
角果藜的结实格局及地下散布单位的入土机制
2014-11-07 01:46王喜勇高蕊周晓青魏岩
草业学报 2014年2期
王喜勇,高蕊,周晓青,魏岩
(1.新疆农业大学草业与环境科学学院 新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052; 2.中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011;3.辽宁丹东市青山保护局,辽宁 丹东 118000; 4.陕西宝鸡石油中学生物组,陕西 宝鸡 721001)
植物物候是植物为了适应气候条件的节律性变化而形成的与此相应的植物发育节律[1],是植物在进化过程中遗传下来的为生存不断适应环境的结果[2]。植物在不同物候期的物候特点,集中体现了植物体在不同环境条件下所采用的适应策略。
在异质性环境中,种子多型性被认为是对时空异质环境的一种适应[3]。地上地下结果性(amphicarpy) 是种子多型性中的特殊一种,是指同一植株产生不同形态的地上果实(aerial fruit)和地下果实(subterranean fruit)的现象[4-5]。其中,地下结果性又分为两类,即植物体地下开花地下结果和基部开花地下结果[6]。关于基部开花地下结果植物地下果实形成机制的研究已较为深入,Barker[6]提出了主动地下结果性和被动地下结果性。主动地下结果性指植物体通过一定的生长方式或向性方式形成的小梗或小柄的主动方式使果实入土;而被动地下结果性指植物体借助风或水的作用将覆盖植物体或果实的土壤沉积下来的被动方式使果实入土。地上地下结果性现象作为植物对生态环境的一种独特适应方式,在研究植物的生态适应机制、生活史对策进化方面具有重要价值。
角果藜(Ceratocarpusarenarius)隶属藜科角果藜属,为一年生草本植物,广泛分布于亚洲中部的荒漠和沙漠中[7]。在我国仅生长在准噶尔盆地,并能在准噶尔盆地的局部地区形成层片或群聚[8]。角果藜是首次在藜科中发现的具有地上地下结果性的物种[9]。那么,角果藜的地下散布单位是如何入土的?其入土的时空特点如何?本文通过对角果藜的物候、地上地下结实格局及地下散布单位入土方式的研究,揭示地下散布单位的入土机制,探讨角果藜适应荒漠环境的生活史特征,对于深入了解荒漠植物的进化规律及其对特定环境的生态适应机制具有重要的意义。
1 材料与方法
1.1 研究材料与研究地概况
角果藜为藜科角果藜属一年生草本植物,3月初萌发,花期4-8月,花单性,雌雄同株,具有地上地下结果特性。角果藜的果实为胞果,由雌花发育形成。雌花无花被具2枚苞片,受精后,2苞片完全愈合将果实包被。本文将果实及包被其外面的苞片称为散布单位(dispersal units)。地上散布单位倒三角形,地下散布单位倒卵形[9]。
研究区位于新疆准噶尔盆地南缘低山砾质荒漠的自然种群(43°48′ N, 87°40′ E, 750~850 m),土壤为细砾质灰棕荒漠土,植被类型为小半灌木荒漠小蓬群系,天然植被以小蓬(Nanophytonerinaceum)、散枝猪毛菜(Salsolabrachiata)、紫翅猪毛菜(Salsolaaffinis)、角果藜为主,盖度20%~30%。该地区属中温带荒漠区,春秋季较湿润,夏季干旱,热量充足,冬季漫长而寒冷。≥5℃初日温度出现在3月下旬,≥10℃初日温度出现在4月下旬;高温出现在7月,平均最高气温为29.7℃[10]。
1.2 研究方法
1.2.1物候观测 2007—2008连续2年对角果藜自然种群的物候进行了观察。物候参照宛敏渭与刘秀珍[11]的物候观测法,在萌动期至开花期每隔2 d观察1次,果实形成期至成熟期每隔7 d观察1次。
1.2.2分支方式与结实格局的观察 在果熟期观察植株的分支方式、散布单位在植株上的空间位置,并用AutoCAD软件绘制角果藜的分枝方式和散布单位空间分布图。
1.2.3地下果实入土方式的测定 于2007年4月下旬-10月下旬每隔30 d随机采集20株角果藜,测量地下散布单位顶端及其着生基部离地距离(20个重复,取平均值)。同时,在Motic B5型生物显微镜下测量地下散布单位与其主茎间的角度(20个重复,取平均值),采用SPSS 11.0软件对夹角与生殖生长天数间进行相关性分析,确定地下散布单位的入土方式。
2 结果与分析
2.1 物候特征
角果藜的种子萌发早,于3月中上旬冰雪未完全融化时就开始萌发;3月中上旬子叶出现到4月中上旬枯落,子叶期持续时间长(2007年29 d;2008年28 d);完全营养生长期较短,从萌发到现蕾(2007年36 d,2008年29 d),仅占整个生活周期的约1/5,之后,营养生长和生殖生长并存;生殖时间早,萌发后约30 d现蕾,生殖期持续时间相对较长(2007年179 d,2008年173 d),约占整个生活周期的4/5以上(表1,图1)。
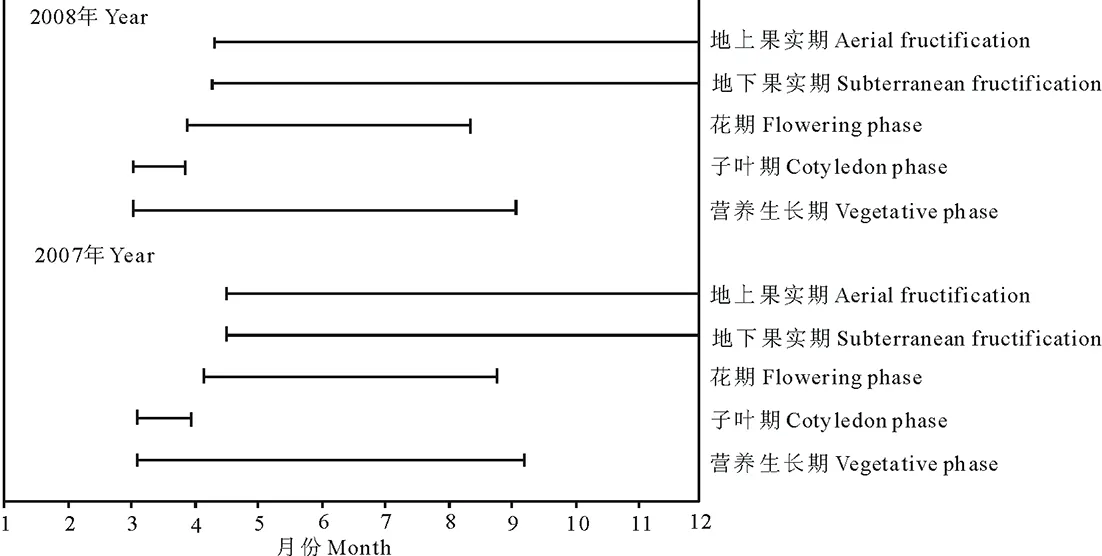
图1 角果藜的物候图(2007,2008年)Fig.1 The phenological chart of C. arenarius in 2007 and 2008
角果藜开花后,苞片迅速愈合并增大形成倒三角形或倒卵形的散布结构,而果实则“停滞”发育,至8月下旬果实开始膨大,9月末以“集中、快速”的模式发育成熟。
角果藜2007,2008年的物候见表1。
2.2 角果藜的分枝方式及结实格局
观察发现,角果藜为三歧聚伞状分枝方式,枝顶形成聚头状雄花序后,停止发育,其下部叶腋依次形成3个分枝,植物体共有5~6级分枝形成球状。同时,主茎第1轮雄蕊着生的节膨大并形成离层。角果藜地上散布单位为倒三角形,多数,单生于叶腋,呈螺旋上升状排列于每个分枝上。地下散布单位为倒卵形,2个,位于主茎第1节处,仅为第1对真叶叶腋处的1对雌花受粉后发育形成(图2)。
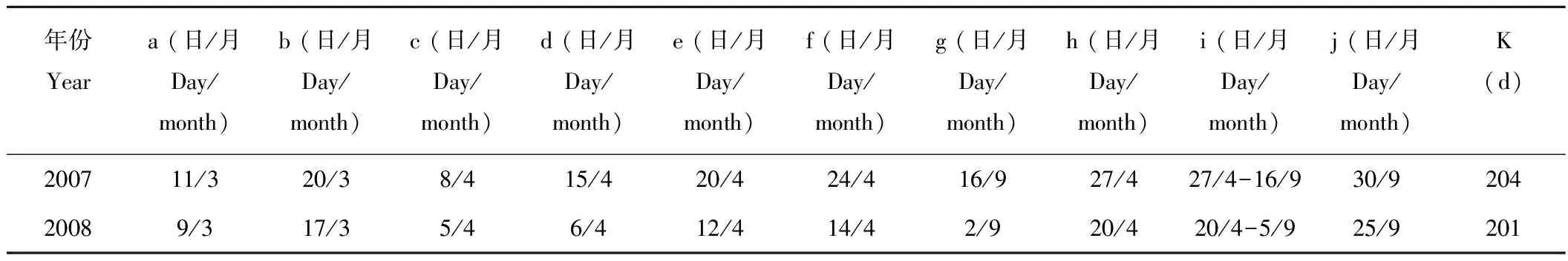
表1 角果藜的物候特点(2007,2008年)Table 1 Phenological characteristics of C. arenarius in 2007 and 2008
a,萌动 Germination;b,展叶 Seedling;c,子叶败落 Cotyledon decline;d,现蕾 Flower bud appearing;e, 始花 First flower appearing; f,盛花 Full blooming;g,末花 Flowering termination;h,地下果形成Underground fruit appearing;i,地上果形成Aboveground fruit appearing;j,果熟 Fruit maturing;k,生活周期Life cycle.
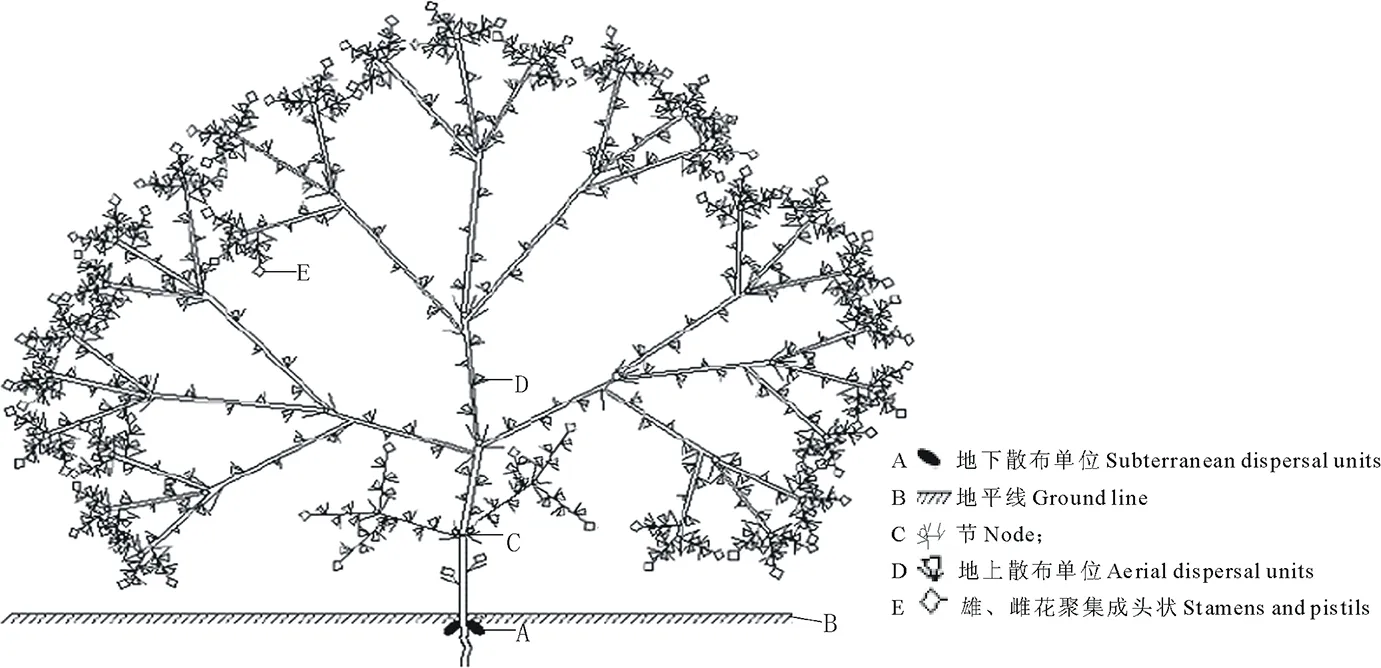
图2 角果藜结实格局Fig.2 Branching mode and fruit-set patterns of C. arenarius
2.3 地下散布单位的入土方式
地下散布单位是由第1节对生叶腋处的2朵雌花发育而来。4月下旬,2朵雌花地上开花授粉,雌花外2枚苞片于5月中上旬完全愈合形成散布单位,散布单位于6月中下旬入土继续发育形成地下散布单位。从图3和表2可以看出,地下散布单位与角果藜主茎间的夹角随时间变化逐渐增大,由初形成时的(32.05±0.32)°(图3-1)达到生殖生长135 d后(134.01±0.60)°(图3-3)的最大值。相关性分析显示,地下散布单位与主茎间夹角与生殖生长天数之间呈显著正相关关系(Person相关系数为0.920,P<0.01);表3显示,随地下散布单位的翻转与增大,其顶端离地表距离逐渐减小,在6月末、7月初地下散布单位顶端进入土壤。同时,在风沙及雨水带来的土表覆盖物掩埋作用下,地下散布单位完全进入土壤。因此,地下散布单位在主动性的“翻转”和被动性埋藏的双重作用下,顺利完成入土。

图3 角果藜地下散布单位翻转过程Fig.3 Reversing process of subterranean dispersal units of C. arenarius

项目Item生殖生长时间 Time of procreation growth 15 d (4月 April)30 d (5月 May)45 d (5月 May)60 d (6月 June)75 d (6月 June)90 d (7月 July)夹角 Angle (°)32.05±0.3254.62±1.04106.24±2.02133.17±0.45134.01±0.60133.68±0.88
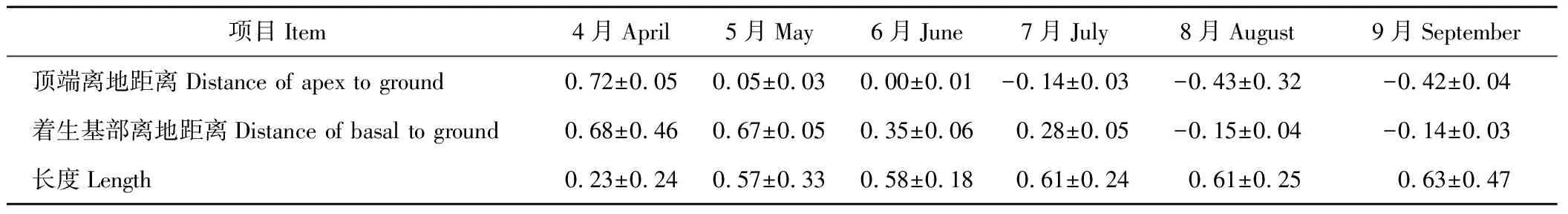
表3 地下散布单位着生基部、顶端离地距离及其长度Table 3 The distance of apex and basal to ground and length of subterranean dispersal units for C. arenarius (mean±SE) cm
3 讨论
不同的物候特征对植物的生殖成功有重要影响[12-14]。在新疆准噶尔盆地3-4月的早春,气温逐渐升高,≥5℃初日温度出现在3月下旬,≥10℃初日温度出现在4月下旬,实践表明5~6℃是植物发育的基础温度[15-16],同时积雪融化,浅层土壤形成充足的水分,整个生活史内早春是土壤含水量最高的时期,这些条件对植物的生长发育极为有利。角果藜利用有利的环境条件,提早萌动从而保证较高的资源利用效率。子叶是种子植物胚的组成部分之一,为贮藏养料或幼苗时期进行同化作用的器官,所以子叶在种子萌发成幼苗的初期,其作用十分重要。角果藜的子叶持续长约30 d,不但将其储藏的养料最大程度的提供给幼苗,同时绿色子叶的光合产物也是幼苗早期主要的能量来源,保证幼苗的成功生长。角果藜完全营养生长时间短,为有性生殖积累基本能量;生殖时间早,在幼态下进行生殖,营养生长和生殖生长同时进行,持续时间较长,避免了夏季干旱高温来临,植物尚未结实而导致种群灭绝。繁殖时间是由遗传决定的,以适应不同的环境压力[17-18],如有较大的死亡危险,推迟种子产生就可能导致繁殖的彻底失败。6月夏季来临,温度逐渐升高,土壤含水量下降,环境条件对植物的生长发育极为不利,短命植物大多果实已成熟并完成了其生活史[19]。而非短命植物角果藜仍处于营养生长和生殖生长并存期,通过观察发现,角果藜开花后,花外苞片愈合,但包被其内的果实处于“停滞发育”期,直至8月下旬恶劣环境逐渐好转时果实才以“集中、快速”的模式发育成熟。这种繁殖策略,既能使植物体实现最大的繁殖产量,又能使其逃避准噶尔荒漠夏季干热气候对其种子繁殖所带来得高死亡风险,最终达到生殖成功。
研究表明种子的异型性符合两面下注(bet-hedging) 策略,主要表现在通过产生不同休眠程度的种子和不同空间扩散程度的种子,在时间和空间上分散风险,从而适应环境时空异质性[20]。在数量上,角果藜地上散布单位多数,地下散布单位仅为2个,植物通过感受环境胁迫,产生更多的容易扩散的种子,使植物的后代能够逃离不利的环境,寻找比较适宜的环境[21];在休眠程度上,地上散布单位比地下散布单位的休眠程度浅,易于萌发,而休眠程度较深的地下散布单位则可以形成持久地下种子库;在扩散方式上,地上散布单位随母体“风滚”散布拓展后代分布范围,地下散布单位保留在母体附近不易扩散[9];在形成机制上,角果藜形成散布单位的花具有专一性,即只有第1对真叶叶腋处的1对雌花受粉后发育成地下果实,而其他雌花(两种雌花无形态差异)受粉后均形成地上果实,这种专一性使植物体开花后首先确保地下散布单位的形成;在散布单位的分布格局上,地下散布单位近地表形成利于入土并被沙土埋藏,并且保留在母体附近进行原位萌发,对地下果实的投资为种群在原分布区的繁衍提供了可能,使其能够在恶劣的荒漠环境达到最大的繁殖成功。地上散布单位着生于三歧聚伞状分枝形成的球型植株上,利于其风滚散布时的散布效率。角果藜的地上地下两种散布单位在数量、萌发程度、扩散方式[9]、形成机制、结实格局上的差异性都充分体现了这种两面下注的适应策略。
在基部开花地下结果植物地下果实入土机制中,地下果实被动性的入土行为主要是借助风沙、雨水等外力因素来实现,主动性入土机制主要是通过特化结构来实现的[6]。豆科中的3个种(Trifoliumsuberraneum,Voandzeiasubterranean,Derstingiellageocarpa)是通过某些向性生长方式形成小梗或小柄的方式完成地下果实的主动入土[22]。单子叶植物(Narcissus,Eucomispallidiflora,Eucomisautumnalis)和双子叶植物Catanancheluten(Compositae)通过收缩根使茎基部的果实入土[23-25]。Wilson 和 Honey[26]通过解剖实验研究发现收缩根通过收缩区中内部皮层薄壁组织细胞的纵向缩短和垂直扩展,同时还有包括皮层外部细胞、外皮层和表皮在内的外部组织越过收缩区表面被推入巨大的褶皱中进行收缩。本文研究结果表明,角果藜是通过主动性和被动性的双重作用使地下散布单位入土的。一方面,不仅可借助风沙等外力因素来埋藏地下散布单位。同时,果实主动性的动态“翻转”模式是更为节省且有效的入土策略。作为荒漠植物,生存环境极其恶劣,而特化组织的形成需要更多的能量分配。因此角果藜选择这种特殊的主动入土模式即保证了果实的顺利入土,又节省了能量分配,是其对荒漠环境长期适应的结果。
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
农业工程学报(2021年23期)2021-03-11
安徽科技学院学报(2018年5期)2019-01-18
农业知识(2018年18期)2018-05-28
种子(2017年12期)2018-01-17
西南农业学报(2017年5期)2017-06-23
创新作文(3-4年级)(2016年9期)2016-12-22
河北林业科技(2016年5期)2016-11-08
农家顾问(2009年11期)2009-11-28
