
绵麦37特异位点在其衍生品种中的遗传贡献率分析
2014-11-17 06:42:32任勇李生荣罗建明何中虎杜小英周强何员江魏育明郑有良
遗传 2014年2期
任勇,李生荣,罗建明,何中虎,杜小英,周强,何员江,魏育明,郑有良
1.四川农业大学小麦研究所,成都 611130;2.绵阳市农业科学研究院,国家小麦改良中心绵阳分中心,绵阳 621023;3.中国农业科学院作物科学研究所,国家小麦改良中心,北京100081
亲本选配是小麦杂交育种成败的关键因素之一,好的亲本应该具有配合力高、优良性状遗传力强等优点。在长期育种实践过程中,一些亲本品种(系)在不同时期和不同生态区的小麦品种更新换代中发挥了核心作用,衍生出一系列大面积推广品种,为小麦育种和生产做出了突出贡献,成为骨干亲本[1,2]。通过系谱分析和衍生品种推广应用情况,已明确了碧蚂4号[3]、燕大1817[4]、欧柔[5]、矮孟牛[6]、小偃6 号[7]、繁 6[8]、周 8425B[9,10]和鲁麦 14[11]等 20 余个小麦骨干亲本。这些品种除本身具有突出优良性状外,其重要的农艺性状如丰产性和抗病性等具有较强的传递能力,在后代选择过程中被高频率保存下来,对后代品种产量潜力提高起到重要作用[11,12]。品种改良过程必须以育种材料整体水平的提高为基础。一些优良的农艺性状只有被转至综合性状好的背景中,才易被生产利用或成为优秀的杂交亲本[13]。每一批大面积推广品种的形成都与一个或数个育种骨干亲本的出现密不可分,突破性亲本的出现是推动品种更新换代的内在动力[2]。如通过聚敛杂交法育成的繁6,衍生出了绵阳11号、绵阳26号等38个品种[8,14,15]; 周 8425B矮秆、高产、配合力好、抗条锈病和白粉病,用做亲本育成矮抗58和周麦16等22个品种[12]; 鲁麦14既是好品种,又是好亲本,用做亲本育成济麦22和良星99等11个品种[11,12];6VS/6AL 易位系(如92R137等)[16,17]和人工合成小麦[18]作为优良亲本也育成了一系列品种。绵麦 37是绵阳市农业科学研究院育成的小麦品种,具有丰产性好、抗病性突出、品质优良、矮秆抗倒等特点,自2004年审定以来在四川大面积推广,2008年至今作为四川省小麦区域试验对照品种[19]。绵麦37高抗条锈病和白粉病,尽管在四川已发现对Yr24/Yr26具有毒性的条锈菌株 V26,四川部分小麦品种已丧失抗性[20],但绵麦37依然表现高抗,其抗性已持续14年。目前,绵麦37已成为四川小麦育种的重要亲本材料,育成了一系列小麦新品种(系)。这些衍生品种(系)均表现出良好的丰产性和突出的抗病性,出现了绵麦 367等单产 9 000 kg/ha以上的高产典型品种[21]。因此,研究绵麦37特异位点在其衍生品种中的遗传贡献,为了解绵麦37对其衍生品种影响机制提供依据,为进一步发挥这些特异位点在四川小麦育种中的作用奠定基础。
1 材料和方法
1.1 材料及来源
亲本品种绵麦 37(96EW37/绵 90-100)及其 4个衍生品种绵麦367、绵麦51、BL227和绵麦228由本课题组育成提供。其中,绵麦367和绵麦51为国审品种,来自于同一组合,系谱为绵麦 37/川麦 43;绵麦228和BL227为四川省审定品种,来自于同一组合,系谱为绵麦 37/内麦 8号//川麦 43。川麦 43由四川省农业科学院作物研究所提供,感病对照品种铭贤169由本课题组繁殖。
1.2 试验设计
2009~2012年连续3年在绵阳松垭试验田调查绵麦37及其衍生品种产量性状。试验采用随机区组设计,3次重复,小区面积12 m2,小区间走道0.4 m,区组间走道0.5 m,试验四周设置保护行。条沟点播,出苗后通过定苗,确保各小区基本苗一致。
田间调查基本苗数、有效穗数和每穗粒数,成熟期取样考种,测定千粒重,收获整个小区测定小区产量。
1.3 抗条锈性鉴定
苗期抗条锈性鉴定在温室内进行,选用小麦条锈菌生理小种CYR29、CYR31、CYR32、CYR33、SUN11-4、SUN11-7和V26(由西北农林科技大学植物保护学院植物抗病遗传研究室提供) 接种鉴定。成株期抗条锈鉴定在田间进行,采用混合小种接种鉴定。待感病对照铭贤 169 充分发病后,调查各参试材料反应型,记载采用0、0、1、2、3、4 的6级标准。
1.4 DNA提取及SSR标记检测
采用CTAB 法提取小麦基因组DNA。根据SSR图谱[22],选择分布在 21条染色体上的 292对 SSR引物,根据http://wheat.pw.usda.gov报道序列合成。PCR扩增体系总体积为 20 µL,包含 1×buffer(100 mmol/L Tris-HCl,pH 8.3,1.5 mmol/L MgCl2),0.2 mmol/L dNTPs,240 nmol/L引物,1 UTaqDNA 聚合酶,50~100 ng模板DNA。PCR 扩增程序为:94℃预变性5 min; 94℃变性1 min,50~60℃复性1 min,72℃延伸1 min,共38个循环; 72℃延伸10 min。扩增产物用6%变性聚丙烯酰胺凝胶电泳检测,经硝酸银染色后观察照相。
1.5 数据处理
采用Microsoft Excel 2007和SPSS19.0软件对产量性状进行方差分析。根据PCR扩增结果,计算衍生品种中绵麦37特异位点遗传贡献率,即后代中扩增出绵麦 37特异位点数与双亲多态位点数的百分比。根据 http://wheat.pw.usda.gov已知 SSR引物位置,用Mapdraw软件绘制遗传连锁图。
2 结果与分析
2.1 产量构成及抗条锈性
绵麦37及其衍生品种产量构成见表1。由表1可知,4个衍生品种产量水平显著高于其亲本绵麦37,表明衍生品种丰产性明显提高,表现出较强的超亲遗传。其中绵麦367表现尤为突出,较绵麦37增产21.4%; 其次是绵麦51、绵麦228和BL227,分别增产16.7%、15.5%和 14.9%; 从产量构成要素上看,穗粒数的显著提高是衍生品种增产的主要因素。
用当前主要条锈菌流行小种分别鉴定绵麦37、川麦43、内麦8号及后代品种的抗条锈性,结果表明,绵麦37对7个小种及混合小种均表现免疫或近免疫,川麦43和内麦8号对V26分别表现中抗和中感; 后代品种中除绵麦 228对 CYR33表现感病外,其余衍生品种对所有鉴定小种均表现抗病,但反应型有差异。由此表明,绵麦 37优良的抗条锈性,特别是对V26的抗性较好地传递给了后代品种(表2)。
2.2 绵麦37特异位点检测
利用均匀分布在 21条染色体上的 292对 SSR引物筛选绵麦 367双亲多态性,共 114对引物能在绵麦37与川麦43的基因组DNA中扩增出稳定的多态性片段,多态性比例达39.0%。用这114对具有稳定扩增结果和清晰条带的 SSR引物检测绵麦37及其衍生品种绵麦367,结果表明,有90对SSR引物在两者间扩增出稳定一致的条带,表明绵麦 367的这些位点来自于绵麦 37,其遗传贡献率达 78.9%。利用部分绵麦37特异位点检测所有衍生品种,均表明这些位点来自于绵麦37(图1)。
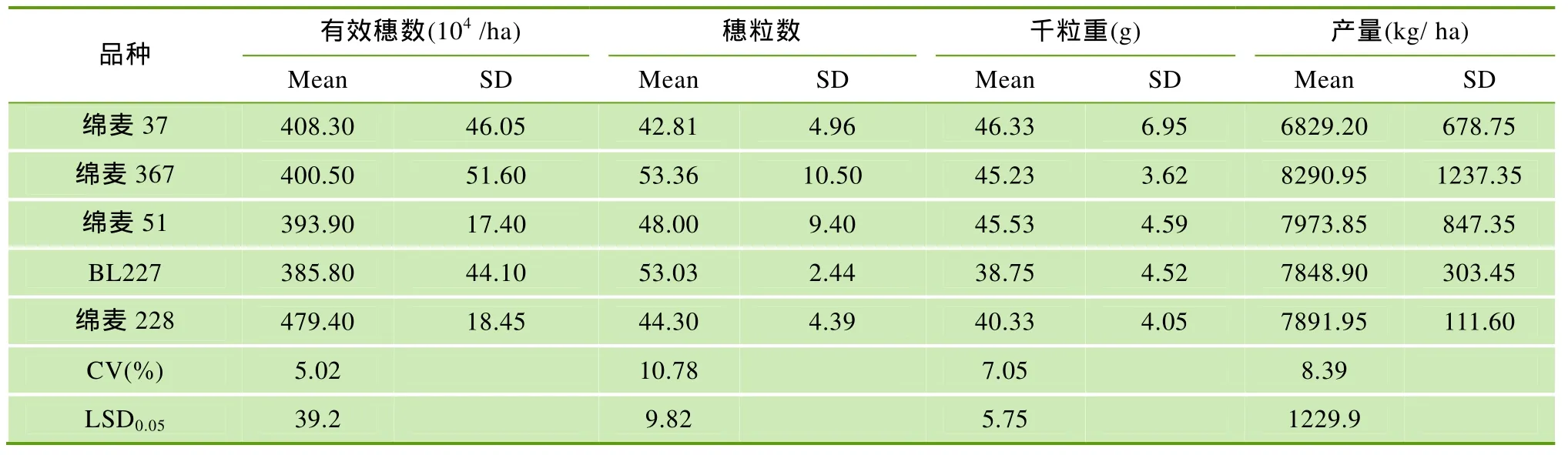
表1 绵麦37及其衍生品种产量性状平均值、标准差及差异显著性
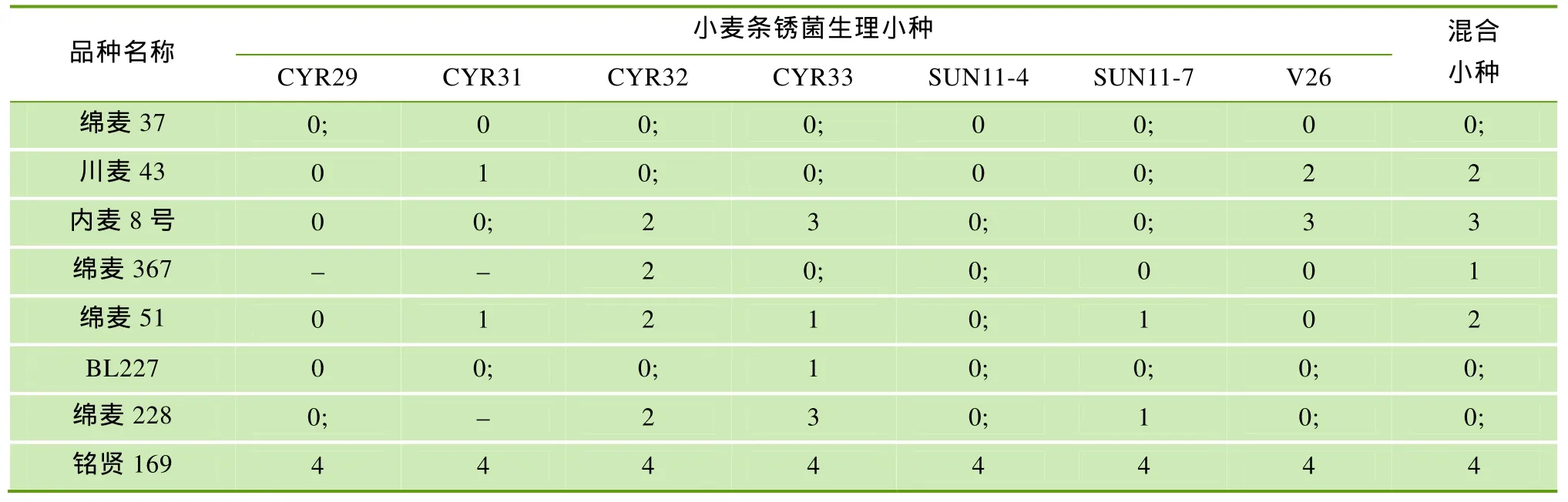
表2 绵麦37及其衍生品种抗条锈鉴定结果

图1 SSR引物Barc267-7B(A)和WMC291-3B(B)在不同品种中的扩增结果
2.3 绵麦37特异位点不同染色体分布及遗传贡献
检测到的90个绵麦37特异位点在绵麦367的A、B、D基因组中分布是不均匀的,分别为21、46和 23,且 B组>D组>A组; 遗传贡献率为 B组(83.6%)>A组(75.0%)>D组(74.2%)(表3)。在绵麦367的大多数染色体上都检测到来自绵麦37的遗传位点,但不同染色体间存在较大差异。其中,遗传贡献率大于70%的染色体有5A、1B、2B、3B、5B、6B、2D和5D,部分遗传位点在染色体上的分布见图2。由图2可知,绵麦367与绵麦37相同染色体片段主要为2B的Xgwm374-Xbarc167-Xbarc128-Xgwm129-Xgwm388-Xbarc101,3B的Xwmc446-Xwmc366-Xwmc533-Xbarc164-Xwmc418等区段。这些区段包含了许多重要性状位点如穗粒数[23]、千粒重[24]和抗病性[25,26]等。表明绵麦37特异位点在育种家定向选择的作用下,较好地传递给了后代品种。
3 讨 论
建国以来四川的小麦生产大致经历了6次大的品种更替,特别是以繁6及其衍生品种绵阳11号、绵阳26号等的大面积推广,将四川及长江上游麦区小麦单产和品质都提高到一个新的水平[14,15]。但由于条锈菌新小种的出现,这些品种相继丧失抗性,退出生产。2004年前后,以绵麦37和川麦42为代表的一批高产抗病品种及其衍生品种的育成并迅速推广,四川小麦产量水平跃上了一个新的台阶,出现了一批单产9 000 kg/ha、个别达10 500 kg/ha以上的高产品种[21,27],为四川的小麦生产做出了突出贡献。绵麦37不仅作为优良品种在生产上大面积推广应用,作为重要亲本材料已育成一系列新品种(系)[19]。本研究结果表明,这些衍生品种的丰产性均显著提高,表现出明显的超亲遗传。另一方面,由于条锈菌新菌株 V26在四川被首次发现,四川小麦如川麦 42、MR168等品种(系)分别携带的抗条锈病基因YrCH42和YrMR168已丧失抗性[20,28,29]; 6VS/6AL易位系和贵农号品系所携带的主效基因Yr24/Yr26已开始感病,引起了植病学家和育种家的高度重视[12,30,31]。据四川省农业科学院植物保护研究所2003~2012年连续10年鉴定,绵麦37对四川主要条锈菌流行小种均表现免疫至高抗; 2009~2010年田间接种V26,绵麦37成株期表现免疫[19,32],与本研究鉴定结果一致。绵麦 37对条锈菌新菌株 V26的抗性受一对隐性基因控制,被初步定位在7B染色体上(另文发表),与其紧密连锁的SSR标记Barc267
在后代品种中均被检测到(图1),表明该抗性基因很好地传递给了后代品种。这些新品种的育成将对进一步提高四川小麦单产、减轻条锈病危害发挥重要作用。
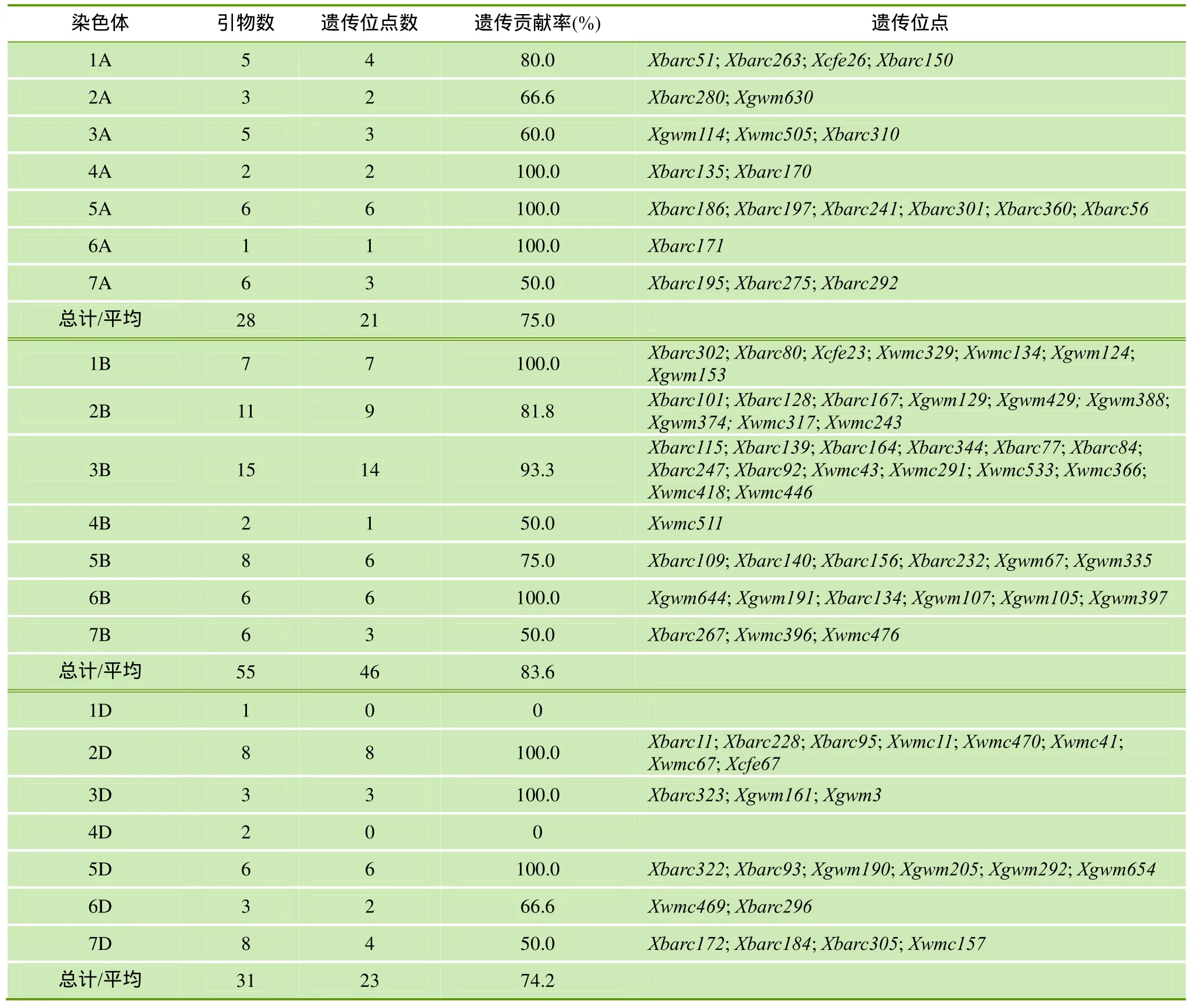
表3 绵麦37特异位点不同染色体的遗传贡献率
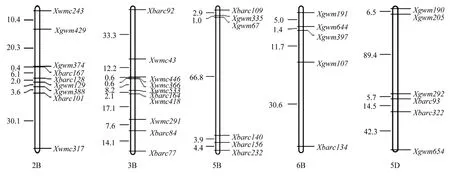
图2 绵麦367部分染色体上来自于绵麦37的遗传位点
利用分子标记追踪骨干亲本特异位点在后代衍生品种中的遗传轨迹,有利于优异基因的挖掘。陈国跃等[8]利用骨干亲本繁 6及其衍生品种的条锈病成株期抗性与SSR标记关联分析,发现了6个与小麦条锈病成株期抗性显著相关的遗传位点; 李小军等[5]发现了小麦骨干亲本欧柔中Xwmc710、Xbarc235和Xbarc252等3个位点高频率传递给后代并与穗粒数和产量相关。廖杰等[33]检测川麦42遗传背景中人工合成小麦位点显著低于理论值,但这些位点可能与选择目标性状如抗条锈性密切相关。本研究发现,高产品种绵麦367中来自于绵麦37的位点达78.9%,显著高于理论值50%,部分染色体更是100%来自于绵麦37。这可能是由于人工选择的结果,导致绵麦37遗传位点在后代A、B、D染色体组以及不同染色体上分布不均。那些被高频率保留的位点,可能与育种目标性状如穗粒数、抗病性紧密连锁,在人工定向选择压力下,较好地传递给了后代。根据产量构成分析,衍生品种的丰产性提高主要来自于穗粒数的贡献,这与育种学家选择大穗型品种的育种目标是吻合的。但是,哪些位点与穗粒数相关?哪些位点与抗病性相关?还有待进一步研究。
国内外长期育种实践表明,用于杂交的两个亲本材料,其综合性状应该尽可能好; 两亲本不能有共同的缺点,互补性要强,产量构成因素的一般配合力要好,遗传关系应相对较远[1,2]; 这样后代理想基因型出现的机会更大。本研究发现,绵麦37与川麦43遗传多样性达39.0%,这可能是该组合能够快速育成绵麦367等4个高产品种以及一批新品系的原因之一。值得指出的是,优良亲本的创制和利用能有效提高育种效率,但应避免过分集中使用单一亲本,导致遗传基础狭窄和抗性过快丧失。
[1]庄巧生.中国小麦品种改良及系谱分析.北京:中国农业出版社,2003.
[2]盖红梅,王兰芬,游光霞,郝晨阳,董玉琛,张学勇.基于 SSR标记的小麦骨干亲本育种重要性研究.中国农业科学,2009,42(5):1503–1511.
[3]袁园园,王庆专,崔法,张景涛,杜斌,王洪刚.小麦骨干亲本碧蚂 4 号的基因组特异位点及其在衍生后代中的传递.作物学报,2010,36(1):9–16.
[4]韩俊,张连松,李静婷,石丽娟,解超杰,尤明山,杨作民,刘广田,孙其信,刘志勇.小麦骨干亲本“胜利麦/燕大 1817”杂交组合后代衍生品种遗传构成解析.作物学报,2009,35(8):1395–1404.
[5]李小军,徐鑫,刘伟华,李秀全,李立会.利用 SSR 标记探讨骨干亲本欧柔在衍生品种的遗传.中国农业科学,2009,42(10):3397–3404.
[6]王珊珊,李秀全,田纪春.利用 SSR 标记分析小麦骨干亲本“矮孟牛”及衍生品种(系) 的遗传多样性.分子植物育种,2007,5(4):485–490.
[7]李琼,王长有,刘新伦,高冬丽,吉万全.小偃6号及其衍生品种(系)遗传多样性的 SSR分析.麦类作物学报,2008,28(6):950–955.
[8]陈国跃,刘伟,何员江,苟璐璐,余马,陈时盛,魏育明,郑有良.小麦骨干亲本繁 6 条锈病成株抗性特异位点及其在衍生品种中的遗传解析.作物学报,2013,39(5):827–836.
[9]Li ZF,Zheng TC,He ZH,Li GQ,Xu SC,Li XP,Yang GY,Singh RP,Xia XC.Molecular tagging of stripe rust resistance geneYrZH84in Chinese wheat line Zhou 8425B.Theor Appl Genet,2006,112(6):1098–1103.
[10]Zhao XL,Zheng TC,Xia XC,He ZH,Liu DQ,Yang WX,Yin GH,Li ZF.Molecular mapping of leaf rust resistance geneLrZH84in Chinese wheat line Zhou 8425B.Theor Appl Genet,2008,117(7):1069–1075.
[11]盖红梅,李玉刚,王瑞英,李振清,王圣健,高峻岭,张学勇.鲁麦 14对山东新选育小麦品种的遗传贡献.作物学报,2012,38(6):954-961.
[12]何中虎,夏先春,陈新民,庄巧生.中国小麦育种进展与展望.作物学报,2011,37(2):202–215.
[13]程顺和,张伯桥,高德荣.小麦育种策略探讨.作物学报,2005,31(7):932–939.
[14]严济.五十年四川小麦育种研究的回顾与前瞻.四川农业大学学报,1999,17(1):108–113.
[15]李生荣.20世纪绵阳号小麦品种选育及应用.西南科技大学学报,2003,18(4):79–83.
[16]Chen PD,Qi LL,Zhou B,Zhang SZ,Liu DJ.Development and molecular cytogenetic analysis of wheat-Haynaldia 6VS·6AL translocation lines specifying resistance to powdery mildew.Theor Appl Genet,1995,91(6-7):1125–1128.
[17]Li GP,Chen PD,Zhang SZ,Wang X,He ZH,Zhang Y,Zhao H,Huang HY,Zhou XC.Effects of the 6VS·6AL translocation on agronomic traits and dough properties of wheat.Euphytica,2007,155(3):305–313.
[18]Yang WY,Liu DC,Li J,Zhang LQ,Wei HT,Hu XR,Zheng YL,He ZH,Zou YC.Synthetic hexaploid wheat and its utilization for wheat genetic improvement in China.J Genet Genomics,2009,36(9):539–546.
[19]李生荣,任勇,彭慧儒,杜小英,周强,李太军.小麦品种绵麦37选育与利用.科技成果管理与研究,2012,6:52–55.
[20]Liu TG,Peng YL,Zhang ZY.First detection of virulence inPuccinia striiformisf.sp.triticiin China to resistance genesYr24(=Yr26) present in wheat cultivar Chuanmai 42.Plant Dis,2010,94(9):1163.
[21]李生荣,杜小英,任勇,李太军,李茂有.小麦新品种绵麦367的选育.农业科技通讯,2012,(7):157–158.
[22]Somers DJ,Isaac P,Edwards K.A high-density microsatellite consensus map for bread wheat(Triticum aestivumL.).Theor Appl Genet,2004,109(6):1105–1114.
[23]张国华,高明刚,张桂芝,孙金杰,靳雪梅,王春阳,赵岩,李斯深.黄淮麦区小麦品种(系)产量性状与分子标记的关联分析.作物学报,2013,39(7):1187–1199.
[24]武玉国,吴承来,秦保平,王振林,黄玮,杨敏,尹燕枰.黄淮冬麦区175个小麦品种的遗传多样性及SSR 标记与株高和产量相关性状的关联分析.作物学报,2012,38(6):1018–1028.
[25]He XY,Singh PK,Duveiller E,Schlang N,Dreisigacker S,Sing RP.Identification and characterization of international Fusarium head blight screening nurseries of wheat at CIMMYT,Mexico.Eur J Plant Pathol,2013,136(1):123–134.
[26]Paillard S,Trotoux-Verplancke G,Perretant MR,Mohamadi F,Leconte M,Coëdel S,de Vallavieille-Pope C,Dedryver F.Durable resistance to stripe rust is due to three specific resistance genes in French bread wheat cultivar Apache.Theor Appl Genet,2012,125(5):955–965.
[27]李俊,魏会廷,胡晓蓉,李朝苏,汤永禄,刘登才,杨武云.川麦42中源于人工合成小麦的一个高产位点鉴定.作物学报,2011,37(2):255–262.
[28]Liu TG,Peng YL,Chen WQ,Zhang ZY.First detection of virulence inPuccinia striiformisf.sp.triticiin China to resistance genesYr24(=Yr26) present in wheat cultivar Chuanmai 42.Plant Dis,2010,94(9):1163.
[29]任勇,李生荣,李俊,周强,杜小英,李太军,杨武云,郑有良.两系杂交小麦恢复系 MR168抗条锈病基因遗传分析及分子标记定位.遗传,2011,33(11):1263–1270.
[30]张培禹,曾庆东,王琪琳,康振生,韩德俊.四川盆地小麦品种(系)抗条锈性鉴定与评价.麦类作物学报,2012,32(4):779–783.
[31]韩德俊,王琪琳,张立,魏国荣,曾庆东,赵杰,王晓杰,黄丽丽,康振生.“西北-华北-长江中下游”条锈病流行区系当前小麦品种(系)抗条锈病性评价.中国农业科学,2010,43(14):2889–2896.
[32]章振羽,姬红丽,沈丽,徐世昌,倪建英,彭云良.四川58 个小麦品种苗期抗条锈基因推导及成株期抗性表现.植物保护学报,2012,39(1):13–23.
[33]廖杰,魏会廷,李俊,杨玉敏,曾云超,彭正松,杨武云.川麦42遗传背景中人工合成小麦导入位点的SSR标记检测.作物学报,2007,33(5):703–707.
猜你喜欢
中国糖料(2023年4期)2023-11-01 09:34:46
湖北植保(2022年4期)2022-08-23 10:51:52
今日农业(2021年7期)2021-07-28 07:07:36
空间科学学报(2021年1期)2021-05-22 01:36:34
农民致富之友(2019年16期)2019-07-01 02:25:59
中国果业信息(2019年11期)2019-01-05 20:47:24
西藏农业科技(2018年4期)2018-04-25 06:39:20
环境保护与循环经济(2017年5期)2018-01-22 02:56:44
西南农业学报(2016年5期)2016-05-17 05:42:23
中国果菜(2016年9期)2016-03-01 01:28:44
