄主体内羽化出来 (图2,B),此蜂虻在寄主的茧中化蛹。寄主的蜕保存在寄主的茧中,在寄主的蜕上有蜂虻羽化时留下的羽化孔。

图2 蜂虻科寄生类型Fig.2 Parasitic type of bee fly
而以观察蜂虻从不同类型的寄主中孵化出来为依据,可以将蜂虻寄生类型分为:内寄生,外寄生和取食蝗虫卵块等三种不同类型。
(1)内寄生(endoparasitoid)Marson (1964)详细报道了Anthrax fur (Osten Sacken)内寄生泥蜂科Sphecidae的生物学习性。在进入寄主之后蜂虻一龄幼虫取食量很少,一龄期较长,约为29 d,而后进入二龄幼虫阶段,该龄期幼虫发育很快,仅持续约7 d的时间,然后进入三龄幼虫阶段,持续约4 d,发育速度非常迅速,体长每天增加2.5 mm。最后A.fur 从寄主的蛹中羽化出来。Billiotti 等 (1965),Du Merle & Delpech (1973)报道了Villa brunnea 寄生松毛虫 Thaumetopoea pityocampa的生物学习性。V.brunnea的雌成虫在落叶层产卵之后,其闯蚴进入落叶层后需自行寻找寄主。闯蚴在未能找到寄主的情况下可存活1个月之久,其最远移动距离为1 m。一旦幼虫找到并侵入寄主,直至其羽化才从寄主体中出来。闯蚴仅能寄生处于蛹期的寄主,而当闯蚴遇到末龄寄主幼虫时,它将粘附在其表皮上,被带到寄主化蛹的地点(Du Merle,1979a),然后钻过寄主的表皮进入寄主的蛹中,钻入点通常位于寄主尾部折叠处的表皮(Du Merle,1979b)。进入寄主体内后,闯蚴附在表皮下,但不取食,寄主化蛹时幼虫即可在蛹内(Du Merle,1979c),同时迫使寄主形成一呼吸管。
幼虫在寄主蛹内的一、二龄发育情况未见报道。寄生后一个月左右幼虫蜕皮进入三龄幼虫期,这时寄主的内含物几乎被消耗掉,蜂虻在松毛虫空的蛹壳中化蛹,即将羽化时蜂虻的蛹利用头和胸部强壮的鬃和钩爬出寄主的蛹和茧到地表羽化。
(2)外寄生 (ectoparasitoid)早在1960年Bohart 等详细报道蜂虻Heterostylum robusturn 寄生茧蜂Nomia melanderi的生物学习性,蜂虻的一龄幼虫在寄主体中可长达36 h,体长由1.8 mm 增长至2.3 mm,直径增长为原来的2 倍多。一龄幼虫的位置不固定,经常转移位置。幼虫在二龄期间体长由2.3 mm 增长到8.3 mm,直径增长3 倍,此龄期的幼虫通常不动,弯曲于寄主体边,在受到干扰之后其细刀状的上颚能轻易地插入寄主之中,这时期寄主依然是存活的,但是在蜂虻二龄幼虫的取食下显著变得干瘪。三龄幼虫取食3 或4 d,长度增加1 倍。此蜂虻的幼虫可在单个寄主中完成发育,但其依然有能力去寻找并攻击新的寄主。蜂虻H.robusturn的幼虫为了完成发育先后寄生两个寄主时,它仅能消耗后一寄主的一半体积。根据Bohart的报道蜂虻H.robusturn 发育过程中幼虫体重增加了458 倍,十分高效地转移了寄主所包含的营养物质,蜂虻最终的体重与寄主被寄生前的体重近似。蜂虻H.robusturn的幼虫完成发育后从蜂的巢中出来在地表挖5-8 cm 深的洞越冬,此幼虫在蛹期前越冬。
(3)取食蝗虫卵块(Egg pod consumer)所有雏蜂虻属Systoechus (Bombyliinae:Bombyliini)的幼虫期均有取食蝗虫(Orthoptera,Acrididae)的卵块的习性。Hynes (1947)和Greathead (1958)报道了在非洲东部Systoechus somali 取食沙漠飞蝗Schistocerca gregaria 卵块的习性。蜂虻S.somali 成虫在距沙地面1-3 cm处悬飞时将卵轻弹到蝗虫卵块上方,每块蝗卵上产卵数量10 到40 粒不等。蜂虻卵孵化后,一龄幼虫只能从蝗虫卵块中通过取食前行,蝗虫卵被取食率从10%到100%不等。每头Systoechus 幼虫大概需要取食8-10 粒蝗虫卵来完成其发育,约为整个卵块中卵粒数量的15%。在一个卵块中可以找到1 到60 条不同龄期的蜂虻幼虫,但一般情况下数量都在10 以内。
蜂虻幼虫通常位于卵块的边缘或者在卵块上因取食而形成的凹陷内。幼虫取食蝗虫卵粒的时候,将口器对着卵粒的中部进行取食,卵粒因其内含物被蜂虻幼虫取食消耗而变得干瘪。蜂虻S.Somali的幼虫发育非常迅速,从到达蝗虫卵块开始取食计算只需要4 到11 d的时间就可完成幼虫阶段的发育。蜂虻幼虫往往取食单粒卵,不影响其它卵粒的发育,因此被取食的卵块中还有部分蝗虫卵可以正常孵化。通过蜂虻幼虫的取食,蝗虫卵块形成一个顶部1-2 cm 底部5-10 cm的椭圆形空室。
3 蜂虻的寄主范围
蜂虻在种级水平的寄生范围十分广泛,寄生发生主要的因素可能是一龄幼虫的生态环境而非寄主种类。例如,取食蝗虫卵块的幼虫种类,在相同的环境条件下发现其也取食其它幼虫。Anastoechus mylabricidus Zakhvatkin 被发现外寄生髓甲虫幼虫(Zakhvatkin,1934),而其幼虫是取食蝗虫卵块的。在属级水平上,寄主范围还有所增加,例如Petrorossia Bezzi 从蜘蛛类的卵囊中孵化出来,其幼虫还寄生很多其它的昆虫例如膜翅目昆虫和舌蝇科昆虫等 (Zaitzev and Charykuliyev,1981)。Exhyalanthrax 属的种类通常寄生土栖的双翅目昆虫,并从其蛹中羽化出来,但同时也有报道从黄蜂,飞蛾,叶蜂的茧中孵化出来,还有的从蟑螂卵鞘中孵化出来 (Greathead and Grandcolas,1995)。这些寄主都是土栖昆虫。而Exhyalanthrax属的种类寄生膜翅目和双翅目,所有的寄主都是生活在同类环境中。可见蜂虻科昆虫的寄主种类的专一性并不是很强。
不同于膜翅目和其它双翅目如头蝇科Pipunculidae 和寄蝇科Tachinidae,为什么蜂虻科昆虫的寄主会有如此广泛?Eggleton and Belshaw 在1993 指出可能是由于蜂虻科昆虫未能进化成穿刺型的产卵器,而导致无法直接穿入寄主体内,这也是所有蜂虻的一龄幼虫都能活动并攻击寄主的原因。少部分蜂虻属昆虫如Lordotus (Lordotinae)和Antonia (Antoniinae)进化出特别的雌性产卵器(图3B),与产卵行为相关,据推测这些是用来刺入寄主所处的环境如土壤中,而非刺入寄主体内产卵。蜂虻科中仅Toxophorinae 亚科的Systropus 属的雌性成虫进化为产卵与寄主直接接触。因此仅Systropus 属的蜂虻寄主具专一性,且目前仅知其寄主为刺蛾科Limacodidae 昆虫。
蜂虻寄主范围广泛的另一原因可能是雌成虫将卵散播于合适的环境中,蜂虻一龄幼虫(闯蚴)自行寻找寄主,但其活动范围有限,在这种情况下,寄主范围较大的蜂虻比寄主选择范围窄的蜂虻更具选择优势(Vinson,1976)。
4 应用寄生习性分析蜂虻系统发育
4.1 寄主类型与蜂虻的系统发育
迄今为止,有大量的报道记录蜂虻寄主,我们收集整理了990个报道记录并进行汇总(见表1)。依据Yeates (1994)提出完整的蜂虻寄生系统发育假说进行分析,乌蜂虻亚科Usiinae、弧蜂虻亚科Toxophorinae、蜂虻亚科Bombyliinae、炭蜂虻亚科Anthracinae 下各族的寄生习性和寄主表现出内部趋同性(表1),对指导和印证高级阶元系统发育关系具有重要的意义。同一亚科或者族下的不同属与不同的寄主之间存在联系(例如:蜂虻亚科寄生蜜蜂和黄蜂的幼虫或者取食直翅类的卵块),但由于未能找到属之间更加详细的联系,我们还没发现其寄主的变化和进化方向的关系。一旦形成完整的属级阶元的系统发育假说,并且发现更多蜂虻属的寄主,将能更加有力地证明其寄主与进化之间的相互关系。
炭蜂虻亚科Anthracinae 共有超过40%的已知寄生的种类,超过60%寄生记录数(表1)。这主要是由于Anthracini 占据寄生记录数的34%和种类数的6%。这可能是由于Anthracini 寄主比较容易采集和观察。岩蜂虻属Anthrax Scopoli 物种丰富,分布广泛,且其幼虫外寄生于蜜蜂和黄蜂,这可能是其记录较多的原因。
目前关于蜂虻寄生习性的报道和记录都是属于偶然发现的。其中大部分报道来自对昆虫生态学习性研究(e.g.Aiello,1980),部分来源于对寄主生物学习性研究 (Knisley 和Pearson,1981)。Du Merle (1975)指出,蜂虻寄生习性都是在饲养寄主的时候偶然情况下发现的。蜂虻寄生记录突然增加,也是得益于针对寄主饲养的研究计划增加 (e.g.Brooks,1952),这可能 是导致Anthracinae 亚科中报道居多的主要原因。

表2 蜂虻寄主统计Table 2 Summary of Bombyliidae host species
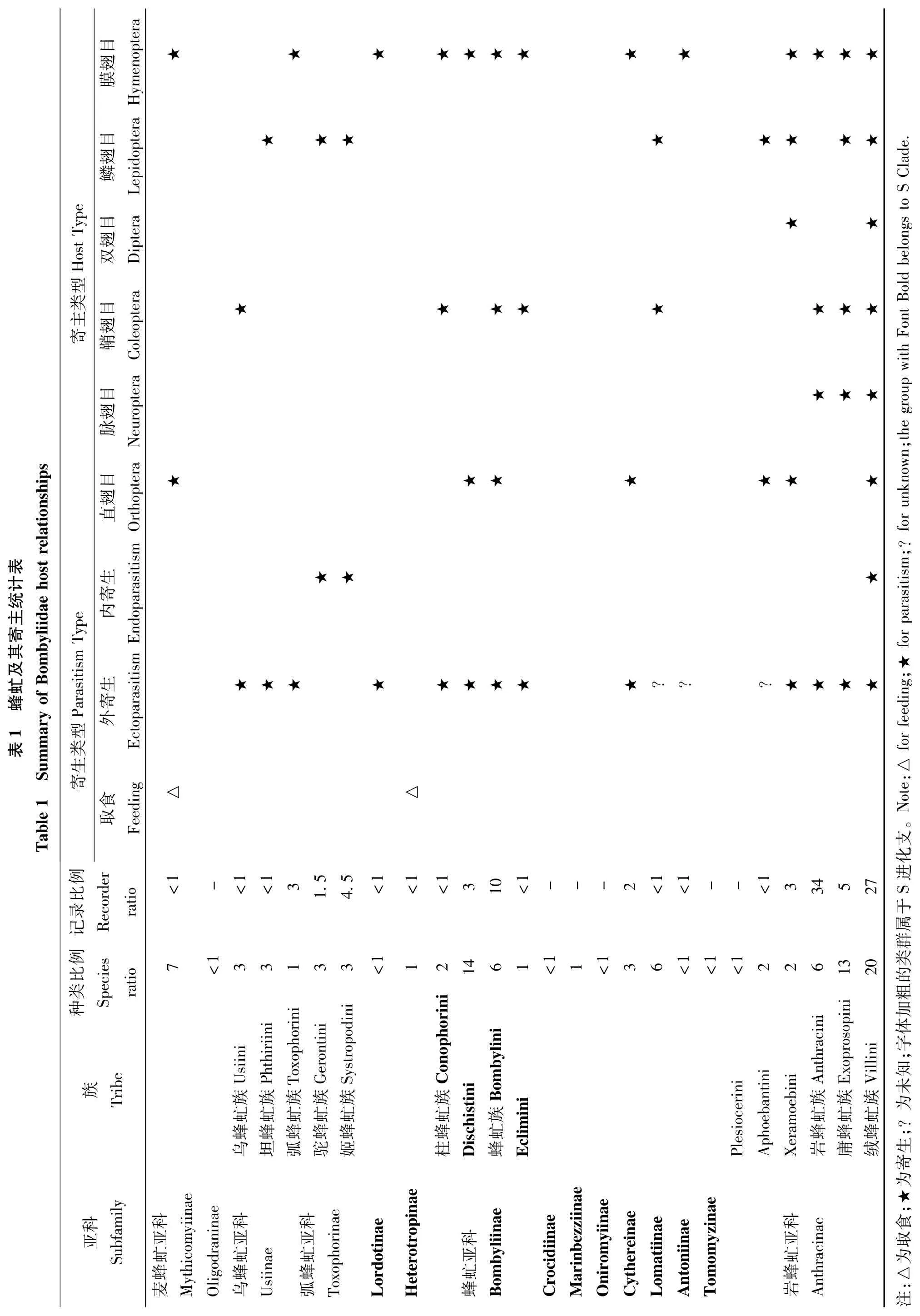
表2 将寄生记录分列于几个类型的寄主中。从表中可以看出几乎半数的寄生记录数(48%)来自于蜜蜂和黄蜂,而这大多数的记录来自于动物行为学家的采集。蜂虻科寄生蛾类、叶蜂、蝗虫和螽斯的记录(39%)主要来自农业和林业的昆虫学家的研究。蜂虻对舌蝇的寄生(5%)主要来自医生和兽医的贡献。从表中分类表象和数据来看,由于很多蜂虻科昆虫的生活史很难研究或很少研究而导致统计数值偏小,例如土栖的鞘翅类幼虫和幼期生活在朽木中的昆虫。
4.2 蜂虻产卵方式的进化
绝大部分蜂虻科昆虫其雌性成虫的腹部末端在生殖腔后面有一个袋,其术语叫做“sand chamber”(Schremmer,1964),周边着生长刚毛(图3A),从系统发育角度来看它们的亲缘关系比较接近,我们将这些类群简称为“S 进化支”(图4)。Painter (1932)首先注意到雌性蜂虻复杂的产卵行为与“沙室”的功能和关系,具有“沙室”的雌性成虫都是在悬停时产卵。

图3 A,Bombylius major Linnaeus 雌性外生殖器的“沙室”;B,Antonia suavissima Loew Antoniinae 雌性外生殖器无“沙室”Fig.3 A,Bombylius major Linnaeus,female genitalia with“sand chamber”;B,Antonia suavissima Loew Antoniinae,female genitalia without“sand chamber”
S 进化支中的雌性蜂虻直接产卵到潜叶寄主巢的边上(Bohart et al.,1960),或者地表任意位置(Du Merle and Delpech,1973),因此至少部分一龄幼虫需自行寻找寄主。幼虫主要在落叶层寻找寄主的情况称为底部区域(substrate zone)的习性(Hlavac,1971)。小部分蜂虻科的种类如Beckerellus Greathead (Bombyliinae) 和 Antonia Loew (Antoniinae),其产卵器为管状,属于S 进化支,但无沙室(图3B)。虽然目前对这一类群的产卵行为并不了解,但是从产卵器在形态上存在显著差异,推断可能与有沙室的类群的产卵行为截然不同。

图4 蜂虻亚科进化分枝图Fig.4 Cladogram of Bombyliidae subfamilies from Yeates (1994)
这个类群的很多种类在地面之上攻击寄主,可以断定存在沙室的蜂虻与寄主的分布区更加接近。姬蜂虻属Systropus Wiedemann (Toxophorinae)的雌虫将卵直接产在刺蛾科幼虫身上 (Genty,1972;Greathead,1987)。乌蜂虻属Usia Latreille雌虫在细枝上停息的时候将卵散播于其表面(Du Merle,1971)。其它属例如弧蜂虻属Toxophora Meigen 雌虫将卵产于地表。虽然目前为止还没确凿的证据,但是可以推测这一类群的蜂虻可直接将卵产于寄主巢穴。
将蜂虻科与寄蝇科(双翅目环裂亚目)产卵行为进行比较具有十分重要的启发性。寄蝇科常见的产卵方式是将卵产于寄主表皮 (Wood,1989),而蜂虻昆虫仅有姬蜂虻属Systropus为此种产卵方式。部分寄蝇科昆虫将卵产到寄主体内或者产于寄主取食的植物表面,而截止目前,我们尚未发现任何蜂虻科昆虫存在以上两种产卵方式。部分寄蝇的种类和大部分蜂虻科的种类(S 进化支)将卵散播于适合于寄主栖息的环境中。寄蝇科昆虫的活动区域以及寄生的种类和数量上都远远超过蜂虻科昆虫。因此在同一区域内膜翅目昆虫和寄蝇的寄生优越性,在一定程度上限制了蜂虻科昆虫的寄生。这也可以从侧面解释为什么具有沙室的蜂虻昆虫与在进化支上较原始蜂虻种类具有更多的产卵方式。
5 蜂虻在害虫防治中的应用
Parker 和Wakeland (1957)年整理了北达科他州1938、1939 和1940年对蝗虫卵的调查数据,发现蜂虻科卷蜂虻属Systoechus 不仅是调查中最常见的天敌,而且分布广泛、取食率高,蝗虫卵单年被蜂虻取食率最高可达77.52%。国内对蜂虻科昆虫作为天敌昆虫防控害虫的研究开始于1993年杜树国等对东亚飞蝗的重要天敌中华雏蜂虻进行了详细的研究,从中华雏蜂虻的形态特征、生物学特性、适生环境、保护利用等方面进行详细研究,并在2003年田方文等报道中山东无棣县应用中华雏蜂虻Anastoechus chinensis Paramonov 防控环渤海湾蝗区东亚飞蝗Locusta migratoria manilensis Meyen 奠定了坚实的基础,中华雏蜂虻以幼虫取食蝗虫卵,并且可以通过种植双色补血草等蜜源植物来吸引蜂虻,从而达到控制蝗虫数量的目的。国内其它地区还报道了多型岩蜂虻Anthrax distigma Wiedemann,其幼虫寄生于胡蜂及切叶蜂,大蜂虻Bombylius major Linnaeus 捕食蚜虫、叶蝉、蛾类及膜翅目昆虫幼虫(杨彦龙和任炳忠,2003)。另外,近些年还有一些报道中华雏蜂虻取食蝗虫卵作为天敌的应用的(王振平和严毓骅,1999;郝伟等,2007;胡奇和张龙,2007)。我国对蜂虻科昆虫作为天敌资源昆虫在农林产业中应用的研究还处于起步阶段。
6 结论与展望
蜂虻科昆虫种类资源丰富,分布广泛,从天敌昆虫应用角度来说,在我国北部地区和西部地区半干旱和干旱的条件下具有比较显著的优势,特别体现在对蝗虫的防控上。从目前国内外的研究来看,我们对蜂虻科昆虫的研究还是十分的匮乏,对于大多数蜂虻科昆虫的生活史了解甚微,更别说其生物学习性等,这使我们目前还无法将其作为天敌昆虫有效利用。因此我们需要更加深入地开展研究,摸清我国蜂虻科昆虫的资源种类、研究其各种的生活史,生物学特性,寄主专一性等,以便其可以在农林生产实践中得到更好的利用,提升害虫防控效率。
References)
Aiello A.Systropus columbianus (Diptera:Bombyliidae)reared from larva of limacodid moth [J].Entomological News,1980,91:89-91.
Billiotti E,Demolin G,Du Merle P.Parasitisme de la Processionnaire du pin par Villa quinquefasciata Wied.apud Meig.(Dipt.Bombyliidae).Importance du comportement de ponte du parasite[J].Annales des Epiphyties,1965,16:279-288.
Bohart GE,Stephen WP,Eppley RK.The biology of Heterostylum robustum (Diptera:Bombyliidae),a parasite of the alkali bee[J].Annals of the Entomological Society of America,1960,53:425-435.
Brooks A.Identification of bombyliid parasites and hyperparasites of Phalaenidae of the prairie provinces of Canada,with descriptions of six other bombliid pupae (Diptera)[J].Canadian Entomologist,1952,84:357-373.
Du Merle P.Cycle biologique d'un Diptère Bombyliidae du genre Villa[J].Comptes Rendus Hebdomadaires des Seances.Academie des Sciences,1964,259:1657-1659.
Du Merle P.Biologie de deux espèces du genre Usia Latreille (Dipt.,Bombyliidae)[J].Annales de la Société de Entomologique de France,1971,7:241-259.
Du Merle P.Les hôtes et les stades pré-imaginaux des Diptères Bombyliidae:revue bibliographique annotée [J].Bulletin de la Section Regionale Ouest Palearctique (Organisation Internationale de Lutte Biologique),1975,4:1-289.
Du Merle P.Biologie de la larve planidium de Villa brunnea Beck.,Diptère Bombyliide parasite de la processionnaire du pin.Ⅰ.Rechereche et decouverte de l' hôte [J].Annales de Zoologie-Ecologie Animale.Institute National de la Recherche Agronomique,1979,11:289-304.
Du Merle P.Biologie de la larve planidium de Villa brunnea Beck.,Diptère Bombyliide parasite de la processionnaire du Pin.Ⅱ.Infestastion de l' hôte [J].Annales de Zoologie-Ecologie Animale.Institute National de la Recherche Agronomique,1979,11:305-326.
Du Merle P.Biologie de la larve planidium de Villa brunnea,Diptère Bombyliide parasite de la processionnaire du Pin.Ⅲ.Le début de la reactions de l'hite et les échecs du parasitisme [J].Annales de la Société Entomologique de France,1979,15:525-551.
Du Merle P.Biologie de la larve planidium de Villa brunnae [Dipt.Bombyliidae]parasite de la processionaire du pin.Ⅳ.Intensité des reactions siphonogänes de l' hôte [J].Annales de la Société Entomologique de France,1981,17:191-206.
Du Merle P,Delpech M.Dispositifs de terrain destinés à l' étude de la biologie et du role limatif de Villa brunnae (Dipt.,Bombyliidae)vis-a-vis de Thaumetopoea pityocampa (Lep.,Thaumetopoeidae)[J].Annales de la Société de Entomologique de France,1973,9:471-482.
Du SG,Li SZ,Liu DP,Wang CS,Chang ZZ.Studies onAnstoechus chinensis Paramonov—a natural enemy of Locusta migratoria manilensis (Meyen) [J].Acta Entomologica Sinica,1993,36(4):444-450.[杜树国,李述增,刘德萍,王春山,常兆芝.东亚飞蝗天敌——中国雏蜂虻的研究[J].昆虫学报,1993,36 (4):444-450]
Eggleton P,Belshaw R.Comparisons of dipteran,hymenopteran and coleopteran parasitoids:provisional phylogenetic explanations [J].Biological Journal of the Linnean Society,1993,48:213-226.
Genty P.Morphologie et biologie de Sibine fusca Stoll,lépidoptère défoliateur du palmier à huile en Colombie [J].Oléagineux,1972,27:65-71.
Gerling D,Hermann H.The oviposition and life cycle of Anthrax tigrinus(Dipt.:Bombyliidae) a parasite of carpenter bee (Hym.:Xylocopidae)[J].Entomophaga,1976,21:227-233.
Greathead DJ.Observations on two species of Systoechus (Diptera:Bombyliidae) preying on desert locust,Schistocerca gregaria(Forskal),in eastern Africa [J].Entomophaga,1958,4:3-22.
Greathead DJ.Bombyliidae.A summary of the recorded bombyliid parasitoids of south-east Asian Limacododidae.In:Cock M,Godfray HCJ,Holloway JD,eds.Slug and Nettle Caterpillars:the Biology,Taxonomy and Control of the Limacodidae of Economic Importance on Palms in South-east Asia [M].Wallingford:CAB International,1987,195-196.
Greathead DJ,Grandcolas P.A new host association for the Bombyliidae(Diptera):an Exhyalanthrax sp.reared from cockroach oothecae,Heterogamisca choprdi (Dictyoptera:Polyphagidae) in Saudi Arabia [J].The Entomologist,1995,114:91-98.
Hao W,Jiang XL,Zhang DJ,et al.Wang AD.The natural enemies of Locusta migratoria manilensis in the Yellow River beach [J].Chinese Bulletin of Entomology,2007,44 (3):406-409.[郝伟,江新林,张冬菊,等.黄河滩区东亚飞蝗天敌种类及其消长规律[J].昆虫知识,2007,44 (3):406-409]
Hlavac TF.The prothorax of Coleoptera:origin,major features of variation [J].Psyche,1971,78:51-66.
Hu Q,Zhang L.Review of research progress on locust natural enemies[J].China Plant Protection,2007,27 (4):13-16.[胡奇,张龙.蝗虫天敌昆虫研究概述[J].中国植保导刊,2007,27(4):13-16]
Hull F.Bee flies of the world.The genera of the family Bombyliidae[M].Washington D.C.:Smithsonian Institution Press,1973,687 pp.
Hynes HBN.Observations on Systoechus somali (Diptera Bombyliidae)attacking the eggs of the desert locust (Schistocerca gregaria(Forskål)) in Somalia [J].Proceedings of the Royal Entomological Society of London (A),1947,22:79-85.
Knisley C,Pearson D.The function of turret building behavior in the larval tiger beetle,Cincindela willistoni (Coleoptera:Cicindelidae)[J].Ecological Entomology,1981,6:401-410.
Marston N.The biology of Anthrax limulatus fur (Osten Sacken),with a key to and descriptions of pupae of some species in the Anthrax albofasciatus and Trimaculatus groups (Diptera:Bombyliidae) [J].Journal of the Kansas Entomological Society,1964,37:89-105.
Painter R.The Bombyliidae of China and nearby regions [J].Lingnan Science Journal,1932,11:341-374.
Parker JR,Wakeland C.Grasshopper egg pods destroyed by larvae of bee flies,blister beetles,and ground beetles [J].U.S.Dept.Agric.,Tech.Bull.,1957,1165:1-29.
Riley CV.Ninth annual report on the noxious,beneficial and other insects of the State of Missouri,made to the State Board of Agriculture [R].Jefferson City,Missouri,1877,1-129.
Riley CV.First annual report U.S.Entomological Commission for the year 1877,relating to the rocky mountain locust [R].Dept.Interior,U.S.Geol.Surv.,1878,16,1-477.
Schremmer F.Gezielter abwurf getarnter eier bei wollschwebern (Dipt.Bombyliidae)[J].Zoologischer Anzeiger,1964,27:291-303.
Tian FW,Sun FL,Zhang XA.Study on the relationship between nectariferous plant and Anstoechus Chinensis Paramonov [J].Pesticides,2003,42 (10):914-916.[田方文,孙福来,张秀安.蜜源植物与中国雏蜂虻发生关系的研究.生物防治,2003,42 (10):914-916]
Vinson SB.Host selection by insect parasitoids [J].Annual Review of Entomology,1976,1:109-133.
Wang ZP,Yan YH.The research advances in utilizing natural enemies of grasshopper.Grassland of China [J],1999,6:54-58.[王振平,严毓骅.蝗虫天敌可利用性分析及研究进展.中国草地[J],1999,6:54-58]
Wood DM.Tachinidae.In:McAlpine J,ed.Manual of Nearctic Diptera.Vol.2[M].Hull,Canada:Research Branch Agriculture,1989,1193-1269.
Yang YL,Ren BZ.The study on the fauna and diversity of natural enemies of insects from Jilin Province (III)[J].Journal of Chang Chun Teacher College,2003,22 (2):69-74.[杨彦龙,任炳忠.吉林省农林天敌昆虫区系及多样性的研究(III)[J].长春师范学院学报,2003,22 (2):69-74]
Yeates DK.Cladistics and classification of the Bombyliidae (Diptera:Asiloidea)[J].Bulletin of the American Museum of Natural History,1994,219:1-191.
Yeates DK,Greathead DJ.The evolutionary pattern of host use in the Bombyliidae (Diptera):a diverse family of parasitoid flies [J].Biological Journal of the Linnean Society,1997,60:149-185.
Zakhvatkin AA.The parasites of the Moroccan locust in Azerbaidyan (In Russian)[J].Trudy po Zascite Rastenji,1934,9:52-71.
Zaitzev VF,Charykuliyev D.On the biology of bee-flies of the genus Petrorossia Bezzi (Diptera,Bombyliidae)with description of a new species from Turkmenia [J].Entomologischeskoe Obozrenie,1981,60:914-916.
猜你喜欢
农村青少年科学探究(2022年3期)2022-05-13
环境昆虫学报(2020年6期)2021-01-18
海峡姐妹(2020年11期)2021-01-18
武夷科学(2019年2期)2019-12-20
植物保护(2019年5期)2019-11-27
现代园艺(2017年21期)2018-01-03
武夷科学(2017年0期)2017-03-10
郑州大学学报(理学版)(2012年4期)2012-03-25
中国蔬菜(2012年24期)2012-02-24
