
放牧强度对草甸植物小叶章及土壤化学计量比的影响
2014-11-07 01:54张婷翁月姚凤娇史印涛崔国文胡国富
草业学报 2014年2期
张婷,翁月,姚凤娇,史印涛,崔国文,胡国富*
(1.东北农业大学生命科学学院,黑龙江 哈尔滨 150030;2.东北农业大学动物科学技术学院,黑龙江 哈尔滨 150030)
草原是我国面积最大的内陆生态系统,其健康稳定的发展对于牧区的经济发展及生态环境的安全都起着非常重要的作用,但近年来中国天然草原有90%发生了不同程度的退化[1]。放牧主要通过动物的采食、践踏以及排泄物归还来影响草地土壤的养分组成,植物自身通过改变养分的利用策略来适应外界环境的变化。草原生态系统中,植物的叶片和茎是动物采食和践踏的主要对象,对放牧的响应最为敏感[2]。而且叶和茎作为植物的重要器官,对植物的生长、发育和生活史的完成都有极其重要的意义。家畜采食不仅能够刺激牧草的生长,还可以促进地上N、P元素向幼嫩器官重新分配,且N的转移量大于P[3],从而使放牧草地地上生物量和N、P含量增加[4],放牧会改变植物地上部分的形态和功能,C同化和积累能力也会发生变化。另外放牧过程中动物的践踏会改变土壤物理状况,影响根系对土壤营养的吸收[3],同时放牧后的凋落物、动物的粪便等会改变土壤营养状况,导致土壤的理化性状发生变化。同时,土壤营养状况、营养吸收能力和同化能力的变化都最终体现到植物C、N和P含量及计量比的差异上。李香真和陈佐忠[5]1998年研究显示,在一定的放牧强度下,植物和根中的C/N比降低,Frank[6]在2008年研究了美国黄石国家公园区5个不同地形的放牧草地,发现放牧增加了N浓度而对P浓度无明显影响,从而提高了枝条N/P值。
氮、磷是植物生长必需的营养元素,也是生态系统中最常见的限制性元素,两者在植物体内存在着功能上的联系,植物叶片的N、P含量能够反映土壤N、P的有效性,所以,研究植物叶片的N、P含量及其比例关系能更好地认识和了解生态化学量学以及全球的碳循环模拟和宏观生态学的变化情况。
1 材料与方法
1.1 试验地自然概况
本试验地位于黑龙江省双鸭山市宝清县东升湿地。地处45°47′8″~46°35′55″ N,131°14′16″~133°29′48″ E。海拔300~400 m, 属寒温带大陆性季风气候, 年均气温2.3~2.4℃, 年均降水量551.5 mm, 主要集中在7-8月。≥0℃年积温2500~2700℃,无霜期143.3 d,年均风速2.5 m/s。植被类型为以小叶章(Deyeuxiaangustifolia)为优势种的草甸草原,土壤肥沃,类型为草甸土,呈微酸性[6]。
1.2 试验地设计
试验地设置4个放牧区,即轻牧区(0.99 AU/hm2·月,LG)、中牧区(1.65 AU/hm2·月,MG)、重牧区(2.30 AU/hm2·月,HG)和对照区(0 AU/hm2·月,CK),每个样区设置3个重复(共12个小区),如表1所示。本放牧试验时间为2012年5月15日至2012年9月15日, 家畜为当地肉牛, 出牧时间为早6:00-晚18:00,放牧完后赶出小区,试验条件和设施完善。
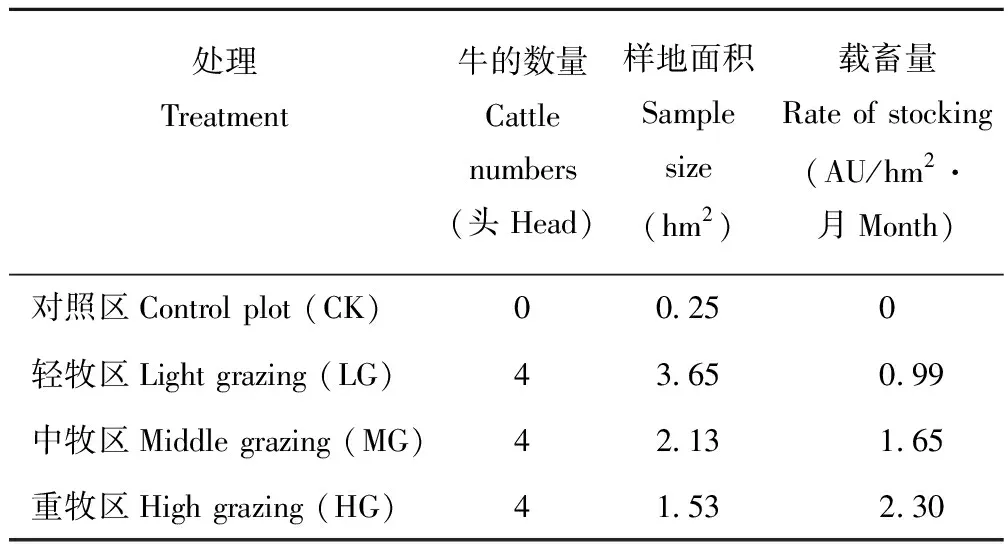
表1 试验样地设计Table 1 Plot design
AU:牛头Cattle number.
1.3 试验地取样方法
2012年5月至9月,每月中旬在每个试验小区随机选取6个0.5 m×0.5 m样方,将样方内的所有小叶章,用剪刀齐地面剪下,将立枯物与植物活体分开,分别装入密封袋中。将植物样品带回实验室后,将采集样从每个小区的每个样方中随机选取30株小叶章植物活体,将其茎、叶分开,分别称其鲜重,在105℃下杀青30 min,置于65℃下烘至恒重[8]。
在植被观测样方内,在小叶章成簇生长的地方,采用五点取样法,用土钻(内径为5.0 cm)分别取0~10 cm, 10~20 cm的土壤样品,将每个样方内5个点的同一土层混合,装入封口袋中。将土壤样品分成两部分:其一,将每个封口袋中土壤分出一小部分(无根系),经风干后粉碎并过筛(0.25 mm),放入封口袋中待测;其二,将剩余带根系的土壤放入孔径大小为0.28 mm的尼龙网袋内,置于水中浸泡后,挑拣出所有杂物,取出根系并用清水冲洗干净。将得到的根系在85℃下烘至恒重。
将烘干后的小叶章全株、根、茎、叶及立枯物粉碎后过0.25 mm筛后,留样分析。
1.4 样品的分析测定
植物与土壤的全C含量的测定采用重铬酸钾氧化-外加热法、全N含量的测定采用凯氏定氮法;植物样品全磷的含量用浓H2SO4-H2O2消煮,土壤样品采用NaOH熔融法,并用钼锑抗比色法测定。
1.5 数据分析
使用EXCEL软件进行初步处理。用SAS 9.1分析软件进行统计分析,结果以均值±标准差形式表示。
2 结果与分析
2.1 放牧对小叶章全株养分和化学计量比的影响
由表2可以看出,全株的全碳含量:在整个放牧季,全碳含量变化规律不明显,中牧区全株全碳含量呈逐渐上升的趋势,而轻牧、重牧及对照区均呈先下降后上升趋势。受放牧强度的影响,6和9月份,各牧区小叶章全株全碳含量差异不显著(P>0.05);7月份,各小区小叶章全株全碳含量随放牧强度的增加而升高;8月份,中牧和重牧区均显著高于轻牧区、对照区(P<0.05),以中牧区为最高。小叶章全株的全氮含量:各小区均在6月份为最大值,之后在7-8月份均有降低,在9月份又有小幅度的回升。受放牧的影响,6-7月份,随着放牧强度的增加,小叶章全株的全氮含量均呈增加趋势,在8-9月份重牧区的含氮量有所降低,且均以中牧区小叶章全株的全氮含量为最大。全株的全磷含量:在整个放牧季,对照区(除8月份)和重牧区全磷含量逐渐升高,轻牧区和中牧区则先升高后降低,且分别在7和8月份达到最大值。每个月份,各小区的全磷含量均随着放牧强度的增加,呈上升趋势。
在整个放牧季,各放牧区植物全株的C/N,均在8月份达到最大值。受放牧的影响,6-8月份随着放牧强度的增加C/N先降低后升高,且以中牧区为最小值,但各小区的差异均不显著;9月份随着放牧强度的增加C/N呈逐渐下降的趋势。小叶章全株的N/P值范围在3.96~10.61之间,6-9月份随着放牧强度的增加N/P先升高后降低。
2.2 放牧对小叶章茎的养分和化学计量比的影响
由表3可以看出,小叶章茎的全碳含量季节变化规律不明显。受放牧的影响,6和8月份,各小区小叶章茎全碳含量差异显著,大小分别依次为LG>CK>HG>MG和LG>HG>CK>MG;7月份,轻牧和重牧区显著高于中牧区、对照区(P<0.05),以重牧区小叶章茎的全碳最高;9月份,重牧区和对照区均显著高于轻牧、中牧区(P<0.05)。在整个放牧季,茎全氮含量的规律性不明显。受放牧影响,6和9月份对照区茎的含氮量显著高于放牧区(P<0.05),7月份随着放牧强度的增加而升高,8月份在轻牧区茎的全氮含量显著高于其他放牧小区(P<0.05)。茎的全磷含量受放牧的影响,7-9月份,轻牧区茎的全磷含量显著高于其他牧区(P<0.05),6月份对照区显著高于其他牧区(P<0.05)。
小叶章茎的C/N化学计量比变化没有全株C/N的规律性强,6-7月份茎的C/N均在轻牧区达到最高值,且显著高于其他小区(P<0.05),8月份,随着放牧强度的增加,茎的C/N呈先降低后升高的趋势;9月份,随着放牧强度的增加C/N逐渐升高,但各小区之间差异并不显著(P>0.05)。茎的N/P值总体变化规律不明显。
2.3 放牧对小叶章叶的养分和化学计量比的影响
由表4可以看出,叶的全碳含量:在整个放牧季,轻牧区叶的全碳含量逐渐升高,其他牧区的变化规律不明显。受放牧的影响,6月份各放牧区差异不显著,7月份,对照区叶的全碳含量显著高于其他放牧区(P<0.05),8-9月份均呈先升高后降低的趋势。叶的全氮含量:受放牧影响,7-8月份,叶的全氮含量随着放牧强度的增加逐渐升高,但各牧区间差异不显著,9月份(除中牧区外)也有相同的变化规律。叶的全磷含量:受放牧影响,随着放牧强度的增加,各月份叶的全磷含量均呈先降低后升高的趋势,且以重牧区为最大值,总体来看放牧能够增加叶片的全磷含量。
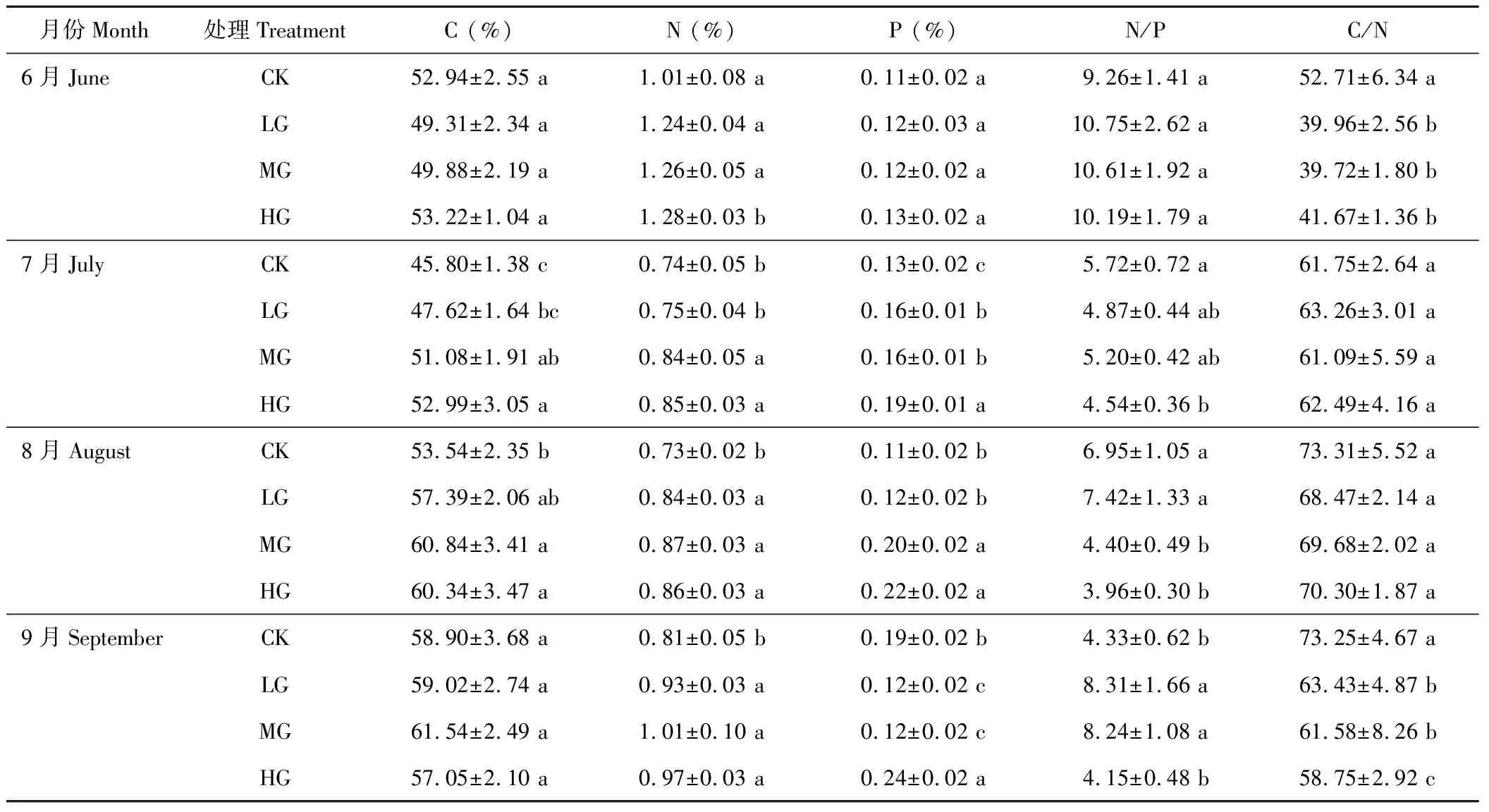
表2 小叶章全株C、N、P化学计量比的变化Table 2 Change of ecological stoichiometry of D. angustifolia
注:表中数据为平均值,多重比较仅限于同月份之间,同列中不同小写字母表示放牧强度间差异显著(P<0.05),下同。
Note:Data are means, multiple comparison is limited in a same month, and different lower case letters in the same column are significantly different among the treatments at the 0.05 level, the same below.
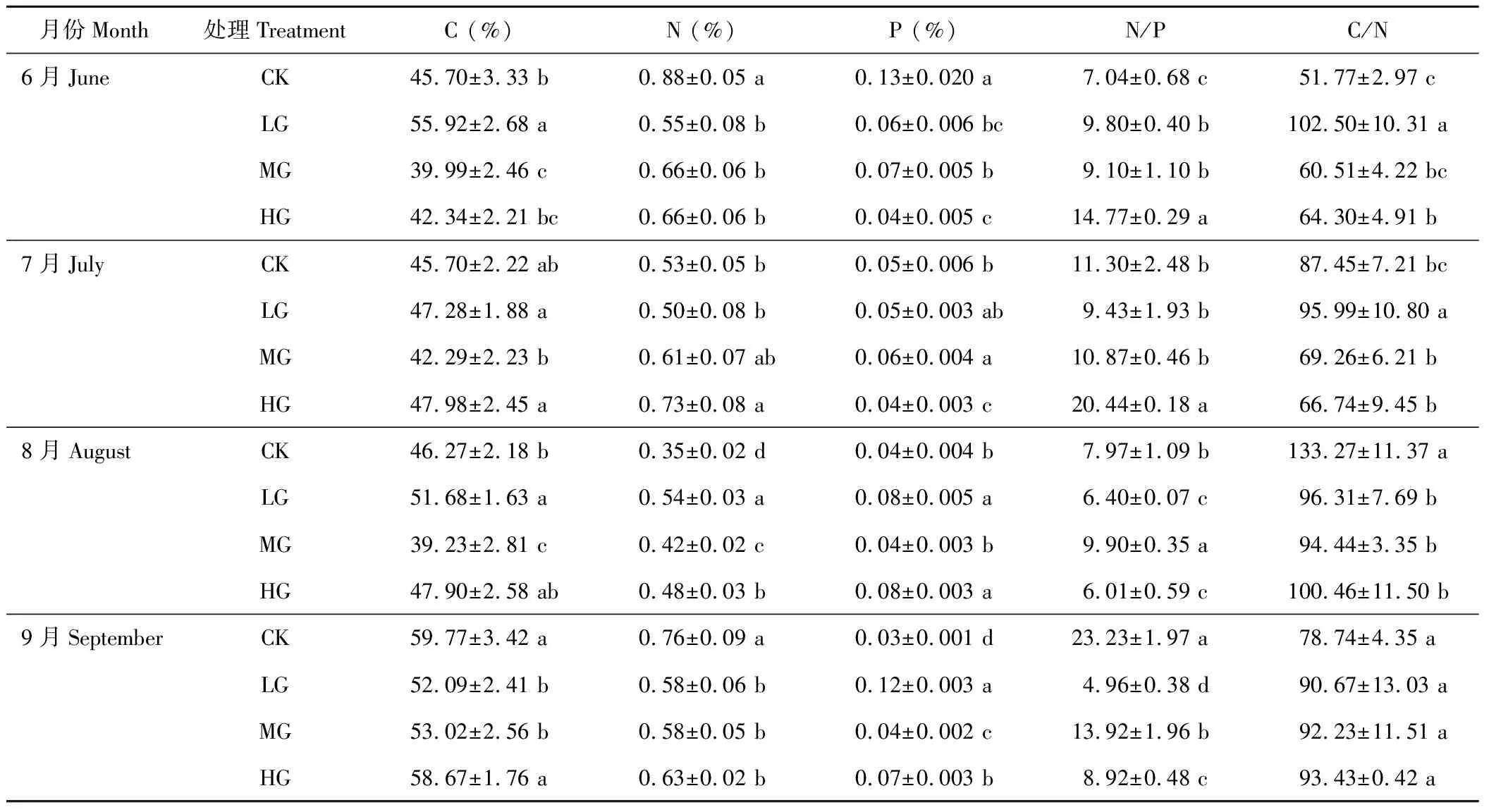
表3 小叶章茎的C、N、P化学计量比的变化Table 3 Change of ecological stoichiometry of D. angustifolia stem
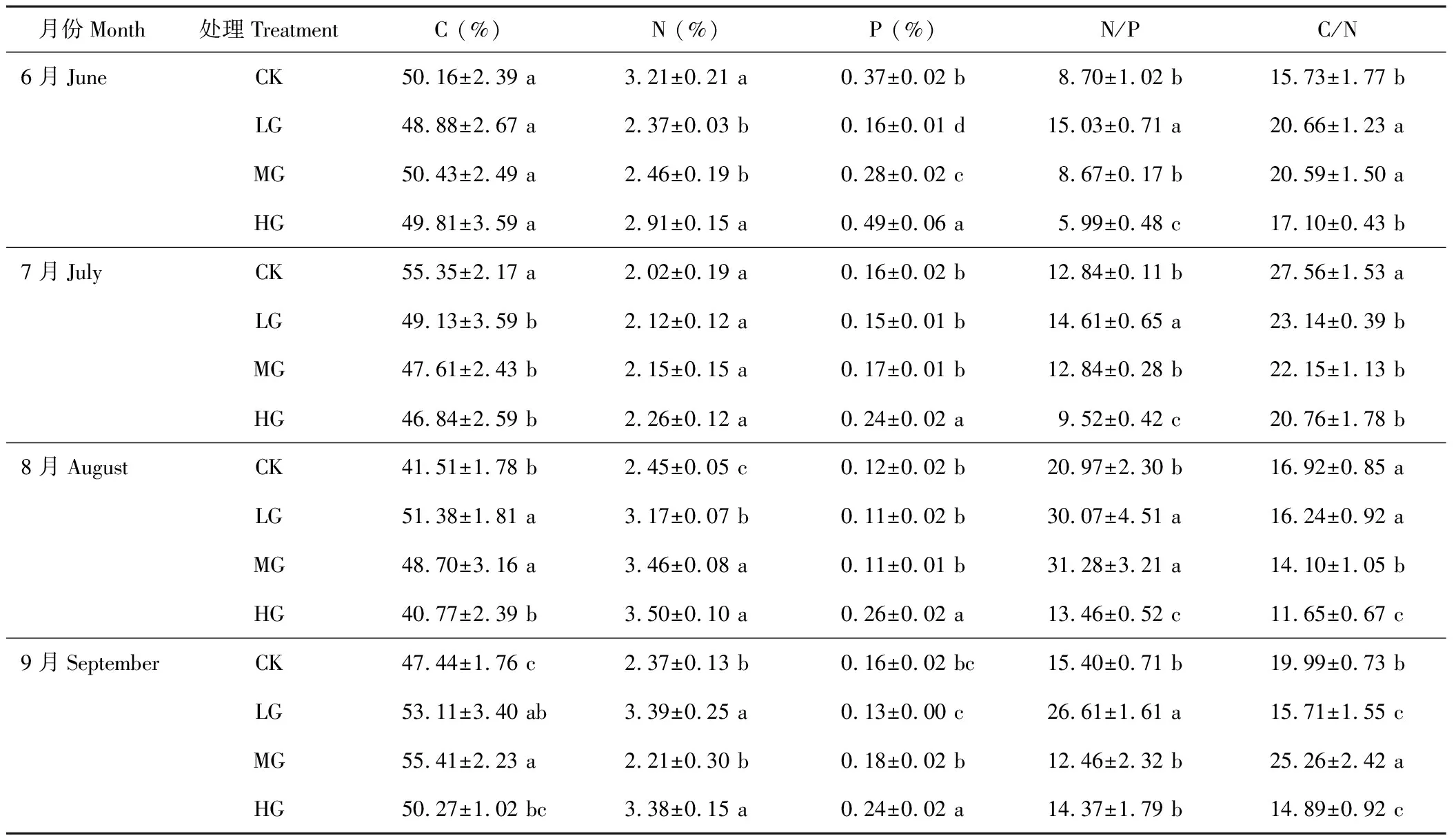
表4 小叶章叶的C、N、P化学计量比的变化Table 4 Change of ecological stoichiometry of D. angustifolia leaves
叶的C/N值季节变化规律不明显,均在8-9月份下降。在放牧初期的6月份,随着放牧强度的增加C/N呈先升高后降低的趋势,7-9月份(除中牧区)随着放牧强度的增加叶的C/N值逐渐降低。叶的N/P值范围为8.70~30.07之间,高于全株与茎的N/P值,其变化规律与全株的相同。
2.4 放牧对小叶章立枯物的养分和化学计量比的影响
由表5可以看出,在整个放牧季,轻牧区和中牧区立枯物的全碳含量均在8月份达到最大值,重牧区和对照区在6月份为最大值。受放牧的影响,随着放牧强度的增加,7-9月份立枯物的全碳含量呈先升高后降低的趋势,且均在轻牧区为最大,6月份各牧区的全碳含量大小依次为:CK>HG>MG>LG。立枯物的全氮含量:受放牧强度的影响,随着放牧强度的增加,6,7,9月份,立枯物的全氮含量均呈上升的趋势,且在9月份各放牧小区之间差异显著(P<0.05);8月份呈先升高后降低的趋势,且对照区立枯物的全氮含量显著低于放牧区(P<0.05)。立枯物全磷含量:受放牧影响,8-9月份,随着放牧强度的增加,立枯物的全磷含量呈上升趋势;6和7月份全磷的变化规律不明显,大小依次为HG>LG>MG>CK。
由于立枯物直接来源于地上植物活体,其C/N变化规律与植物类似。 N/P值,6-9月份,随着放牧强度的增加,呈先升高后降低的趋势,其值在3.53~9.43之间。
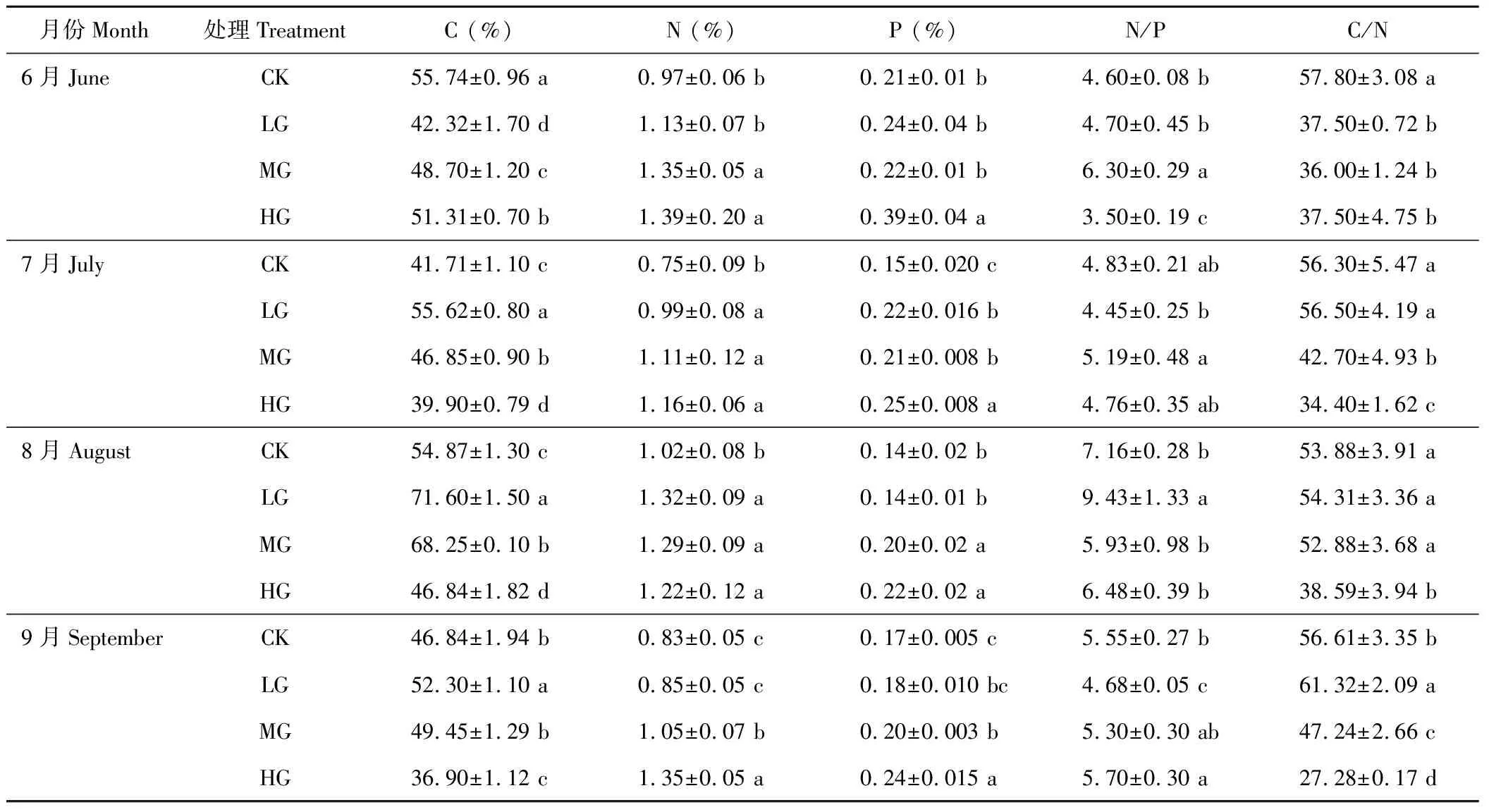
表5 小叶章立枯物C、N、P化学计量比的变化Table 5 Change of ecological stoichiometry of D. angustifolia standing litter
2.5 放牧对地下根系化学计量比的影响
由表6可以看出,0~10 cm土层根系,轻牧、中牧区随着季节的推移C/N值减小,对照区和重牧区均在7月份达到最大值。受放牧的影响,6-8月份C/N值随着放牧强度的增加呈升-降-升的趋势;9月份则为先升高后降低,且在中牧区达到最大值。6-8月份的N/P值均以中牧区为最大值,说明中度放牧使氮、磷元素更多的向根系转移。
10~20 cm土层的根系,C/N值没有明显的变化规律,除8月份外,重牧区碳氮比显著大于轻牧、中牧区(P<0.05);N/P值6月份先降低后升高,9月份逐渐升高,其他月份变化规律不明显。
2.6 放牧对土壤化学计量比的影响
由表7可以看出,0~10 cm土层,7和9月份,土壤的C/N值随着放牧强度的增加而降低;6和8月份各个放牧区C/N值差异不显著,分别在轻牧区和重牧区为最小值;N/P值的变化在各牧区差异不显著。10~20 cm土层的C/N值比0~10 cm的相对要低,但变化规律不明显。N/P值均以对照区为最高(9月份除外),且各牧区之间的差异性不显著。
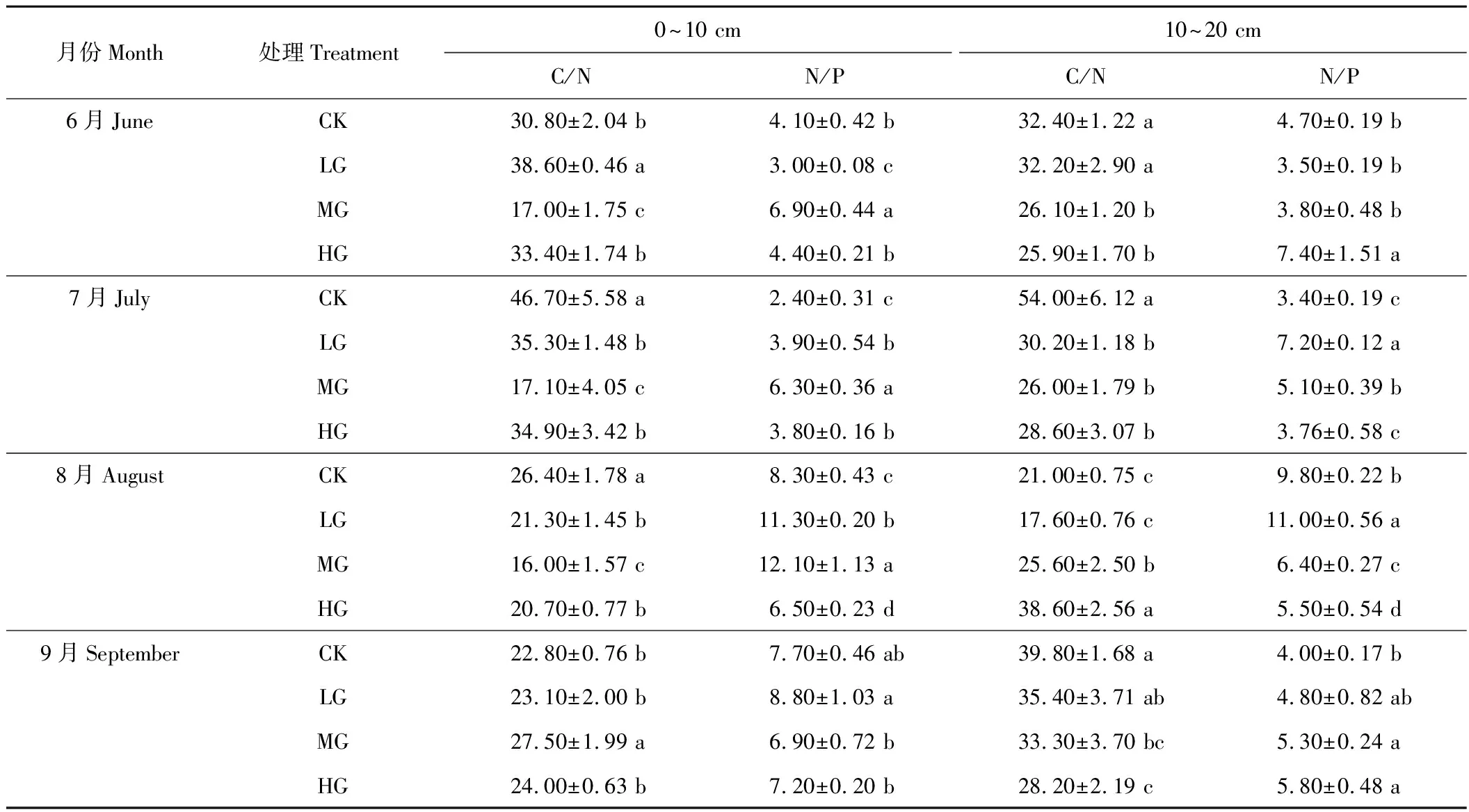
表6 地下根系C、N、P化学计量比的变化Table 6 Change of ecological stoichiometry of roots underground
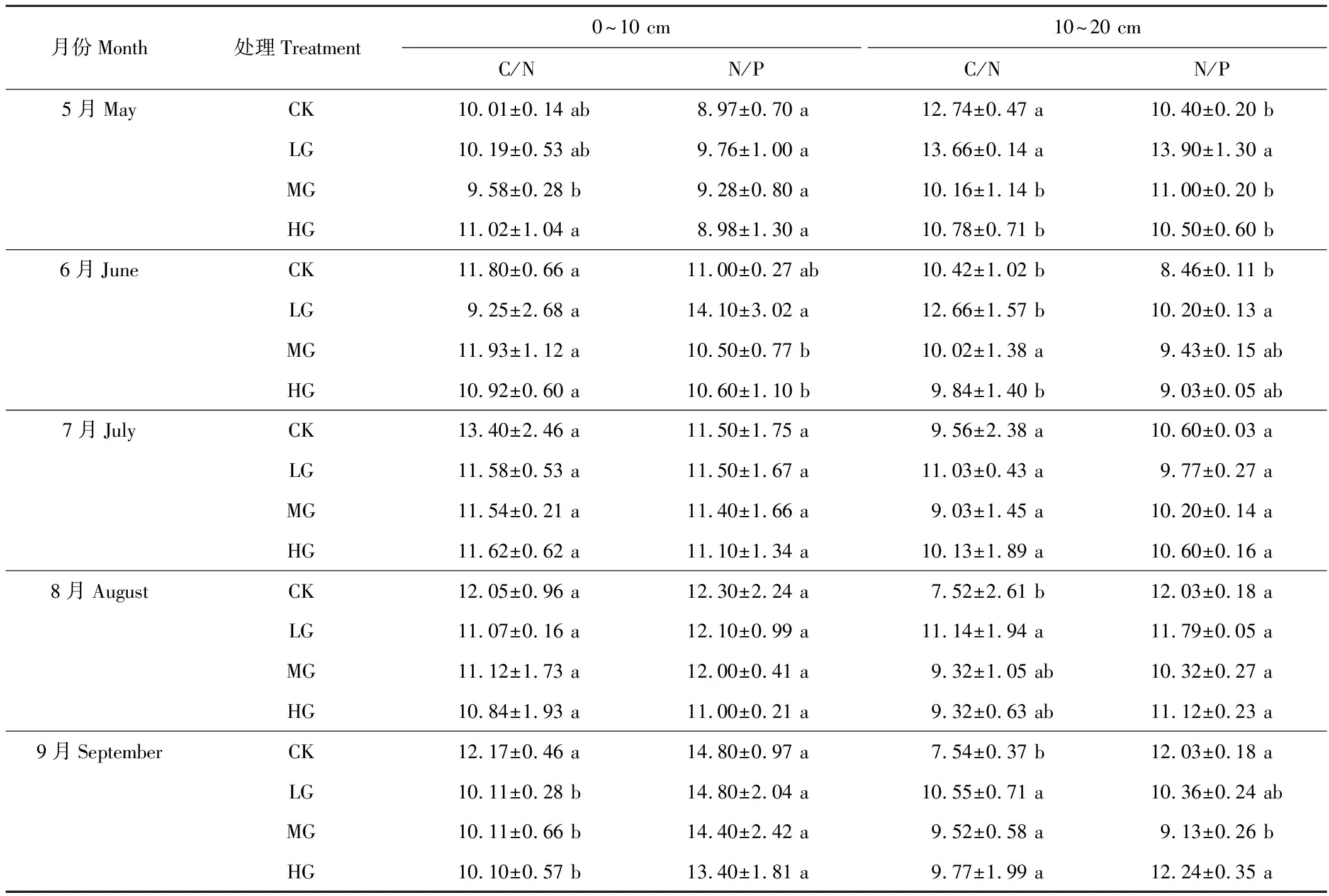
表7 土壤C、N、P化学计量比的变化Table 7 Change of ecological stoichiometry of soil
3 讨论
3.1 放牧对小叶章化学计量特征的影响
植物体内C、N代谢是植物正常生长发育和产量形成的物质基础,植物C/N代表其吸收N时同化C的能力,反映了其N利用效率和固C效率的高低[9]。因此,组织C、N含量及其生态化学计量比(C/N)是反映植株体内生理代谢状态的一项重要指标。氮和磷是陆地生态系统植物生长的主要限制性元素[10],它们在植物的各种生理代谢中彼此独立而又相互影响,共同发挥着重要的作用[11]。C是构成植物体内干物质的最主要元素,N、P分别与植物的光合作用和细胞生长分裂等重要生理活动有关。C/N和C/P值反映植物的生长速度[2],N/P反映植物生长受N或P的限制情况[12-13]。
植物体的C/N值是植物化学组成中的重要参数。李香真和陈佐忠[5]1998年研究表明,在一定的放牧强度之内,随着放牧强度的增加,地上植物体中C/N值变小,放牧率再增强,则使植物地上部C/N变大。在2001年又研究了放牧对旱黄梅衣(Xanthoparmeliacamtschadalis)C/N值的变化,结果表明随着放牧率的增加,旱黄梅衣体内的C/N降低,而在极重牧处理下又有所增加[14]。植物体C/N与养分循环速率之间密切相关,比值小时,植物残体分解速率快,C、N、P循环速率也快[15]。丁小慧等[16]研究了放牧对呼伦贝尔草地植物化学计量特征的影响,发现放牧草地的植物和土壤均具有较低的C/N,其中植物C/N显著低于围封草地。
本试验结果显示,在6-8月份小叶章全株的C/N,随着放牧强度的增加先降低后升高,且在中牧区为最小值,说明适度的放牧会使植物的C/N减小,这与李香真等[14]在2001年的研究结果相同。茎的C/N受放牧的影响变化规律不明显,只有在8月份中牧区的C/N最低。叶的C/N的变化趋势与全株的相类似。丁小慧等[16]指出植物C/N越小,残体分解速率越快,生态系统养分循环速率有所提高。分析原因可能是由于植物体的C/N较低时,植物生长速率较快,需要吸收大量的氮素和磷素,家畜的排泄物提供的速效的氮素,在一定程度上满足了植物对氮素的需要,使得养分周转的周期变短,最终导致放牧草地养分循环速率高于围封草地。因为枯落物来自活体植物,所以枯落物的C/N直接受控于活体植物的碳氮含量,所以枯落物的C/N的变化特点和活体植物类似。本试验中叶的N/P值高于全株和茎N/P值,且随着放牧强度的增加有先升高后降低的趋势。N/P值的变化主要受环境营养元素的影响,放牧对N/P值的影响主要是通过改变土壤的性状来实现的。He等[17]2008年研究指出叶片作为植物的主要光合器官,其N/P的大小经常被用来表明生态系统生产力受到哪种元素的限制作用,但是这种限制关系会随着外界环境的改变而改变。
3.2 放牧对小叶章地下部分化学计量特征的影响
草地地下部分的化学计量特征的变化与地上部分不同,土壤C/N、C/P通常反映有机质的分解与积累,N、P的富瘠和有效程度,一定范围内作为土壤肥力的指标,并影响植物体的养分积累与分配[18]。肖红艳等[19]在2012年研究发现,放牧干扰能够改变土壤动物的群落组成,且在不同放牧强度下干扰程度差异较大,而土壤中微生物的分解状况是影响土壤含量变化的重要原因。本试验表明,7-9月份放牧强度的增加会造成0~10 cm土壤C/N值降低,但趋势不明显。这与魏巍等[20]2012年的研究结果相似。分析原因一方面由于放牧改变了土壤呼吸的微环境[21],使土壤的物理性质及微生物的生存环境都有所改变,增加了N、P的矿化。另一方面,家畜排泄物含有大量N、P元素,使微生物的活性增强,N、P矿化作用加强。10~20 cm土层的变化规律性不明显,说明放牧主要引起土壤表层理化性质的变化。高永恒[22]2007年研究指出,放牧降低了地上部分的C/N,却增加了根系的C/N,魏巍等[20]研究了放牧对高寒杜鹃(Rhododendron)灌丛草地的根系化学计量特征的影响,结果表明0~10 cm土层随放牧强度的增加根系的C/N值降低,N/P值升高。本试验结果显示,0~10 cm土层的根系,在7、8月份其C/N值随着放牧强度的增加先降低后升高,均在中度放牧下为最低,原因可能是在适度放牧的刺激下,植物将更多的营养元素分配到了其地下部分,可能是植物对放牧干扰的一种补偿性反应[23]。6-9月份的N/P值随着放牧强度的增加先降低后升高,且在中牧区达到最大值,在重度放牧下又有所降低。
4 结论
本研究基于小叶章草甸开发利用的现状,以优势植物小叶章为研究对象,将其分为地上活体、立枯物、地下根系3个部分,在不同放牧强度下对这三个部分生物量及碳、氮、磷元素化学计量特征及动态变化进行了分析,为草地管理和元素循环中数学模型的建立提供理论及实践依据。
1)随着放牧强度的增加,小叶章活体与立枯物的全氮(TN)、全磷(TP)含量也随之增加,放牧使小叶章叶的全氮含量明显增加,而茎的变化不显著;0~10 cm土层根系的全碳、全氮、全磷变化比10~20 cm的更敏感;放牧降低了土壤全碳(TC)含量,从单月份来看,土壤表层的全氮含量随放牧强度的增加呈先增加后降低的趋势,且以重牧区为最小,从季节来看,各小区在9月份时的全氮含量均高于6月份。
2)在6-8月份小叶章全株及叶的C/N值随着放牧强度的增加先降低后升高,且在中牧区降为最小值,茎的C/N值受放牧的影响变化规律不明显,只有在8月份中牧区表现为最小值;N/P值的变化主要受外界环境营养元素的影响,放牧对N/P值的影响主要是通过改变土壤的性状来实现的。
3)放牧主要影响土壤表层的理化性状以及所含根系的元素含量,7-9月份随着放牧强度的增加,0~10 cm土壤的C/N值降低;7、8月份0~10 cm土层根系的的C/N值随着放牧强度的增加先降低后升高,且在中牧区为最小值,而N/P值却在中牧区为最大值。
猜你喜欢
矿山安全信息(2022年22期)2022-11-24
High Technology Letters(2021年4期)2022-01-09
西藏艺术研究(2021年3期)2021-06-02
河北果树(2020年4期)2020-11-26
中国新闻周刊(2019年46期)2019-12-20
西藏农业科技(2019年1期)2019-07-25
海峡姐妹(2019年1期)2019-03-23
中成药(2018年9期)2018-10-09
铜仁学院学报(2018年5期)2018-06-11
中国水利(2015年4期)2015-02-28
