
两种麝鼩的核型和G-带研究
2014-10-25 06:38:56蔡灵超叶建平陈萍萍
绍兴文理学院学报(自然科学版) 2014年2期
蔡灵超 叶建平 陈 冲 陈萍萍
(绍兴文理学院 生命科学学院,浙江 绍兴312000 )
两种麝鼩的核型和G-带研究
蔡灵超 叶建平 陈 冲 陈萍萍
(绍兴文理学院 生命科学学院,浙江 绍兴312000 )
采用空气干燥法制作染色体标本,以胰酶法制作G带,报道了云南2种麝鼩的核型和G带.白尾梢麝鼩(Crocidurafuliginosa)二倍染色体数目(2n)为40,常染色体臂数(FN)为50;灰麝鼩(Crociduraattenuate)2n=40,FN=52.G-带比较结果显示,2种麝鼩的G带很相似,进一步表明该属动物的核型进化很保守;两者核型差异在于:灰麝鼩6号染色体是亚端着丝粒染色体,它与白尾梢麝鼩6号端着丝粒染色体以及19号端着丝粒染色体近着丝粒部分的带型相一致.
麝鼩属;白尾梢麝鼩;灰麝鼩;核型;G-带
麝鼩属(Crocidura)隶属食虫目鼩鼱科(Insectivora:Soricidae),是哺乳动物中最大的一个属,共有180余种,广泛分布于非洲区、古北区和东洋区[1].该属1/3以上的物种都有染色体核型报道,其染色体数目从2n=22到2n=68不等,是哺乳动物中染色体数目变化最大的属之一[2].但是麝鼩属动物的核型国外研究较多,比如Grafodatsky等[3]曾报道了5种麝鼩属动物的核型,并研究了其中几个物种的G-带和C-带;Biltueva等[4]也曾对麝鼩属5个物种的G-带核型进行了比较,研究结果表明,罗伯逊易位、串联融合以及整条染色体的易位是造成这几个物种核型差异的主要原因.而我国麝鼩属动物的核型研究起步较晚,迄今为止,国内期刊上有关我国麝鼩核型的文章共2篇,涉及2个种[5,6].本文着重研究来自于云南省的白尾梢麝鼩(C.fuliginosa)和灰麝鼩(C.attenuate)的常规核型和G-带核型,旨在为研究其种属间进化关系提供一定的理论依据.
1 材料与方法
1.1细胞培养
本实验所用细胞均为成纤维细胞(组织材料分别来源于耳朵、肺、心脏等部位),细胞培养按照常规的方法进行.均采用DMEM添加10%-15%的新生牛血清(NCS,Hyclone)或者胎牛血清(FBS,杭州四季青公司)、青霉素100 μg/mL、链霉素100 μg/mL,于37℃,5% CO2下培养.
1.2染色体标本制备
终止培养前,用10 μg/mL BrdU处理5 h,之后用0.025-0.05 μg/mL的秋水仙素处理30-60 min.中期染色体的制备按常规方法进行,包括用0.075M KCl溶液低渗20分钟,用甲醇∶冰乙酸(3∶1)固定液固定2~3次,最后悬液置于-20℃保存.采用常规空气干燥法进行染色体制片.
1.3 G显带
按常规胰酶-Giemsa法进行G-带显示.将染色体玻片标本在65℃烤箱中老化3-12 h,取出冷却至室温后,用0.005%胰酶处理5-7 min(不同物种胰酶处理时间有所不同),然后将染色体玻片标本置于2×SSC中,终止胰酶反应,最后用Giemsa染液(Giemsa原液:0.025M KH2PO4缓冲液=1:6稀释,PH 7.0)染色5分钟,清水冲洗干净,室温下晾干后即可在显微镜下检测.对染色体显带和分散较好的分裂相用CytoVision System (Applied Imaging) 软件拍摄并进行图像分析.
1.4染色体命名
白尾梢麝鼩的染色体分类按Levan等[7]的标准.根据染色体的长度、着丝粒位置和带型将染色体配对、排序,得到常规核型和G-带核型.灰麝鼩的常规核型和G-带核型根据Motokawa等[6]的报道排列.
2 结果
白尾梢麝鼩的染色体数为2n=40,常染色体臂数FN=50,与先前的报道相同[5],包括3对中或亚中着丝粒染色体(1~3号染色体),3对亚端着丝粒染色体(4、5、7号染色体),13对端着丝粒染色体(6及8~19号染色体);X染色体是相对较大的亚中着丝粒染色体,Y为端着丝粒染色体(图1:A).
灰麝鼩的染色体数目与先前报道的台湾产灰麝鼩相同[6],也是2n=40,FN=52:包括3对中或亚中着丝粒染色体(1~3号染色体),4对亚端着丝粒染色体(4~7号染色体),12对端着丝粒染色体(8~19号染色体);X染色体是相对较大的亚中着丝粒染色体,Y为端着丝粒染色体(图1:B).灰麝鼩的核型与白尾梢麝鼩的基本相同.

3 讨论
Motokawa等[8]曾报道了分别来自于台湾、广东的灰麝鼩的核型和G-带,并依据这两地的灰麝鼩核型存在差异,建议将来自台湾的灰麝鼩更名为C.tanakae,而产自广东的灰麝鼩(C.attenuata)则保留该物种名.然而本研究中的云南产灰麝鼩却与台湾产灰麝鼩的染色体数目相同,均为2n=40,常染色体中含有3对中或亚中着丝粒染色体,4对亚端着丝粒染色体,12对端着丝粒染色体,常染色体臂数FN=52,G-带核型也基本一致.但是与广东产灰麝鼩的染色体数目明显不同[8].Motokawa等[8]的研究表明,广东产灰麝鼩的二倍体染色体数目存在多态性,具有35、36和38三种类型,但是这三种类型的灰麝鼩的常染色体臂数相同,FN=50,三者之间的核型差异是由于罗伯逊易位所造成的.因此,Motokawa等[8]提出将台湾的灰麝鼩更名为C.tanakae的建议有待商榷.
白尾灰麝鼩与灰麝鼩形态相似,存在同域分布现象[9],并且都属于中等大小麝鼩,这给分类工作带来极大的困难.本研究中的白尾灰麝鼩和灰麝鼩样品是在同一地点采集的,两者的染色体数目相同,均为2n=40,但常染色体臂数不同.白尾灰麝鼩的常染色体臂数FN=50,而灰麝鼩FN=52,即多一对双臂染色体,据此可轻易将两者区分.
麝鼩属动物的常规核型研究表明,该属的二倍体染色体数目在22~68之间,常染色体臂数从34到86不等[2].Biltueva等[4]曾对麝鼩属中的Crociduradsinezumi(2n=40)、Crocidurawatasei(2n=26)、Crociduragueldenstaedtii(2n=40)、Crociduraleucodon(Georgia,2n=28)以及Crociduraleucodon(Czech Rep,2n=28)这几个物种的G-带核型进行了比较,研究结果表明麝鼩属动物的染色体具有高度的保守性,物种之间的带型很相似.本文中白尾梢麝鼩和灰麝鼩这二个物种的G-带比较结果也证实这一观点(图2).从图2中可以看出,这2个物种的1~5号染色体、7~18号染色体以及X染色体的G带带型基本相同.两者的G-带核型差别在于:灰麝鼩6号染色体是亚端着丝粒染色体,它与白尾梢麝鼩6号染色体以及19号染色体近着丝粒部分的带型相一致.
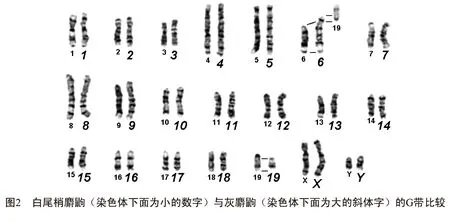
上述有关麝鼩属动物具有保守的核型的观点,仅是依据物种间的G带比较所做出的推论,仍需要进行荧光原位杂交(fluorescenceinsituhybridization,FISH)研究加以验证.
[1]Hutterer R.Order Soricomorpha.In:Wilson DE,Reeder D,(eds),Mammal species of the world:a taxonomical reference.Baltimore:John Hopkins University Press,2005:220-311.
[2]Zima J,Lukacova L,Macholan M.Chromosomal evolution in shrews.In:Wojcik JM,Wolsan M,(eds),Evolution of Shrews.Bialowieza:Mammal Research Institute,Polish Academy of Sciences,1998:175-218.
[3]Grafodatsky AS,Radzhabli SI,Sharshov AV,et al.Karyotypes of 5 species of Soricidae-Crocidura in the fauna of the USSR.Tsitologiia,1988,30(10):1247-50.
[4]Biltueva LS,Rogatcheva MB,Perelman PL,et al.Chromosomal phylogeny of certain shrews of the generaCrociduraandSuncus(Insectivora).Journal of Zoological Systematics and Evolutionary Research,2001,39(1):69-76.
[5]王应祥,李崇云,陈宜峰.白尾梢麝鼩的染色体研究[J].遗传,1983,5(3):29-31.
[6]Motokawa M,Harada M,Lin LK,et al.Karyological study of the gray shrewCrociduraattenuata(Mammalia:Insectivora) from Taiwan.Zoological Studies,1997,36(1):70-73.
[7]Levan A,Fredga K,Sandberg AA.Nomenclature for centromeric position on chromossomes.Hereditas,1964,52(2):201-220.
[8]Motokawa M,Harada M,Wu Y,et al.Chromosomal polymorphism in the gray shrewCrociduraattenuata(Mammalia:Insectivora).Zoological Science,2001,18(8):1153-1160.
[9]Jiang XL,Hoffmann RS.Arevision of the white-toothed shrews (Crocidura) of Southern China.Journal of Mammalogy,2001,82(4):1059-1079.
2014-03-18
教育部科学技术研究重点项目(编号:212069);浙江省自然科学基金青年科学基金项目(编号:LQ12C04002).
蔡灵超(1992-),女,浙江台州人,主要从事动物染色体研究.
叶建平,E-mail:yejp02@126.com.
Q953
A
1008-293X(2014)08-0072-04
猜你喜欢
中学化学(2024年4期)2024-04-29 22:54:35
生物技术进展(2022年1期)2022-02-11 03:45:46
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
科学之谜(2018年9期)2018-12-17 09:05:08
中国民族医药杂志(2016年5期)2016-05-09 07:43:50
生物学教学(2016年12期)2016-04-10 23:04:28
作文大王·低年级(2016年3期)2016-03-11 00:48:53
哈尔滨医药(2015年2期)2015-12-01 03:57:21
中国当代医药(2015年9期)2015-03-01 02:02:12
遗传(2015年4期)2015-02-04 06:29:22
