
不同地理种群紫茉莉茎秆扦插繁殖及种子萌发特性研究
2014-10-29 11:34蒋拥军闫小红刘文阳
井冈山大学学报(自然科学版) 2014年2期
周 兵,蒋拥军,闫小红,王 宁,刘文阳
不同地理种群紫茉莉茎秆扦插繁殖及种子萌发特性研究
*周 兵1,2,蒋拥军3,闫小红1,2,王 宁1,2,刘文阳1
(1. 井冈山大学生命科学学院,江西,吉安 343009;2. 江西省生物多样性与生态工程重点实验室,江西,吉安 343009;3.遂川县林业局,江西,遂川 343900)
为探索紫茉莉不同地理种群间繁殖特性的差异,本研究以11个不同地理总群的紫茉莉为研究对象,通过室内生物测定的方法对不同地理种群紫茉莉种子萌发及茎秆扦插繁殖特性进行了比较研究,并分析了两者之间的关系。结果表明,不同地理种群紫茉莉种子萌发特性存在差异,江苏(JS)地理种群种子萌发率和发芽势最高,分别为77.50%和72.00 %,福建(FJ)地理种群种子的萌发率和广西(GX)地理种群种子的发芽势最低,分别为22.50%和3.50 %。不同地理种群紫茉莉茎秆扦插繁殖特性也存在差异,北京(BJ)等8个地理种群的扦插成活率相对较高,均高于90 %,而JS种群的最低,为46.67 %;GX种群插穗平均生根数最大(69.83根),显著高于其它地理种群(<0.05),重庆(CQ)种群的最低(6.57根);GX种群插穗平均根长最大,为2.74 cm,上海(SH)的最小,为1.04 cm。种子萌发率与茎秆扦插成活率间存在负相关,但没有显著性,表明紫茉莉的无性繁殖与有性繁殖间可能存在一定的权衡。本研究不仅揭示了紫茉莉的繁殖表型可塑性,也为制定更具针对性的防控措施提供了指导作用。
地理种群;紫茉莉;种子萌发;扦插
繁殖是入侵种建群时面临的根本挑战,因而,与繁殖有关的特性直接影响到入侵植物的归化和入侵的成功[1]。植物的繁殖方式包括无性繁殖和有性繁殖两种,无性繁殖有利于维持居群的大小,有性繁殖有利于维持居群的进化潜力和对变化环境的适应能力[2]。一些入侵植物兼有两种繁殖方式,且两种方式产生后代个体的能力均很惊人,如大叶落地生根()[3]、剑叶金鸡菊()[4]等,而一些入侵植物虽然也兼有两种繁殖方式,但两种方式产生后代个体的能力存在一定差异,如空心莲子草()[5]、美洲商陆()[6]等。同时,入侵植物宽广的分布范围和快速扩散能力使其在入侵过程中不断面对和适应新的环境[7-8],形成不同的地理种群。目前的研究更多的采用分子生物学手段研究不同地理种群间的遗传差异,分析之间的亲缘关系,从而揭示其入侵历史[9-10],而从开花物候、繁殖分配、繁育系统等繁殖特性方面研究不同地理种群间的形态分化和对环境的适应性相对较少[11-12]。
紫茉莉()为紫茉莉科(Nyctaginaceae)紫茉莉属(L.)多年生草本植物,原产南美热带地区,最初作为药用和观赏类植物人工有意引入,各地常逸为野生,现广泛分布于全国各地,对当地的生物多样性造成潜在伤害[13]。紫茉莉具有灵活的交配系统[14],种子繁殖能力强,根和茎也具有营养繁殖能力[15],对其它植物具有较强的化感抑制作用[15],对重金属有较强的耐受性[16-17]等特性增强了其入侵性,使其成功入侵不同生境成为可能。然而,作为广泛入侵种,紫茉莉不同地理种群间的生物学特性,尤其是繁殖特性间是否存在差异,目前还未见报道。为此,本研究从不同地理种群紫茉莉茎秆扦插繁殖能力和种子萌发能力开展比较研究,以期探明:(1)紫茉莉不同地理种群间的扦插繁殖能力是否存在差异?(2)紫茉莉不同地理种群间的有性繁殖能力是否存在差异?(3)紫茉莉的无性繁殖和有性繁殖间是否存在权衡?本研究将不仅为从繁殖可塑性的角度揭示该入侵植物的入侵机制提供了理论基础,也为制定针对性的防治策略提供了理论指导。
1 材料与方法
1.1 植物材料
紫茉莉不同地理种群的种子于2011年收集于各省市,收集情况如下:BJ:北京中国农业大学校园;SC:四川成都四川大学校园;CQ:重庆缙云山;FJ:福建福州福建农林大学校园;AH:安徽宣城;SH:上海同济大学校园;ZJ:浙江临安;JX:江西吉安;GX:广西南宁;JS:江苏南京南京林业大学校园;GZ:贵州安顺。
1.2 种子萌发
于2012年4月选择籽粒饱满的不同地理种群种子若干,浸种1 h,取出播种于铺有7 cm厚素沙的塑料方盆(45 cm × 32 cm × 11 cm)中,并在种子上加盖1 cm厚素沙,每盆50粒,设4个重复,浇水保持沙面湿润,每天观察记录种子萌发情况,第10天统计种子萌发率,第7天统计发芽势。具体公式如下:
萌发率= 10 d发芽种子数/供试种子数 × 100%
发芽势= 7 d发芽种子数/供试种子数 × 100%
1.3 茎秆扦插测试
1.3.1 扦插处理
于2012年9月取母枝上当年生、无病虫害、健壮、芽饱满的绿枝为插穗,长度5~10 cm,去除叶片,上端剪成平切口,切口距芽l~2 cm,下端成45度斜切口。插前插穗基部速蘸浓度为1000 μg/mL 的IBA溶液10 s,扦插于铺有7 cm厚素沙的塑料方盆(45 cm × 32 cm × 11 cm)中,扦插的深度为2~3 cm,株行距为4 cm ×4 cm。试验采取完全随机试验设计,每种群为一个处理,3次重复,每重复30~40个插穗。
1.3.2 扦插后管理
扦插后浇透水,前7 d,每天喷水6次,从上午8:00~18:00,每2 h喷1次水,相对湿度保持在60 %~90 %,之后适当减少喷水次数。扦插后40 d,统计插穗的生根率、并随机选取10株扦插成活的插穗统计生根数、测量根长等。
1.4 数据统计分析
使用Excel软件进行数据处理,并运用SPSS19.0统计分析软件进行差异显著性分析及pearson相关性分析。
2 结果与分析
2.1 不同地理种群的种子萌发特性
2.1.1 不同地理种群种子的基本特征
表1显示了不同地理种群紫茉莉种子的基本特征,可见不同种群间存在差异。11个地理种群中,其种子的竖径、横径及百粒重分别为4.82~5.61 mm、3.74~5.00 mm、5.3343~7.3870 g。BJ、AH、JX、GX和GZ地理种群种子的竖径较大,分别为5.38、5.61、5.18、5.49、5.48 mm,且显著大于其它地理种群(<0.05);AH地理种群种子横径最大(5.00 mm),显著大于除SQ和SH地理种群之外的其它地理种群(<0.05);BJ地理种群种子百粒重最大(7.3870 g),JS的最小(5.3343 g),前者是后者的1.38倍。
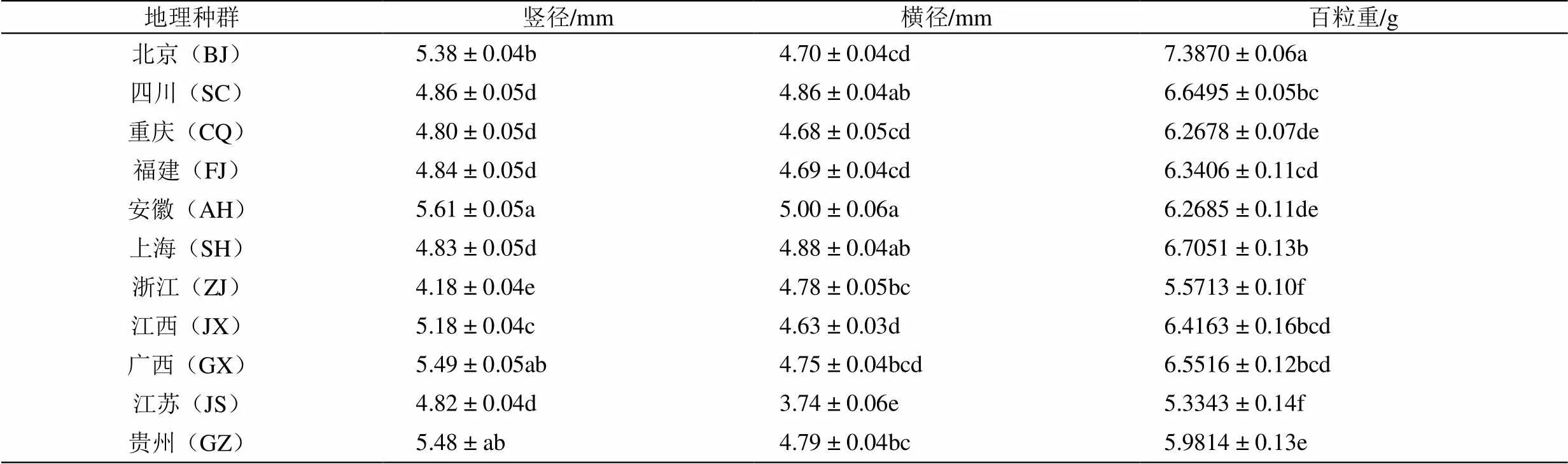
表1 不同地理种群紫茉莉种子的基本特征
2.1.2 不同地理种群种子的萌发
不同地理种群紫茉莉种子的萌发存在差异(表2)。在11个地理种群中,JS地理种群种子的萌发率最高(77.50 %),AH次之(76.50 %),二者之间没有显著差异(>0.05),但显著高于其它地理种群种子的萌发率(<0.05),FJ地理种群种子的萌发率最低,为22.50 %,不足JS地理种群的1/3。JS地理种群种子的发芽势最高,为72.00 %,显著高于其它地理种群(<0.05),而GX地理种群最低(3.50 %);种子萌发率高的发芽势并不一定高,AH地理种群种子萌发率显著高于CQ地理种群(55.50 %) (<0.05),但前者的发芽势(32.50%)显著低于后者(43.50 %) (<0.05)。
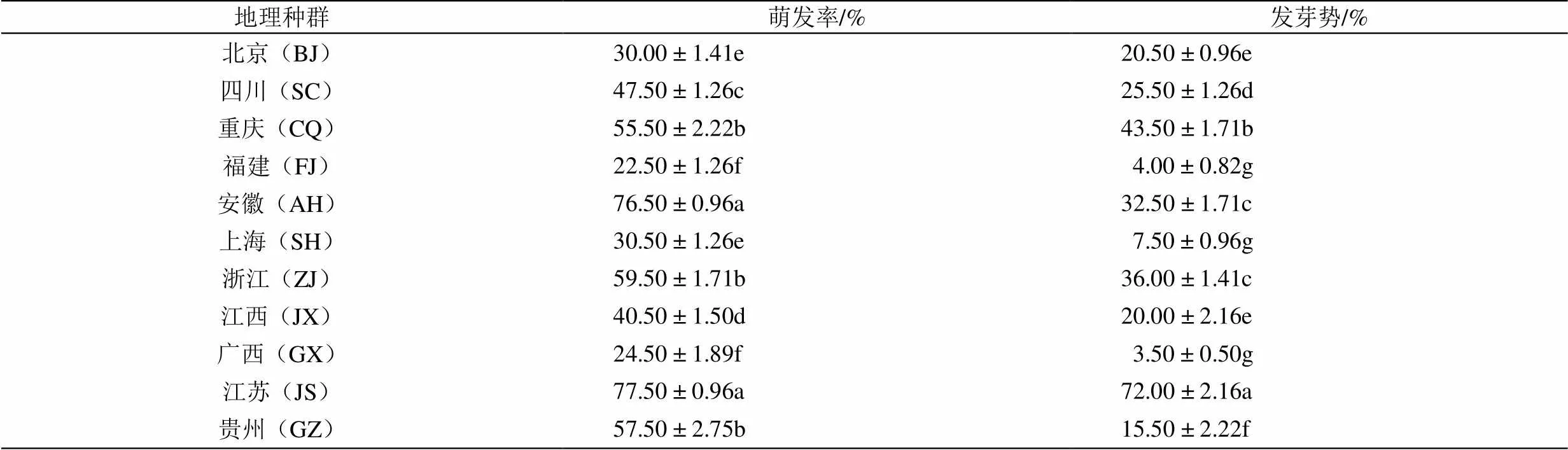
表2 不同地理种群紫茉莉种子的萌发特性
2.2 不同地理种群茎秆扦插繁殖特性
2.2.1不同地理种群茎秆扦插繁殖的成活率
紫茉莉不同地理种群茎秆扦插繁殖能力见图1。紫茉莉不同地理种群茎秆扦插成活率存在差异,在11个地理种群中,北京(BJ)、四川(SC)、重庆(CQ)、福建(FJ)、江西(JX)、广西(GX)和贵州(GZ)等8个地理种群的扦插成活率相对较高,均高于90 %,尤其是CQ种群,其成活率为100 %;而浙江(ZJ)和江苏(JS)两个地理种群茎秆扦插的成活率相对较低,分别为74.44 %和46.67 %,且与前面8个地理种群的扦插成活率间存在显著差异(<0.05)。
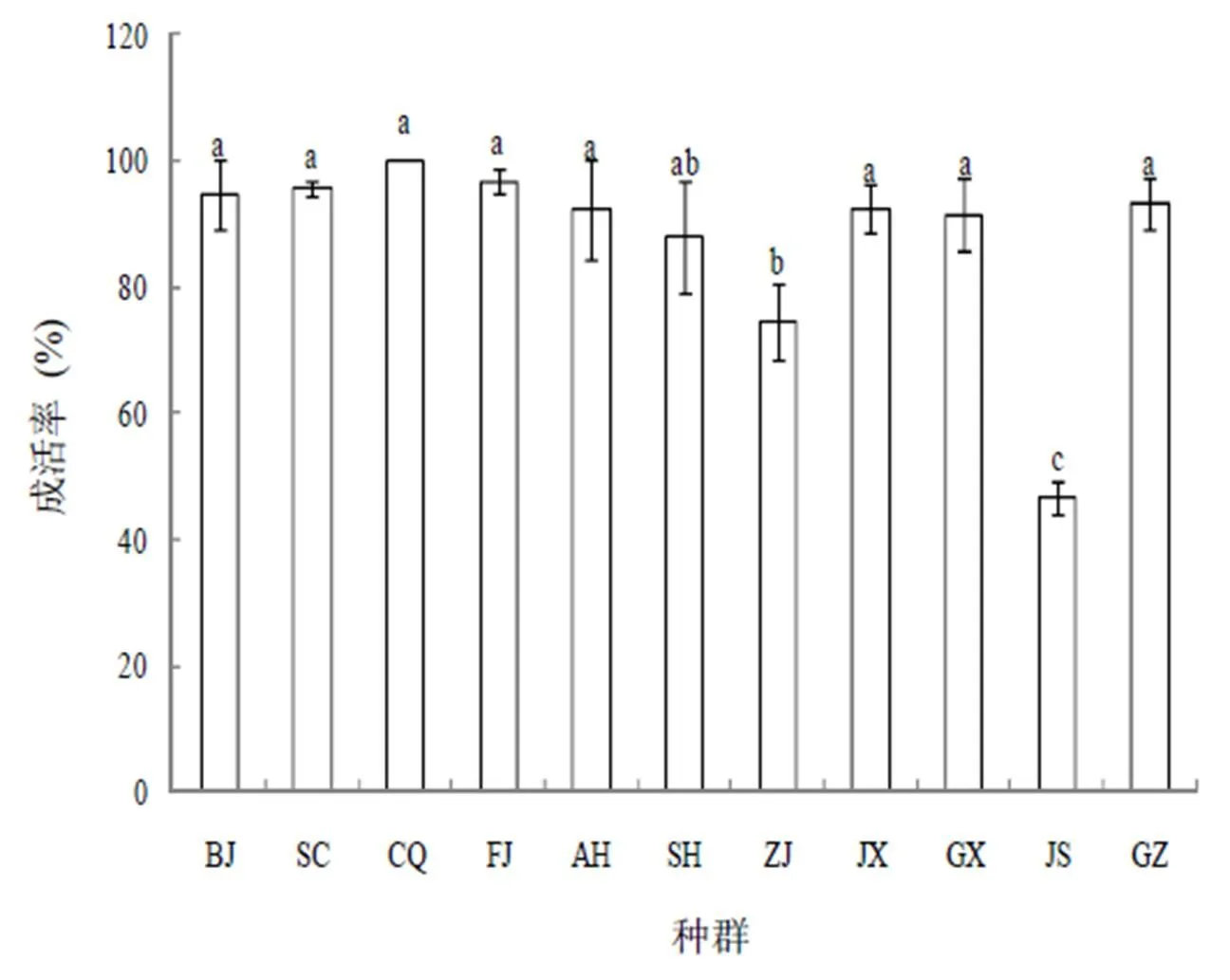
图1 紫茉莉不同地理种群茎秆扦插成活率
图中不同小写字母表示差异显著(<0.05),下同
2.2.2 不同地理种群茎秆扦插生根数及插穗根长
紫茉莉不同地理种群茎秆插穗平均生根数存在较大差异,其中JX、GX、JS和GZ等4个地理种群插穗平均生根数相对较多,均大于29.06根,而BJ等其它7个地理种群插穗平均生根数相对较少,均小于15.96根。且GX种群插穗平均生根数最大,为69.83根,显著高于其它地理种群(<0.05),CQ种群的最低,为6.57根,前者是后者的10.63倍(图2)。不同地理种群紫茉莉茎秆扦插插穗根长也存在差异。GX种群插穗平均根长最大,GZ种群次之,根长分别为2.74和2.51 cm,两种群间没有显著差异(>0.05)。GX种群和GZ种群插穗平均根长显著大于BJ等其它9个地理种群(<0.05),在9个地理种群中,SH地理种群插穗平均根长最小,为1.04 cm,除与BJ地理种群间没有显著差异外,其与SC等其它7个地理种群间均存在显著差异(<0.05)(图2)。
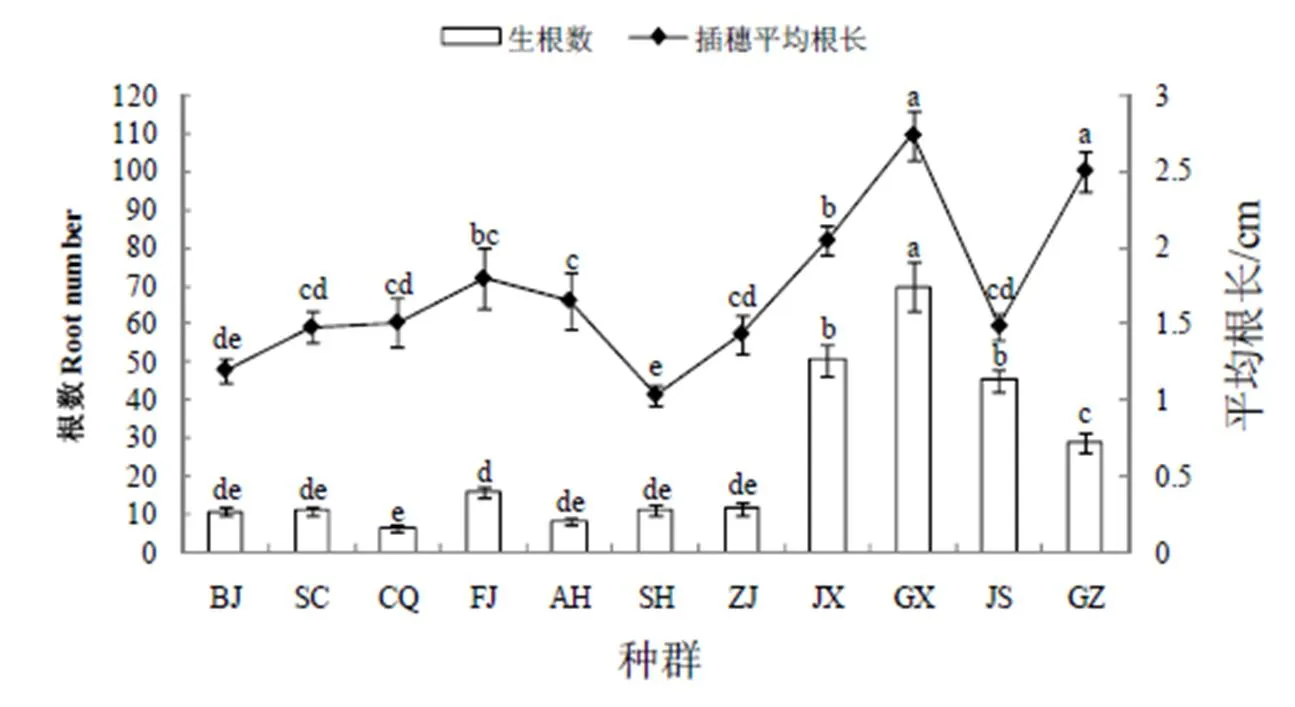
图2 紫茉莉不同地理种群茎秆扦插生根数及插穗根长
2.3 不同地理种群的种子萌发率与茎秆扦插成活率间的相关性
图3显示了紫茉莉不同地理种群种子萌发率与茎秆扦插成活率间的相关性,可见,两者之间存在负相关,且以二次方程最能拟合二者之间的关系,模拟方程为y = -0.0174x2+1.3223x+70.299,相关系数为0.6402。Pearson相关性分析表明,二者间存在负相关,但没有显著性(>0.05)。

图3 紫茉莉不同地理种群茎秆扦插成活率与种子萌发率间的关系
3 讨论
表型可塑性是个体通过改变自身形态结构和/或生理特性而对环境变化作出反应的能力[18],其可以使物种具有更宽的生态幅和更好的耐受性,占据更加广阔的地理范围和多样化的生境,成为广布种[19]。外来入侵植物常具有较强的表型可塑性,体现在植株的高度、种子的大小、各器官的大小、结构等个体的形态[20-22],生长繁殖行为[11-23]以及光适应性、水分利用效率等生理生化特性[24]等方面。紫茉莉为广布外来入侵植物之一,在全国范围内均有分布,研究表明,11个不同地理种群种子的大小、重量存在差异,安徽(AH)地理种群种子最大,北京(BJ)的百粒重最大,而不同地理种群种子的萌发率和发芽势也存在差异,江苏(JS)地理种群种子萌发率和发芽势最高,分别为77.50和72.00 %,福建(FJ)地理种群种子的萌发率和广西(GX)地理种群种子的发芽势最低,分别为22.50和3.50 %,且萌发率高并不表明其发芽势也高,AH地理种群种子萌发率显著高于CQ地理种群(55.50 %) (<0.05),但前者的发芽势(32.50 %)显著低于后者(43.50 %)(<0.05)。一般认为,种子的重量与种子的传播距离、种子库寿命、定居成功率相关,种子的形状与种子库寿命相关,种子表现出的高度多样性,使其适应于强烈人为干扰的环境[25]。小粒种子植物比大粒种子植物具有更大的多度空间,占据更广阔的空间[26],但小粒种子所含的营养物质有限,从而对种子的繁殖力产生负面效应[27]。入侵植物不同地理种群种子特性方面的差异,在紫茎泽兰()[11]、贯叶连翘()[28]等其它入侵植物中也得到了验证。
在入侵过程中,因受到不同环境条件的影响,入侵植物在不同种群间的繁殖方式、繁殖能力方面存在差异[29]。研究表明,紫茉莉不同地理种群间,不仅种子的萌发特性存在差异,且其茎秆扦插繁殖特性也存在差异,在11个地理种群中, BJ、CQ、FJ、GX、四川(SC)、江西(JX)和贵州(GZ)等8个地理种群的扦插成活率相对较高,均高于90%,而JS地理种群茎秆扦插的成活率相对较低为46.67%(<0.05)。这种不同地理种群间繁殖特性的差异在其它入侵植物中也被证实了。紫茎泽兰的云南不同地理种群间在分蘖数、开花数、种子产量、种子萌发率等繁殖特征方面存在显著差异[11]。入侵植物酢浆草()在由南美向北美扩散的过程中,其交配系统发生多次从三型花柱到二型花柱的转变[12]。入侵植物和入侵地和原产地种群交配系统的自交亲和力存在显著差异[30]。这些繁殖特征的差异体现了不同种群间的不同繁殖能力和繁殖方式,表明紫茉莉具有较强的适应能力和繁殖表型可塑性,从一定程度上增强了其入侵性。
面临环境变异,植物可能演化出最佳的繁殖对策,以达到对局部环境条件的最佳适应,这种适应常表现在生活史特征或生殖方式上的变异上[31]。在生殖方式方面,当植物同时存在无性繁殖和有性繁殖式,而个体内可用资源有限,那么投资到两种繁殖方式的资源通常呈现一种权衡关系[32-33]。紫茉莉不同地理种群茎秆扦插成活率和种子萌发率的相关性分析表明两者之间存在负相关,虽然这种相关性没有达到显著水平,但在一定程度上从繁殖力的角度证明了两种繁殖方式共存时之间可能存在的权衡关系,同时,紫茉莉的无性繁殖还存在根的萌蘖,综合分析两种无性繁殖方式的繁殖能力与其种子萌发的能力间的相关性,将能更全面的反映两种繁殖方式之间的关系。作为入侵植物,这种权衡关系的存在将增强了其对不同生境的适应能力,有利于其入侵和扩散。
可见,紫茉莉不同地理种群间的种子萌发及茎秆扦插繁殖特征存在差异,体现了该入侵植物较强的繁殖表型可塑性,且两种繁殖方式间存在一定的权衡关系。研究结果从繁殖适应性的角度初步揭示了该入侵植物的入侵机制,繁殖特征的差异表明抑制其繁殖将具有更好的防控效果,比如在种子萌发期及幼叶生长期进行施药处理等,从而为制定更具针对性的防控措施提供了理论指导作用。
[1] Pysek P, Richardson D M. Traits associated with invasiveness in alien plants: Where do we stand[A]? In: Nentwig W, et al. Biological Invasions, Section Ⅱ[C]. Berlin: Verlag-Springer, 2007.
[2] 张玉芬, 张大勇. 克隆植物的无性与有性繁殖对策[J]. 植物生态学报, 2006, 30(1): 174-183.
[3] Herrera I, Nassar J M. Reproductive and recruitment traits as indicators of the invasive potential of(Crassulaceae) and(Apocynaceae) in a Neotropical arid zone[J]. Journal of Arid Environments, 2009, 73(11): 978-986.
[4] 曾建军, 肖宜安, 孙敏. 入侵植物剑叶金鸡菊的繁殖特征及其与入侵性之间的关系[J]. 植物生态学报, 2010, 34(8): 966-972.
[5] Sainty G, McCorkelle G and Julien M. Control and spread of alligator weed(Mart.) Griseb., in Australia: Lessons for other regions[J]. Wetlands Ecology and Management, 1998, 5(3): 195-201.
[6] 周兵, 闫小红, 肖宜安, 等. 外来入侵植物美洲商陆的繁殖生物学特性及其与入侵性的关系[J]. 生态环境学报, 2013, 22(4): 567-574.
[7] Goodwin B J, Mcallister A J, Fahrig L. Predicting invasiveness of plant species based on biological information[J]. Conservation Biology, 1999, 13(2): 422-426.
[8] Alpert P, Bone E, Holzaprel C. Invasiveness, invisibility and the role of environmental stress in the spread of non-native plants[J]. Perspectives in Plant Ecology, Evolution and Systematics, 2000, 3(1): 52-66.
[9] Zong X X, Guan J P, Wang S M, et al. Genetic diversity and core collection of alienL. germplasm[J]. Acta Agronomica Sinica, 2008, 34(9): 1518-1528.
[10] Ren M X, Li X Q, Ding J Q. Genetic variation and spread pattern of invasivearound China’s Three Gorges Dam[J]. Acta Oecologica, 2010, 36(6): 599-603.
[11] 赵相健, 刘文耀, 周蒙, 等. 不同地理种群紫茎泽兰生长繁殖特征的比较研究[J]. 西北植物学报, 2009, 29(6): 1252-1258.
[12] Gardner A G, Vaio M, Guerra M, et al. Diversification of the American bulb-bearing(Oxalidaceae): Dispersal to North America and modification of the tristylous breeding system[J]. American Journal of Botany, 2012, 99(1): 152-164.
[13] 徐海根, 强胜. 中国外来入侵物种编目[M]. 北京: 科学出版社, 2004.
[14] 陈香, 胡雪华, 肖宜安, 等. 紫茉莉的花部综合特征与繁育系统[J]. 生态学杂志, 2008, 27(10): 1653-1658.
[15] 许桂芳, 刘明久, 李雨雷. 紫茉莉入侵特性及其入侵风险评估[J]. 西北植物学报, 2008, 28(4): 765-770.
[16] Yu Z, Zhou Q. Growth responses and cadmium accumulation ofL. under interaction between cadmium and phosphorus[J]. Journal of Hazardous Materials, 2009, 167(1-3): 38-43.
[17] 薛生国, 朱锋, 叶晟, 等. 紫茉莉对铅胁迫生理响应的FTIR研究[J]. 生态学报, 2011, 31(20): 6143-6148.
[18] Macdonald S E, Chinnappa C C. Population differentiation for phenotypic plasticity in thecomplex[J]. American Journal of Botany, 1989, 76(11): 1627-1637.
[19] Sultan S E. Phenotypic plasticity and plant adaptation[J]. Acta Botanica Neerlandica, 1995, 44(4): 363-383.
[20] Droste T, Flory S L, Clay K. Variation for phenotypic plasticity among populations of an invasive exotic grass[J]. Plant Ecology, 2010, 207(2): 297-306.
[21] Riis T, Lambertini C L, Olesen B, et al. Invasion strategies in clonal aquatic plants: are phenotypic differences caused by phenotypic plasticity or local adaptation[J]? Annals of Botany, 2010, 106(5): 813-822.
[22] 周兵, 闫小红, 肖宜安, 等. 外来入侵植物大狼把草种群构件生物量结构研究[J]. 广西植物, 2012, 32(5): 650-655.
[23] Monty A, Bizoux J P, Escarré J, et al. Rapid plant invasion in distinct climates involves different sources of phenotypic variation[J]. PLoS ONE, 2013, 8(1): 1-10.
[24] Matesanz S, Horgan-Kobelski T, Sultan S E. Phenotypic plasticity and population differentiation in an ongoing species invasion[J]. PLoS ONE, 2012, 7(9): 1-13.
[25] Weiher E, van der Werf A, Thompson K, et al. Challenging theophrastus: A common core list of plant traits for functional ecology[J]. Journal of Vegetation Science, 1999, 10(5): 609-620.
[26] 刘志民, 蒋德明, 高红瑛, 等.植物生活史繁殖对策与干扰关系的研究[J]. 应用生态学报, 2003, 14(3): 418-422.
[27] 郭水良, 王勇, 曹同. 杂草繁殖方式的多样性及其对环境的适应[J]. 上海师范大学学报:自然科学版, 2006, 35(3): 103-110.
[28] Maron J L, Vilue M, Bommarco R, et al. Rapid evolution of an invasive plant[J]. Ecological Monographs, 2004, 74(2): 261-280.
[29] 史刚荣, 马成仓. 外来植物成功入侵的生物学特征[J]. 应用生态学报, 2006, 17(4): 727-732.
[30] Petanidou T, Godfree R C, Song D S, et al. Self-compatibility and plant invasiveness: Comparing species in native and invasive ranges[J]. Perspectives in Plant Ecology, Evolution and Systematics, 2012, 14(1): 3-12.
[31] 袁龙义, 李守淳, 李伟, 等. 水深对刺苦草生长和繁殖策略的影响研究[J]. 江西师范大学学报:自然科学版, 2007, 31(2): 156-160.
[32] 范邓妹, 杨永平. 不同生境下珠芽蓼(蓼科)的繁殖策略比较[J]. 云南植物研究, 2009, 31(2): 153-157.
[33] 彭德力, 张志强, 牛洋, 等. 高山植物繁殖策略的研究进展[J]. 生物多样性, 2012, 20(3): 286-299.
Stem cutting propagation and seed germination of Mirabilis jalapa with different geographical populations
*ZHOU Bing1,2, JIANG Yong-jun3, YAN Xiao-hong1,2, WANG Ning1,2, LIU Wen-yang1
(1. School of Life Sciences, Jinggangshan University, Ji’an, Jiangxi 343009, China; 2. Jiangxi Province Key Laboratory for Biodiversity Science and Ecological Engineering, Ji’an, Jiangxi 343009, China; 3. Forestry Bureau of Suichuan County, Suichuan, Jiangxi 343900, China)
To investigate the difference of the reproductive characteristics among 11 different geographical populations of, characteristics of seed germination and stem cutting of different populations were comparatively studied, and the relationship between them was analyzed. The results showed that, there were differences in the characteristics of the seed germination among different populations, the seed germination rate and germinative force of Jiangsu (JS) population were the highest, with the values of 77.50 and 72.00% respectively, but lowest seed germination rate (22.50%) for Fujiang (FJ) population and lowest gerninative force (3.50%) for Guangxi (GX) population. There were differences in the characteristics of stem cutting reproduction among different populations also, cutting survival rates of eight populations as Beijing (BJ) etc. were relatively high, with the values over 90%, but lowest for JS population (46.67%); the rooting number of the cutting slips for GX population was the highest (69.83 pieces), significantly higher than the others (<0.05), the lowest for Chongqing (CQ) population (6.57 pieces); the root length of the cutting slips for GX population was the highest (2.74 cm), and the lowest for Shanghai (SH) population (1.04 cm). There was negative correlation between seed germination rates and stem cutting survival rates, without significance, which showed there perhaps was some trade-off between asexual reproduction and sexual reproduction of. The study not only reveals the reproductive phenotypic plasticity of, but also provides guiding function for specific strategies in its control.
geographical population;; seed germination; cutting
Q944
A
10.3969/j.issn.1674-8085.2014.02.023
1674-8085(2014)02-0101-06
2013-12-12;
2014-01-26
国家自然科学基金项目(31360090);江西省自然科学基金项目(20114BAB214011);江西省高等学校科技落地计划项目(KJLD12079)
*周 兵(1977-),男,湖北黄梅人,副教授,博士,主要从事植物生态学方面的教学与研究(E-mail: zhoubing113@126.com);
蒋拥军(1971-),男,江西遂川人,主要从事森林生态学方面的研究(E-mail: fengguanping@126.com);
闫小红(1977-),女,内蒙古赤峰人,实验师,硕士,主要从事植物生理学方面的教学与研究(E-mail: yanxiaohong325@126.com);
王 宁(1979-),男,山东淄博人,副教授,博士,主要从事生态学方面的教学与研究(E-mail: wangning13@126.com);
刘文阳(1990-),男,安徽阜阳人,井冈山大学生命科学学院园林专业2010级本科生(E-mail: 1479322382@qq.com).
猜你喜欢
江西农业大学学报(2022年3期)2022-07-07
课外生活(小学1-3年级)(2021年8期)2021-08-28
基层中医药(2021年1期)2021-07-22
现代园艺(2020年19期)2020-10-02
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
农业机械学报(2019年4期)2019-04-29
现代农村科技(2019年4期)2019-01-06
畜牧与饲料科学(2018年5期)2018-06-13
金色少年(奇趣科普)(2017年6期)2017-07-25
