
CONCEPTUAL TROSPA IDENTIFIED FROM IXODES SINENSIS (ACARI:IXODIDAE) IN SOUTH CHINA*
2014-11-10 02:00:18WANGXinSUNYiCAOWuChunJIANGBaoGuiJIANaNIXueBingZHANGYuan
寄生虫与医学昆虫学报 2014年2期
WANG Xin SUN Yi CAO Wu-Chun JIANG Bao-Gui JIA Na NI Xue-Bing ZHANG Yuan
(State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing 100071, China)
Abstract TROSPA, a kind of tick receptor for outer surface protein A (OspA) of Lyme spirochete, was identified from specific vector species, Ixodes sinensis in South China. To improve our standing of the role of tick gene in tick borne pathogen infection/multiplication, larva of I. sinensis were allowed to feed on mice, two cDNA clones including I.sin-1 (258 bp) and I.sin-2 (462 bp) were identified as conceptual TROSPA. The putative amino acid sequence of I.sin-2 was 88.2%, 88.5% and 88.8% identical to that of I. scapularis, I. ricinus and I. persulcatus respectively, while the predicted amino acid of I.sin-1 shows only 46%, 46.1%, 45.6% to each respectively. This finding will lay the basis to analyze the interactions between I. sinensis and pathogens, and evaluate its potential application in blocking pathogen transmission from vector to host.
Key words Ixodes sinensis; Lyme spirochete; Surface protein; TROSPA
As a kind of haematophagous ectoparasites of wild and domestic animals and humans, Ixodid ticks have been proved to transmit various pathogens that cause zoonotic diseases threatening public health (de la Fuenteetal., 2008). Of them, members ofI.persulcatuscomplex are known as efficient vector species contributing to transmit spirochetes, rickettsia, virus and protozoan, which are considered as relevant to various tick borne human diseases (Laneetal., 1991). About 5 species were involved in the complex includingI.persulcatusin Russia, Far East, Asia,I.ricinusin Eurasia andI.scapularis,I.pacificusin America (Kolonin, 2009).Ixodessinensis, another member of the complex sharing closely morphological characters with other members, is known to distribute in South East Asia with high infestation rates in inhabitants and cattle, had been proved to be a competence vector species for human Lyme borreliosis in South China (Sunetal., 2003, 2009; Lietal., 2010).
Previous studies have demonstrated that tick gene expression is modified by in response to pathogen infection to ensure survival and development of both the pathogen and tick vector (Mulengaetal., 2003; Zivkovicetal., 2010). TROSPA,tick receptor of outer membrane surface protein A, was first described inI.scapularisas a receptor of OspA to facilitate Lyme spirochetesBorreliaburgorferito adhere to tick gut (Paletal., 2004). TROSPA mRNA levels fluctuate temporally linked to pathogen infection and transmission events. For example, TROSPA mRNA levels increased following spirochetes infection and decreased to tick engorgement in the Lyme transmission cycles (Paletal., 2006). Anti-TROSPA antibodies or gene knockdown reducedB.burgorferiadhere toI.scapularisgut in vivo (Paletal., 2004), and reduced protozoan parasiteBabesiabigeminaload inRhipicephalusannulatus(Antunesetal., 2012), and preventing efficient colonization of vector and subsequently reducing pathogen transmission to the mammalian host. TROSPA shows an enormous potential in understanding the molecular interactions at tick-pathogen interface and vaccine antigen to control pathogens infection in ticks (Paletal. 2004; Hoviusetal., 2007). For the purpose, the identification of TROSPA fromI.ricinus, andI.persulcatus(Konnaietal., 2012) were achieved in recent years, which raised us to hypothesize the molecular mechanism betweenI.sinensisand its’ pathogens. However, whether similar molecules and mechanism exist inI.sinensisis a barren area for speculation due to little sequence information and functional analysis available. In the present paper, we revealed two TROSPA homologous inI.sinensispossible involving in the transmission of pathogen and discussed the potential application in prevention of diseases transmitted by Ixodid ticks includingI.sinensisandI.persulcatus.
1 Materials and methods
1.1 Purification of I.sinensis RNA and RT-PCR
I.sinensislarva ticks were obtained from second laboratory colonies maintained at Department of Vector Biology and Control, Beijing Institute of Microbiology and Epidemiology and maintained in conventional conditions. The larva were allowed to feed on the mice for three days to semi-engorged and removed immediately to grind under liquid nitrogen. RNA samples were extracted and purified according to manufacturer′s instruction of RNeasy mini kit (QIAGEN Co. Ltd.). An aliquot of the total RNA(1.6 μg) was reverse-transcribed using the oligo-dT primer (20 pmol) in a total reaction volume of 20 μL according to manufacturer’s instructions of First-Strand Synthesis System (Invtrogen). The specific primers TROSPAF: 5′-ATGGCGGCTATGGAGGCTAT-3′ and TROSPAR: 5′-TCAACTTCCAGCGGCGCTCT-3′ were designed according to the existed TROSPA CDS ofI.scapularis(AY189148),I.ricinus(EU384705),I.persulcatus(BAK09229) deposited in GenBank to amplify the TROSPA homologues in the larva ofI.sinensis. The reaction mixture for PCR contains 2 mmol/L Mg2+, 0.2 mmol/L of each deoxynucleoside triphosphates (dNTPs), 1.25 U DNA polymerase(Takara), 20 umol/L of each primer, 3 ng cDNA in final volume of 50 μL. The Cycling conditions for PCR were 3 min pre-denaturation at 95℃, 12 cycles of denaturation at 94℃ for 30 s, annealing at 55.5℃for 40 s, extension at 72℃for 1 min, following with 20 cycles of denaturation at 94℃ for 30 s, annealing at 52℃for 40 s, extension at 72℃ for 1 min. Final extension at 72℃for 10 min followed by a hold step at 4℃, the amplified products were visualized under UV light with a ChampGel-3200 Photographic system.
1.2 Sequencing and Analysis
PCR amplification products were purified according to the manufacturer’s instruction of QIAquick Gel Extraction Kit (QIAGEN) and then linked into the pEASY-T1 Simple Cloning Vector (TransGen) and transformed into competentE.colistrain tans5α (Transgen). The purified positive plasmids were submit for sequencing by a dideoxybucleotide cycle sequencing method with an automated DNA sequencer (ABI 3730) in Sangon Bio-technique Company in Shanghai. The sequences were analyzed using CLC Genomics Workbench software (CLC bio, Inc.). After diminishing the clone vector sequences contamination, the alignment of 5 TROSPA sequences were performed using the ClustalW2 program from EMBL(European Molecular Biology Laboratory). In different species, the nucleotide and amino acid similarities of TROSPA are calculated by needle (v6.0.1) in WebLab (Liuetal., 2009).
2 Results
As expected, two potential amplicons were obtained from semi-engorged larva ofI.sinensisafter RT-PCR. One fragment with 258 bp was namedI.sin-1 and the other (I.sin-2) with 462 bp. The sequences of the two amplicons similar to TROSPA and been submitted to EMBL with accession number HE820736.1 and HE820737.1 respectively. TheI.sinensisTROSPA homologous showed high homology to TROSPA discovered in other member ofI.persulcatuscomplex (Fig. 1). The putative protein sequence ofI.sin-2 was 88.2%, 88.5% and 88.8% identical to that ofI.scapularis,I.ricinusandI.persulcatusrespectively. While the predicted protein ofI.sin-1 shows only 51.6% identical toI.sin-2 and 46%, 46.1%, 45.6% similarity toI.scapularis,I.ricinusandI.persulcatusrespectively. Alignment of the putative different full length TROSPA using the web-based Clustal W program in EMBL, revealed conserved domain in these protein (Fig. 2). Due to the available sequences shortage ofI.sin-1 in N-terminus,I.sin-2 fromI.sinensisshares more conserved sites with other 3 alignment sequences fromI.scapularis,I.ricinusandI.persulcatusthanI.sin-1.
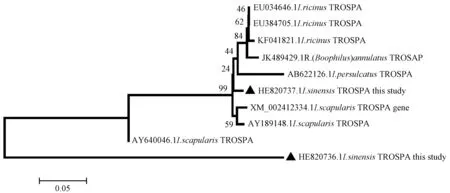
Fig. 1 Unrooted phylogram inferred using Neighbor-Joining method based on deduced amino acid sequences of TROSPA homologs of tickThe tree is drawn to scale with branch length in the same unit as those of the evolutionary distance used to infer the phylogeny tree.
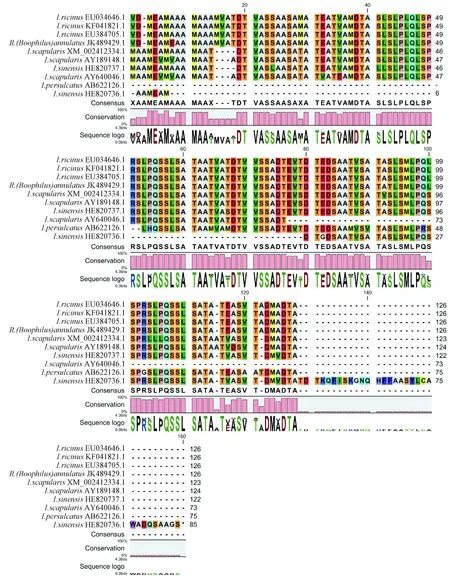
Fig. 2 Alignment of deduced amino acid sequences of TROSPA homologs of tick with Clustal W. Asterisks indicate the consistent amino acids.
Both the two sequences have several potential posttranslational sites of modification, including O-beta-GlcNAc domain, phosphorylation sites, however, onlyI.sin-2 with a putative transmembrane helix (Fig. 3) based on the web-software in ExPASy. For details,I.sin-1 is predicted with 13 O-beta-GlcNAc sites, 8 phosphorylation sites (threshold>0.5), whileI.sin-2 is predicted with 22 O-beta-GlcNAc sites, 10 phosphorylation sites (threshold>0.5), and a transmembrane helix (between residues 9-33) by Temped Prediction.

Fig. 3 Amino acid sequences of cDNA encoding TROSPA (HE820737.1) of I. sinensisThe putative O-glycosylation sites and transmembrane domain are underlining and shading respectively, as well as phosphorylated sites marked by asterisks.
3 Discussion
TROSPA, a precise set of ligand-receptors responsible for successful persistence of human pathogens in nature, has conservative domain in different vector species (Konnaietal., 2012). The putative TROSPA fromI.sinensisshare the similar conservative domains with that ofI.scapularis,I.ricinusandI.persulcatus, which indicates the similar molecular interactions between tick borne pathogens such asBorreliaburgdorferiandI.sinensis. Of the two homologous obtained, whetherI.sin-1 orI.sin-2 fromI.sinensisadhered more effectively to OspA ofBorreliaburgdorferiand how these homologous produced might be partial explained by the following reasons. Firstly, bothI.sin-1 andI.sin-2 might be derived from the same gene ofI.sinensis, and transcribed as the same precursor mRNA. However, the two homologuesI.sin-1,I.sin-2 result from the different post-transcription process; in particular, the shorter one is a splice variant of the longer one. Secondly,I.sin-1andI.sin-2 may originate fromparalogousgenes which responsible for the difference between their coding sequences. Referring to their functions,I.sin-2 may be able to interact with the OspA like its homologues inI.scapulariswhile theI.sin-1 may have a higher or lower affinity to OspA or even lose the ability or serve an unknown different bioprocess for its primary structural absence of 204 nucleotides and only one mutation of A→G in 5’ -terminus. Another possibility about their biological function is thatI.sin-1 can be have the similar affinity of interacting with spirochete or has other natural functions as well asI.sin-2, because OspA is presumed to bound to high molecular mass proteins, or protein receptor complex (Paletal., 2000), which suggests TROSPA and its homologous involves in a more complicated process. In this respect, TROSPA-like proteins may be a part of protein complex to perform its function. For the reason of unclear TROSPA binding sites with OspA, the interaction between TROSPA and pathogens including Lyme spirochetes remained to be investigated.
Attempts to explore the complex mechanisms by which tick borne pathogens persists in a specific enzootic cycle and develop vaccines targeting on pathogen transmission, had been undertaken for many years (de la Fuenteetal., 2011). Due to the known role of TROSPA in the infection of Lyme spirochetes in the members ofI.persulcatuscomplex and protozoan parasites inR.annualus, TROSPA vaccination could contribute greatly to reduce pathogens colonization and survival in the tick gut, thus might impair pathogen transmission to subsequent hosts (Fikrigetal.1990; Fikrigetal.1991; Tsaoetal.,2001; Tsaoetal.,2004; Hoviusetal. 2007; Antunesetal., 2012). The newly finding conceptual TROSPA inI.sinensismight throw a light in the prospects of vaccination with TROSPA to block pathogen transmission in the future. However, detail studies to evaluate the vaccination efficacy of TROSPA and feasible vaccine delivery systems need to be undertaken.
AcknowledgementsWe are grateful to Lindsay Morton at the University of South Florida for reviewing the manuscript. This work was supported by the National Nature Foundation of P.R. China (No. 30872196; No. 81271878).
