
Diversity patterns and palaeobiogeographical relationships of latest Devonian-Lower Carboniferous foraminifers from South China: What is global,what is local?
2014-09-27 08:05MarkusAretzEliseNardinDanielVachard
Journal of Palaeogeography 2014年1期
Markus Aretz , Elise Nardin Daniel Vachard
1.Géosciences Environnement Toulouse (GET), Observatoire Midi Pyrénées, Université de Toulouse, CNRS, IRD, 14 Avenue E.Belin, F-31400 Toulouse, France
2.Université Lille 1, UMR 8217 Géosystèmes, Bâtiment SN5, Avenue Paul Langevin, F-59655 Villeneuve d’Ascq cedex, France
1 Introduction
Foraminifers with a calcareous wall (i.e., of the class Fusulinata sensu Vachardet al., 2010)are among the most abundant and widely distributed organisms in latest Devonian and Carboniferous strata (e.g., see references in Vachardet al., 2010, Somervilleet al., 2013).Their abundance and rapid phylogenetic changes make them essential elements for biostratigraphic schemes of the latest Devonian and Carboniferous on local to global scales (e.g., Mamet and Skipp, 1970; Cózar, 2003; Potyet al., 2006; Cózaret al., 2011; Hanceet al., 2011).Although the amount of data for foraminifers with agglutinated walls (Kaminskiet al., 2010)is increasing, they remain unstudied in many regions, and thus are excluded in the following study.
Southern China illustrates well the importance of calcareous foraminifers for global correlation in the Carboniferous.The Global Stratotype Section and Point (GSSP)for the Tournaisian-Visean boundary situated at Pengchong in Guangxi Zhuang Autonomous Region (Devuystet al., 2003; Houet al., 2011)is so far, the only Devonian and Carboniferous GSSP not based on the first appearance dutum (FAD)of a conodont, but on a calcareous foraminifer (Eoparastaffella simplex).The GSSP is one result of the fruitful collaboration between Belgian and Chinese institutions (e.g., Conilet al., 1988; Coenet al., 1996;Hanceet al., 1997; Aretzet al., 2012).This collaboration is certainly highlighted by the recently published atlas on Strunian-Visean foraminifers from South China (Hanceet al., 2011), which summarizes the work of two decades.
Although calcareous foraminifers are important for biostratigraphy, their palaeobiological requirements and limitations are currently often neglected.The relative facies dependencies of taxa have been described in several studies (e.g., Gallagher, 1998 or models in Vachardet al.,2010).Although the in fl uence of factors like grain-size and texture is indisputable, these information data are rarely taken into consideration in biostratigraphic discussions.The same is true for other palaeobiological factors like life mode.All Palaeozoic calcareous foraminifers were benthic, with epibenthic and endobenthic modes as well as vagile and attached forms.These life modes clearly infl uenced the distribution patterns, and among others, the dispersion of the different foraminifers (species or morphological groups)is crucial for our understanding of patterns in deep time.Alve (1999)listed four principal possibilities for dispersion: release of propagules following reproduction, meroplanktonic life stage, self locomotion and passive entrainment and transport of growth stages.The effective distances for dispersal are very different and range from much localized (self locomotion)to larger distances (e.g., zygotes and embryonic juveniles, which may be pseudoplanktonic to planktonic for several days).The many taxa in the Upper Devonian and Lower Carboniferous with wide palaeogeographical distribution indicate effi cient quick dispersal, which supports the biostratigraphical usefulness of calcareous foraminifers.In fact, due to the benthic, and often endofaunal mode of life of Early Carboniferous foraminifers, and the absence of demonstrated sexual reproduction during this period (Vachardet al., 2010), self locomotion, passive transport by accelerated bottom currents, or rafting with macroalgae lifted by strong streams or hurricanes, are most likely.
Strunian-Serpukhovian calcareous foraminifers have been intensively studied over the last fi fty years and there are hundreds of papers describing foraminifers from almost all palaeocontinents.Although the community of workers has been very productive, surprisingly little has been done on the quanti fi cation of diversity or palaeogeographical patterns.A few exceptions can be found for di-versity plots, ine.g.Groves and Wang (2009), but overall,the interest in Upper Carboniferous-Permian fusulinids overshadows the older forms and time slices (e.g., Leven,1993; Groves and Wang, 2009).Bio-events impacted the abundance and evolution of foraminifers, but the clear separation of assemblages for specif i c time intervals (e.g.,three assemblages for the three Visean substages)does not necessarily correspond to global bio-events (Vachardet al., 2010).Groves and Lee (2008)in referring to the data of Rauser-Chernousovaet al.(1996)and Mamet (1977)estimated that 66% of all Mississippian foraminiferal genera were cosmopolitan.This number supports the idea of relatively few large palaeobiogeographical realms for Upper Devonian-Lower Carboniferous foraminifers (see below).However, more regional models show distinct foraminiferal assemblages, and thus allow the separation of a number of provinces or other regional subunits within the photic, inner carbonate platform realms (e.g., Xu, 1987;Kalvoda, 2002; Somervilleet al.,2013).
Early Carboniferous calcareous foraminifers have essentially three differences from the modern foraminifers(Vachardet al., 2010), that complicate much of the palaeobiogeographical analyses and comparisons: (1)they are almost all endofaunistic; (2)they occupy only the upper parts of the inner carbonate platforms; (3)their dominant,endothyrid-type of coiling is particular and does not exist in modern times.As underlined above, the study of the agglutinated foraminifers (i.e., the class Textulariata)would be very interesting for deeper and/or cooler waters (e.g.,reddish or pink nodular limestones), but there are many palaeobiological, microstructural, and taxonomical problems with the Devonian-Carboniferous taxa assigned to Textulariata (Vachardet al., 2010).
In this study, we use data compiled for Strunian (latest Devonian), Tournaisian, Visean and Serpukhovian (Lower Carboniferous)foraminifers from southern China to discuss diversity patterns and palaeobiogeographical affinities of these forms.It is the aim to discuss and identify the inf l uencing factors and to test if the diversity patterns in South China are comparable to those from other palaeocontinents or to which degree a local signal is preserved in these data.
2 Setting
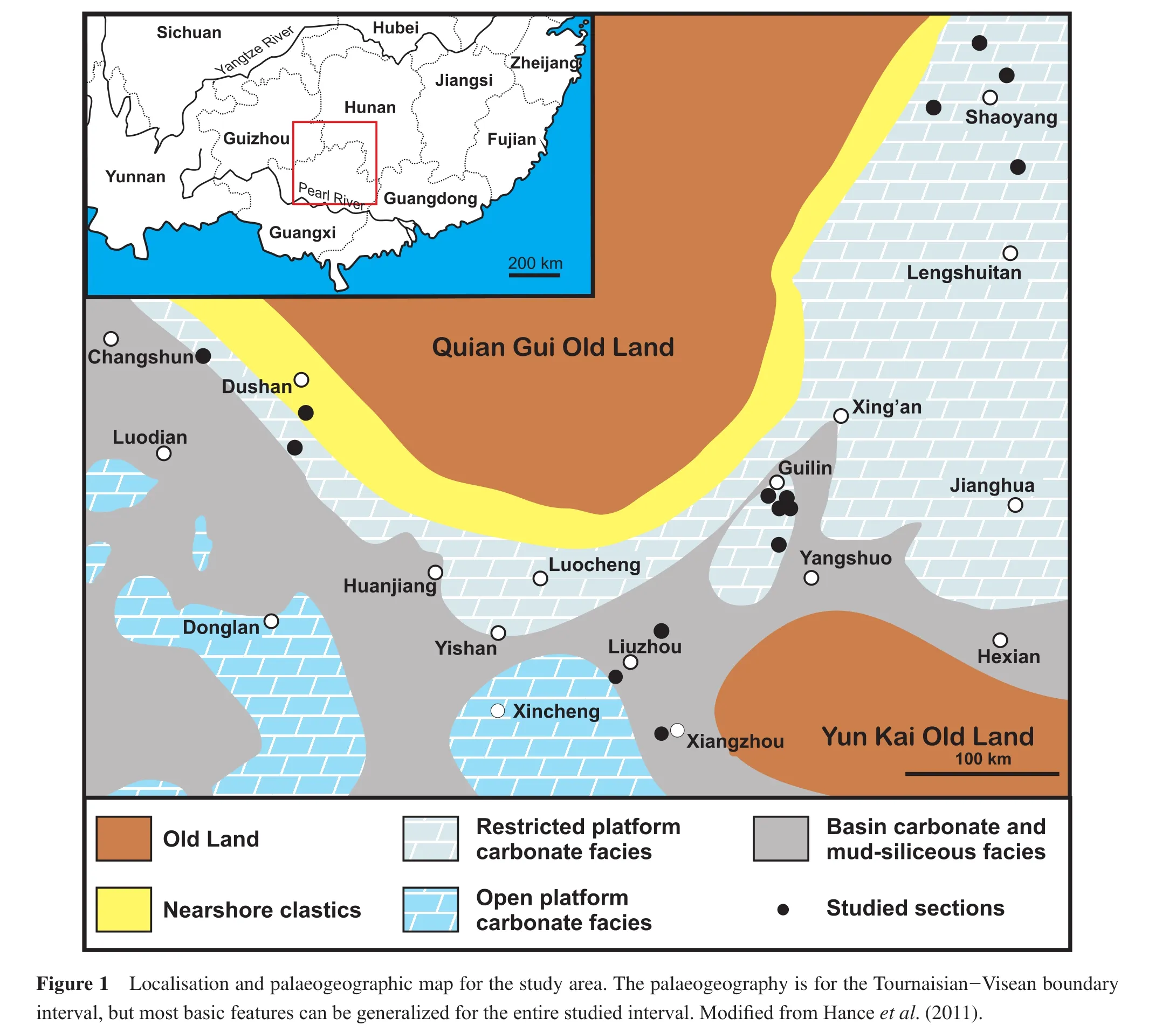
During latest Devonian-Early Carboniferous times, the South China Block was located just north of the palaeoequator in a key position between the eastern Palaeotethyan Ocean and the Panthalassa Ocean (e.g., Blakey, 2007).Characteristic for the South China Block is a differentiated palaeogeography resembling a vast tropical archipelago(Figure 1).Landmasses (Old lands in the Chinese nomenclature)consolidated and deformed during Early Palaeozoic times are surrounded by carbonate platforms, which are either directly attached to the landmasses (numerous terrigenous clastic interbeds)or isolated from the land and situated above intra-basinal highs (without terrigenous clastic interbeds).Narrow intra-platform basins, in which detrital mud and siliceous strata dominate, separate and dissect the carbonate platforms.Moreover, as indicated by Hanceet al.(2011), notable breaks of slope affect the intra-platform basins, and input of shallow- water components into the basins by calciturbidites and debris f l ows can be seen in intercalated limestones in the basinal facies(Jinet al., 2007).On the other hand, the distribution of these major facies realms can f l uctuate and be modif i ed by a series of transgressive-regressive cycles, due to local tectonic movements and/or global glacioeustatic movements.
In consequence of this complex palaeogeography, different lithostratigraphic subdivisions (Figure 2)have been established not only for specif i c facies realms, but also for the various provinces and regions in China (Hunan, Guangxi and Guizhou in our study area).Hanceet al.(1997, 2011)have proposed correlations between the different facies realms and regions, but so far they are tentative and not yet formalized.The application of the biostratigraphic scheme of Potyet al.(2006)to the study area has been proven successful (Hanceet al., 2011; Potyet al., 2011)and allowed some very detailed biostratigraphic correlations.Currently, these are restricted to specif i c sections, and bearing in mind the large extent of Carboniferous strata in southern China, further studies are required to foster correlations between regions and provinces, but also within them.
Hanceet al.(2011)presented an example for the very different facies and thicknesses encountered around the Tournaisian-Visean boundary in Guangxi, even in neighbouring sections in the same facies realm of the same carbonate platform.Thus, the large facies realms and the use of few formations names should not hide the facies variability on a smaller scale, which also comprises specif i c facies types for few outcrops (e.g., mud mounds and reefs;Jinet al., 2007; Aretzet al., 2012).
3 Data and methods
3.1 Data sources
A Microsoft Excel spreadsheet has been compiled for Strunian (latest Devonian), Tournaisian, Visean and Serpukhovian (Early Carboniferous)calcareous foraminifers from key sections in southern China.The spreadsheet contains 420 species assigned to 155 genera.The main source for the data is the atlas of Hanceet al.(2011).This limitation was chosen for two reasons: (1)the atlas is the most recent comprehensive taxonomic work with the most detailed biostratigraphic correlation, which takes into account the previously published studies of Chinese workers, and (2)the taxonomic concept is homogeneous, and thus the bias due to different taxonomic concepts should be largely reduced.
All taxa mentioned and described in Hanceet al.(2011), their given stratigraphic ranges (foraminiferal zones DFZ and MFZ for the Devonian and Mississippian, respectively)and palaeobiogeographical information have been extracted.If necessary, missing information,especially palaeobiogeographical information data were added, based on literature researches and our own unpublished data.For the purpose of the study, eleven palaeobiogeographical units were differentiated (see below).These units correspond to well-studied areas and have already been regrouped in previous studies (e.g., Lipina, 1973;Kalvoda, 1990, 2002; Okuyucuet al., 2013).However,future studies will have to further subdivide these units,but in the current state of play, the data for regions like Far East Russia are too heterogeneous and need to be revised first.The atlas contains relatively few data on latest Visean and Serpukhovian foraminifers, because Hanceet al.(2011)did not study many sections covering this time in-terval.The currently best-known section for shallow-water carbonates of the latest Visean-Serpukhovian in South China, the Yashui section, Guizhou Province, was only preliminary studied and sampled by Hanceet al.(2011).To partly close this gap, the recent data on the Yashui section (Groveset al., 2012)were integrated into the dataset,but the taxonomy was corrected according to the concept used in the atlas.Then all data were controlled and some mistakes corrected by one of us (D.V.).

3.2 Time units
The exact lengths of the foraminiferal biozones (DFZ,MFZ)have not yet been calculated.They represent time units of unequal lengths.In order to compare the diversity trends, more chronological precision is needed.An attempt is made herein to estimate the duration of each biozone(see Figures 4-10).It uses (i)the dates for the base of the stages from the Geological Time Scale (GTS)2012 (Gradsteinet al., 2012), (ii)the estimated lengths of 3rd-order sequences of Hanceet al.(2001)correlated to the biozones of Potyet al.(2006), (iii)an unpublished attempt for the length of coral biozones in the same interval, and (iv)sediment thicknesses observed in the time intervals of the biozones.The GTS dates have been used as f i xed points, but these dates may underestimate the duration of the Tournaisian and Visean.For simplif i cation of the text, the biozone MFZ 4 is attributed to the Hastarian, although its upper part extends into the Ivorian (Potyet al., 2006, in press).This simplif i cation can be justif i ed with the unsuitability of facies for foraminifers in the basal Ivorian (Potyet al.,in press)and the correlation of MFZ 4 to the Maurenne Formation (Potyet al., 2006)in the Namur-Dinant Basin.The latter is late Hastarian in age.
3.3 Geographical units
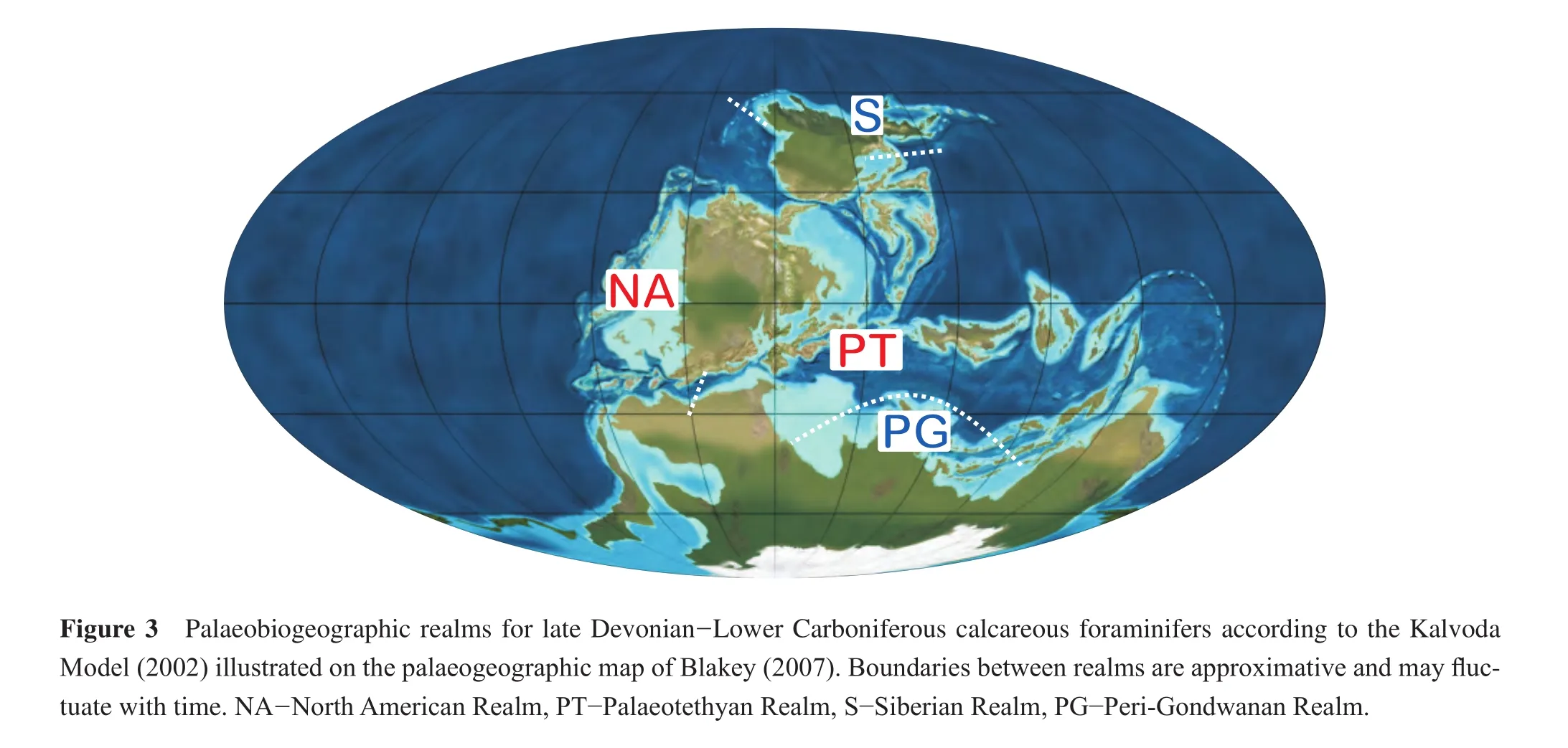
The data used in this study are all from the South China Block situated in the north-eastern Palaeotethyan Realm.In order to discuss the relationship to near, middle and far distanced regions, the South Chinese data have been compared to ten other palaeobiogeographical units (Figures 7-11).The palaeogeographical positions in respect to South China and the availability of data were an essential element for the definition of these units.A set of units comprise the immediate palaeogeographical neighbours of South China (SE Asia, Western Australia, Far East Russia and neighbouring countries).More distant in the Palaeotethyan Realm were Turkey and Iran (Arabian Plate excluded), the Russian Platform including the Urals, Western and Central Europe and Northwest Africa.To test relationships to palaeogeographically more isolated regions, three units have been added; Eastern Australia (eastern margin of Gondwana), Japan (Panthalassa Ocean)and North America including Alaska and northern Canada.The latter region may indicate closer palaeobiogeographical connections than traditionally seen for the North American Realm.
3.4 Diversity dynamics and similarity analyses
Diversity measures are expressed in terms of the total diversity per capita (total number of genera or species recorded in a time interval divided by the estimated length of the interval; see Cooper, 2004)applied to the matrix constituted by the observed occurrences of the taxa (genus or species)in the fossil record and to the matrix constituted by the theoretical range of the taxa.Observed diversity is supposed to be more affected by sampling and effort biases in addition to the potential presence of Lazarus taxa(Cascales-Miñanaet al., 2013).Therefore, most of our reasoning will be based on the theoretical diversity per capita.
Evolution measures are based on metrics described by Foote (2000a).Per-taxon rates have been selected to normalize the number of originations and extinctions by the total diversity per capita.Those rates were the basis for the calculation of the turnover and diversif i cation rates (Nardin and Lefebvre, 2010).
Palaeobiogeographical closeness was evaluated using similarity and ordination methods.Cluster analyses used the dissimilarity distance Raup-Crick method, the similarity distances Euclidean and Jaccard (Raup and Crick,1979)accompanied by the agglomerative nesting with group averaging methods (Sokal and Michener, 1958)or the distance Sørensen associated to the Ward linkage method (Ward, 1963), to respect the concavity (McCune and Grace, 2002).The Nonmetric Multidimensional Scaling (NMDS)ordination method was also based on Raup-Crick distance measures to verify the expected palaeobiogeographical relationships (Clarke, 1993).All statistical analyses were computed with the libraries cluster, vegan,stats for R2.9.1.5 (R.Development Core Team, 2009).
4 Palaeodiversity
4.1 Global diversity
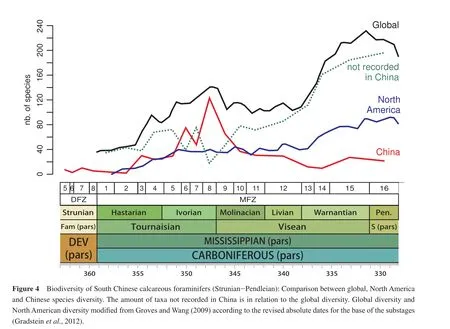
Although calcareous foraminifers are intensively studied, quantitative diversity data are rare.On the genus level,data are available from Sepkoski (2002)who compiled the stratigraphic and taxonomic data from Loeblich and Tappan (1987).Groves and Lee (2008)provided a first curve at the species level when discussing originations and extinctions of foraminifers during the Late Palaeozoic Ice Age.The database was expanded subsequently, and Groves and Wang (2009)presented two curves, one for the diversity of North American foraminifers and a second for the global diversity.The difference between the two curves should ref l ect the diversity in what the authors called the Arctic-Eurasian association, and thus show the diversity for the Arctic-Eurasian realm (see section 5.1).
The methodological approach for the analysis was the same as in this study of southern China, but the data of the Groves and Lee (2008)and Groves and Wang (2009)studies have not been calibrated to biozones, but to 1 Ma intervals.This calibration was achieved by graphic correlation, which used as an absolute reference time scale the dates given by the GTS 2004 (Gradsteinet al.,2004).However, the updated version of this chart (Gradsteinet al., 2012)results in new dates for the base of the Visean and Serpukhovian stages.In assuming that the total duration of the stage changed equally over the entire length of the stage, the Groves curve has been corrected for the Mississippian to enable a better comparison to be made to the results obtained in southern China (Figure 4).
Groves and Wang (2009)noted an overall diversity increase for the Carboniferous, mainly based on the radiation of fusulinoideans in the Pennsylvanian.The Mississippian curve can be described as a double bell-shaped increasing line, with the two maxima in the late Tournaisian and late Visean (Figure 4).Somerville (2008)described a very similar curve for the diversity of western European foraminifers (genus level)with two maxima; the first in the Chadian (basal Visean).Groves and Wang (2009)explained the initial low diversity of the latest Devonian and early Tournaisian was explained as the earliest phase of recovery after the Frasnian/Famennian extinction crises.
4.2 Observations in South China
4.2.1Diversity
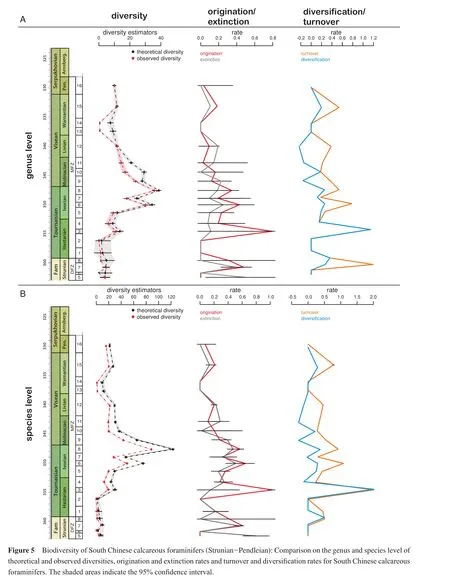
The diversity of calcareous foraminifers, which is basically the expression of the ratio of originations and extinctions, has been calibrated to the duration of the foraminiferal biozones of the Strunian to Pendleian (DFZ 5 - MFZ 16).Foraminiferal species and genus diversity show the same overall pattern for observed and theoretical diversity(Figure 5), and the latter is used in the following.
Singletons (taxa only found in a single biozone)have been included in the analyses contrary to the recommendation of using only boundary-crossing taxa (Foote, 2000a,2000b).This is based on the analysis that the datasheet contains 71% singletons on the species level (299 species)and 41% singletons on the genus level (64 genera).If it assumed that the presence of a taxon coincides with the duration of the biozone in which it is recorded, a maximum theoretical life time for each taxon can be calculated.The average duration of a species is 3.66 Ma (0.8-22.4 Ma),and those for a genus is 6.27 Ma (0.8-24.4 Ma).Long life times are rare among species, only 10% are known in more than 3 biozones.For genera the number rises to 30%.These values can be positively interpreted to ref l ect the fast evolutionary rates for calcareous foraminifers.A more conservative interpretation could evoke taxonomic oversplitting due to concentration on specif i c biozones or too poorly defined intraspecif i c variabilities.
For the studied time interval, the diversity curve is bellshaped (Figure 5).Diversity in the Strunian and Hastarian is low, and the extinctions events of the latest Devonian (Hangenberg Crisis)are visible in a slightly lower diversity in the early Hastarian (MFZ 1-2)compared to the Strunian (DFZ 5-8).Recovery takes the entire early Hastarian, but then markedly accelerates within the late Hastarian.In biozone MFZ 3, the numbers are as twice as much as in the Devonian.The Ivorian is characterised by an important diversity increase culminating in its last biozone (MFZ 8).Two of the three highest foraminiferal species diversities observed in the studied interval are found in this substage (MFZ 6, MFZ 8).The peak in MFZ 6 corresponds to the arrival of genera, such asDainella,EoforschiaandSpinobrunsiina; MFZ 8 is characterised by the arrival of new genera and important originations in the existing stock.The third highest species diversity is found just at the base of the following substage (MFZ 9)and initiates in South China the diversity decrease of the Visean and Serpukhovian.The genus diversity of the Visean-Serpukhovian interval follows the same trend, but the peak values are more evenly distributed between the Ivorian and Molinacian.
4.2.2Evolutionary rates
Origination, extinction, diversif i cation and turnover rates were calculated per time interval (Figure 5).On the genus and species level, the origination and extinction curves are relatively parallel, indicating that times of high origination are also characterised by times of high extinctions.The same can be postulated for times of low origination rate, when extinction rate is low as well.There is one signif i cant exception of the parallelism in the top of the Devonian during the extinction events of the Hangenberg crises (top DFZ 8).The shape of the curves for origination and extinction rates resembles the diversity curve, when the overall diversity is low.Although the high peaks in the diversity correspond to elevated rates of origination and extinction, the highest amplitudes of these rates correspond to the rapid diversity increase of the late Hastarian,which in absolute numbers is less important than those of the Ivorian.It is evident that for the Tournaisian the origination rate exceeds the extinction rate, however the drop in total diversity in the Visean and Serpukhovian is not due to a constant higher extinction rate.Rates are rather variable,and thus the diversity curve shows several small-scale ups and downs in the younger intervals.This is also seen in the diversif i cation rate, which f l uctuates around 0 for most of the Visean and Serpukhovian.The diversif i cation rate in the Strunian and Tournaisian is in generally higher, which not surprisingly results in the bell-shaped diversity curve.
On the genus level, the highest turnover rates are found in the Strunian (DFZ 7)and in the late Hastarian (MFZ 3)(Figure 5A).Only the latter peak is found on the species level (Figure 5B), indicating that the diversity peak in the DFZ 7 results mainly from the origination of genera.Species origination contributed only subordinately to the Strunian diversity increase, because even high origination rates were levelled out by a high extinction rate.The Hangenberg Crises seems to be mainly a problem of low origination and not of extinction, when only using the data for the DFZ 8-MFZ 1 interval.However, DFZ 8 is poorly known and when enlarging the focus, the net extinction and high turnover in the latest Devonian, just below the Devonian-Carboniferous boundary are well illustrated.This overall trend completely changes in the late Hastarian when higher origination rates result in high diversif i cation and turnover rates.
4.3 Discussion
The regional North American diversity curve shows the same general trends as the global diversity curve (Groves and Wang, 2009; Figure 4).The main difference between these curves is the amplitude of changes.The same is true for the Arctic-Eurasian curve (difference between global and North America values).The diversity curve for southern China does not show the same parallelism as these three other curves.It shares the diversity increase in the Tournaisian, and also shows peak values in the late Tournaisian, but the typical diversity increase of the Visean and the high values around the Visean-Serpukhovian boundary of the Groves and Wang curves have not been found.Only 10% of the global diversity is recorded for the late Visean and Serpukhovian in southern China, which is remarkably low compared to the average of 35%-40% for other Mississippian substages.This is even more surprising because in many other regions in Eurasia, the late Vi-sean is a major time period for foraminiferal diversif i cation (e.g., Europe: Conil and Lys, 1964; Vachard, 1977;Cózar and Somerville, 2004; North Africa: Vachard and Fadli, 1991; Vachard and Tahiri, 1991; Cózaret al., 2011,2013; Somervilleet al., 2013).
Hence, such a decoupling from the general trend may have palaeoenvironmental, palaeobiological or geological reasons.A major geological event can be excluded here,since South China was not affected by major tectonic or geodynamic events, such as the Variscan Orogeny, and remained in tropical latitudes during the Carboniferous.Palaeobiological factors can be excluded also, because no important change in the life strategies of calcareous foraminifers is known in the Mississippian (e.g., Groves and Lee, 2008; Vachardet al., 2010).More likely are palaeoenvironmental changes.High-frequency sea-level f l uctuations common in the late Visean and Serpukhovian, often related to major glaciations in southern Gondwana (e.g.,Hanceet al., 2001; Late Palaeozoic Ice Age), may have increased pressure on habitat stability.Habitats may have become more instable, but this should not necessarily result in a diversity decrease.According to Groves and Lee(2008), it could be even the opposite, because foraminifers show higher origination rates in times of habitat instability probably resulting in a diversity increase.This is seen on the carbonate platforms in other Eurasian regions (see above), where calcareous foraminifers were facing the same environmental pressures as in South China.
Hence, other factors should contribute to the low Warnantian-Pendleian diversity in South China.One factor was already mentioned previously (section 3), Hanceet al.(2011)studied signif i cantly fewer outcrops in upper Visean and Serpukhovian strata than in older substages, making the dataset less complete for these time slices.It should be noted that Hanceet al.(2011)only correlated strata to the lower Warnantian, but did not directly find MFZ 13 and 14, which is the prime reason for the Chinese diversity low in the lower Warnantian.This can be at least partly explained with a facies problem.In some regions in South China, the Warnantian strata are characterised by the presence of widespread terrigenous clastic facies (Figure 2)and locally peloidal, micritic and even stromatolithic facies (Shen and Qing, 2010).These unsuitable facies for calcareous foraminifers reduced the potential habitat surface on the platform systems.This certainly contributed to the low diversity in the Warnantian in South China, further accentuated by the generally poor knowledge of the late Visean and Serpukhovian in large regions such as central and northern Guangxi.
The opposite effect can be seen at the diversity peak in the late Tournaisian when comparing the global and Chinese diversity curves.In the latest Ivorian, the diversity curve for South China almost attains the values of the global diversity curve, when 87% of all known species of this biozone are recorded in South China.This value is more than twice the normal average of 35%-40% for wellstudied biozones in the Ivorian, Molinacian and Livian.Due to the search for the GSSP for the base of the Visean,the calcareous foraminifers from the upper Tournaisian and lower Visean strata of South China have been studied in great details (Hanceet al., 1997, 2011; Devuystet al.,2003; Devuyst, 2006; Houet al., 2011).The associations in different facies and different time slices (biozone and subzones)are very well known and documented.This very detailed knowledge is rather unique.In the two decades,more detailed studies of the same time interval have been published for Europe (Devuyst and Kalvoda, 2007; Kalvodaet al., 2010, 2012).Those studies further increase the number of species known in this time interval, but these data are too recent to have been integrated in the curve of Groves and Wang (2009).Thus, with an updated global curve, the distance between the global and the Chinese curves should increase.
The low diversity in the early Hastarian is a combined effect of palaeobiological and environmental factors.Most of the complex and large forms (e.g., Quasiendothyridae)have become extinct at the Devonian-Carboniferous boundary in the Hangenberg Crises.The first biozone of the Mississippian contains mainly the name-giving small unilocular foraminifers.Thus, a ‘Lilliput Effect’ could be proposed in the scenario for the aftermath of the Hangenberg extinction, since the abundance of unilocular foraminifers and the general size relations at the base of the Hastarian are also known outside South China (e.g., Belgium: Potyet al., 2006; and references in Vachardet al.,2010).Recovery took apparently as long as the lower Hastarian (MFZ 1, 2), before a rapid diversity increase is seen in the late Hastarian (MFZ 3)with a major diversif i cation in the Tournayellidae, Septabrunsiinidae and Chernyshinellidae.This long recovery phase and the following rapid diversity increase in South China are a regional signal.The reappearance of plurilocular foraminifers in MFZ 2 (Potyet al., 2006)is poorly documented in South China (Hanceet al., 2011)and it only starts in this region in zone MFZ 3.Thus, the diversity increase in South China is delayed for about one biozone and steeper compared to regions like Western Europe.However, during MFZ 2 time interval a facies problem existed.The carbonate factory was largely stopped and marly and shaly marine facies dominated on the platforms.These were unfavourable conditions for calcareous foraminifers, which contribute to the poor record of Hanceet al.(2011).The deposition of this facies on the platforms is generally attributed to the transgression of the Lower Alum Shale Event (Siegmundet al., 2002).This global event was largely modif i ed on the regional scale,but the reason why it impacted much more severely on the foraminiferal associations in South China than in other regions is not known.
This example of facies impact is probably the most extreme in the studied interval.However, the inf l uence of shale content, carbonate textures, grain sizes,etc.on the distribution patterns of particular calcareous foraminifers has already been mentioned.It cannot be neglected that a bias is introduced.The facies dependency of particular taxa should only become signif i cant when a limited number of samples from a limited number of sections and facies have been studied.To better evaluate this potential bias, more detailed data are needed on the carbonate microfacies containing the calcareous foraminifers have been found in.In the present state, it can only be suspected that the Chinese data for MFZ 2, 10, 13-16 are biased through a low number of studied sections and facies.
Data for the global diversity of Strunian foraminifers have so far not been published, and such a curve is beyond the scope of this study.However, in order to compare the Chinese data, a datasheet has been compiled for calcareous foraminifers from the Devonian-Carboniferous boundary(DCB)interval in the Urals (Kulagina, 2013).It has to be stressed that only the data of Kulagina (2013)have been taken into consideration, which represent a new interpretation of foraminiferal evolution around the DCB in the Urals.The foraminiferal zones of Kulagina (2013)have been approximately correlated to the DFZ and MFZ zones of Potyet al.(2006).The DCB has been placed between the remnantQuasiendothyraBeds -Tournayellina beata pseudobeataZone (= DFZ 8)and theEarlandia minimaZone (MFZ 1),i.e.the biozone of unilocular and bilocular foraminifers affected by a Lilliput Effect, where plurilocular taxa are rare or absent.
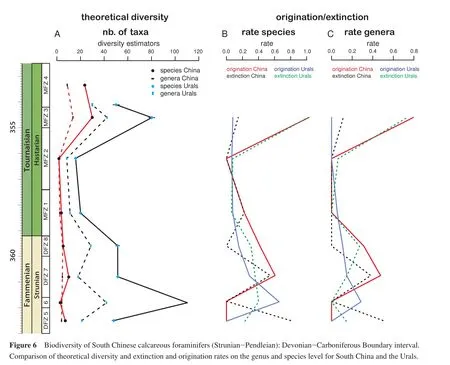
The curves for diversity and origination and extinction rates for the Urals and southern China (Figure 6)show in contrast to the Strunian only in the Hastarian a higher degree of parallelism.The amplitude and absolute values are always higher in the Urals than in South China (Figure 6A).Common to both regions is a low diversity time during the early Hastarian and the rapid diversity increase in MFZ 3.Within the Strunian the evolution of the diversity is antithetic (Figure 6B, 6C).The diversity peak of DFZ 6 in the Urals corresponds to a diversity low in South China, whereas the small diversity peaks in the Chinese DFZ 5 and 7 correspond to intervals of lower diversity in the Urals.Common to both regions is the subsequent diversity drop at the Devonian-Carboniferous boundary and the disappearance of Quasiendothyridea.It should be noted that in several sections quasiendothyrids are known in the basal beds of the Tournaisian (Kalvodaet al., in press).The discrepancy between the Urals and South China could have multiple origins.The translation of the Uralian biozones into the DFZ and MFZ scheme may have induced errors, but many important taxa for these two schemes occur in the Urals and South China and the parallelism of the Hastarian curves seems to indicate a correct correlation.Differences in the biodiversity could also be related to different treatment of morphological differences for the taxonomy and/or less studied material in morphological complex forms.The Uralian sections contain many species of genera likeSeptaglomospiranellaandQuasiendothyra.These genera are also found in South China, but with signif i cantly fewer species.The reasons could be palaeoenvironmental differences, palaeobiogeographical barriers, taxonomic over-splitting or insuff i cient sampling of specif i c time intervals or facies.Currently, none of these reasons can be excluded.Kulagina (2013)mentioned major differences in the composition of contemporaneous foraminiferal assemblages in the Urals related to facies differences.These local and regional differences may mask or accentuate the differences existing between the Urals and South China.In the future, more quantitative data are needed to decide if any of the two patterns is consistent with the global picture.
Groves and Lee (2008)and Groves and Wang (2009)were concerned about the impact of the Late Palaeozoic Ice Age (LPIA)on the origination and extinction rates of calcareous foraminifers.The idea that increased origination and extinction rates characterise glacial episodes(Groves and Lee, 2008)was not conf i rmed by the extensive analysis of Groves and Wang (2009), which took into consideration that the LPIA is much more complex than a single glaciation in the Pennsylvanian (Isbellet al., 2003;Fieldinget al., 2008).The data from southern China can only contribute to the discussion of the initial stages of the LPIA.A first glacial interval is today acknowledged in the late Famennian, but it is uncertain if it extends into the Tournaisian (e.g., Isbellet al., 2003; Issacsonet al.,2008)or not (e.g., Marshallet al., 2013; Potyet al., 2013).If the concept that a glacial climate should result in higher origination and extinction rates (Groves and Lee, 2008)is applied to the Chinese data, then apart from the lower Hastarian, the Strunian to lower Molinacian climate should be glacial.This is in contrast to any reconstruction for the Mississippian climate (see references above and Buggischet al., 2008).Therefore, the positive correlation of glacial climate and high origination rates is not followed herein,especially since the Chinese rates for the Tournaisian and early Visean are comparable to the contemporaneous rates proposed by Groves and Wang (2009).
5 Palaeobiogeographical relations
5.1 Palaeobiogeographical units
Global biogeography of late Devonian-Carboniferous calcareous foraminifers has been the focus of numerous qualitative studies (e.g., Mamet, 1977; Kalvoda, 1990,2002).The understanding of the palaeobiogeography of this time slice is often largely inf l uenced by concepts developed for Pennsylvanian and Permian times (e.g., Ross,1973; Groves and Wang, 2009).
The simplest palaeobiogeographical models are bipolar and differentiate a North American or Midcontinent-Andean Realm from a Tethyan Realm (e.g., Ross, 1973; Groves and Wang, 2009).Other models (Mamet and Belford, 1968;Lipina, 1973; Kalvoda, 1990)differentiate three realms: a subtropical/tropical Palaeotethyan Realm, a subtropical/tropical North American Realm and a northern boreal Siberian Realm.Kalvoda (2002)modif i ed the tripolar model in adding a southern boreal Perigondwana Realm comprising parts of the northern Gondwana margin (Arabia and neighbouring regions)and terranes in Afghanistan and the Himalaya region (Figure 3).The boundaries between the realms are not important palaeo(bio)geographical barriers,since otherwise the high degree of cosmopolitism of Mis-sissippian calcareous foraminifers (according to the distributions mentioned by Rauser-Chernousovaet al., 1996)would not be possible.The evaluation of the degree of faunal interchange between the realms is the main difference between the models.In any case, unless a quantitative global analysis for Mississippian foraminiferal biogeography has been performed the arguments for all models can not be validated, although the Kalvoda Model seems to be the most appropriate one.
One important difference between the two tropical/subtropical realms is the higher diversity of thick-walled, complex calcareous foraminifers in the Palaeotethyan Realm(Kalvoda, 2002; Groves and Wang, 2009).In this context,Groves and Wang (2009)tried to conf i rm the hypothesis of Mamet (1977), indicating that the Palaeotethyan Realm was the biogeographical centre of origination and radiation of almost all the foraminiferal lineages during the Lower Carboniferous.Although the absolute number of taxa in the Palaeotethyan Realm was higher (see section 4.3; Figure 4), the uneven size of the two realms has to be taken into consideration.Absolute numbers are lacking for the potential habitat surface in the realms, but this surface seems to have been much larger in the vast Palaeotethyan Realms according to most palaeogeographical maps (e.g.,Blakey, 2007).Thus, a higher diversity compared to the North American Realm could be simply an expression of the larger size of the Palaeotethyan Realm.
Within the realms, smaller palaeobiogeographical units(provinces or subprovinces)have been differentiated (e.g.,Kalvoda, 1990, 2001, 2002; Somervilleet al., 2013).However, for the definition of these units the same problems exist as for the realms, but the smaller the size of those units is, the more important becomes the inf l uence of facies on the distribution patterns of the calcareous foraminifers.Hence, there is the risk that these units become independent from palaeogeographical features.
Three palaeobiogeographical provinces called South Province, Middle Province and North Province were identif i ed in the Chinese Carboniferous based on foraminifers,brachiopods and corals (Xu, 1987).The boundaries between these provinces are horizontal lines on the recent geography (34° and 42° latitudes), and thus do not necessarily represent former plate boundaries.The North Province is thought to represent the Boreal Realm (Xu, 1987),and seems to comprise the Chinese part of the Siberian Plate.The South Province representing the Tethys Realm(Xu, 1987)corresponds to the South China Block (Yang-Tse)and several peri-Gondwanan Terranes (e.g., Baoshan Block).The study area is a part of this Province.The Middle Province has a transitional character (Xu, 1987).It may correspond to the North China blocks (Sino-Korea).
5.2 Observations in South China
Three palaeobiogeographical provinces called South Province, Middle Province and North Province were identif i ed in the Chinese Carboniferous based on foraminifers,brachiopods and corals (Xu, 1987).The boundaries between these provinces are horizontal lines on the recent geography map (34° and 42° latitudes), and thus do not necessarily represent former plate boundaries.The North Province is thought to represent the Boreal Realm (Xu,1987), and seems to comprise the Chinese part of the Siberian Plate.The South Province representing the Tethys Realm (Xu, 1987)corresponds to the South China Block(Yang-Tse)and several peri-Gondwanan Terranes (e.g.,Baoshan Block).The study area is a part of this latter Province.The Middle Province has a transitional character(Xu, 1987).It may correspond to the North China (Sino-Korean)Block.
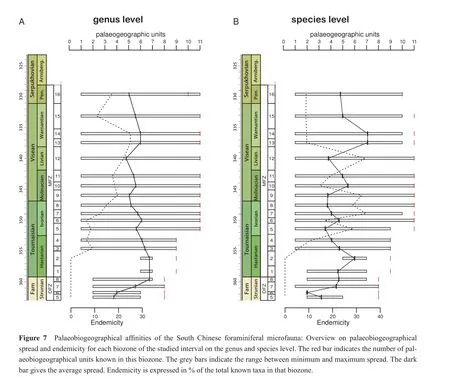
A first analysis (Figure 7)describes for all the 20 biozones the palaeobiogeographical spread of a taxon in the 11 units and the endemicity.The theoretical total diversity was used herein to partly compensate for the lack of data for biozones in the Strunian, Hastarian and Warnantian (Figure 4).The results for both taxonomic levels are comparable.The number of palaeogeographical units from which calcareous foraminifers have been described raises units from the Strunian to the Ivorian from an initial 8 to 11 units.Then, a plateau is reached, and only in the Pendleian the number decreases due to the absence of calcareous foraminifers in Eastern Australia.
Strunian and lower Hastarian taxa are geographically widespread, and no endemic taxon is found in South China.It is interesting to note that only in the DFZ 7 are truly cosmopolitan taxa present, which are found in all 8 palaeobiogeographical units (2-9).Especially, the first biozones of the Mississippian (MFZ 1-2)show a very high degree of widespread taxa during a time of overall low total diversity.This interval corresponds to the time when foraminiferal associations are dominated by simple and small forms.From the upper Hastarian onward (MFZ 3-4), the entire range of palaeobiogeographical spread is found, from endemic to cosmopolitan forms.The only exception is MFZ 5, where no taxa colonized all potential palaeobiogeographical units.Genera are more wide spread than species in the entire study interval, which is the expression of the larger geographical stability of the higher taxonomic rank.
Looking at the mean values for the geographical distribution of genera, this value f l uctuates between 5 and 6,thus about the half of all possible units, for the Ivorian to the Pendleian biozones.In the Strunian and Hastarian,this value is higher.Since species are less widespread than genera, the values on the species level are lower.The resulting curve shows more variation than the genus curve,and especially the amplitudes are more pronounced.The Strunian and Hastarian show pronounced changes, but these changes are often the result from the integration of poorly diversif i ed or known biozones (e.g., DFZ 6, MFZ 2).The values for the better studied and known biozones are relatively similar.During the Ivorian to Pendleian,the amplitude of changes is lower than in the two older substages.The highs and lows in the curve correspond to the peaks in the curve for genera, but again they are more pronounced.The highest mean values for the geographical spread in this interval are those of biozones MFZ 13 and 14 (Figure 7A, 7B), again periods where simple forms dominated.
The endemicity curves show the development of endemic taxa from the upper Hastarian onwards, prior to this not a single endemic taxon is known in DFZ 7.On the genus level, the endemicity rises with minor f l uctuations until the lower Warnantian, before it starts to drop (Figure 7A).Endemicity values for species are higher and the fl uctuations are more important than for the genera.The Ivorian to Livian interval forms a dissected plateau with a mean of 27% (14.7%-34.1%)before endemicity drops considerably in the Warnantion (Figure 7B).The endemicity on the species level correlates relatively well with the intensively studied intervals by Hanceet al.(2011)and partly with the total biodiversity; on the genus level this correlation is less pronounced.
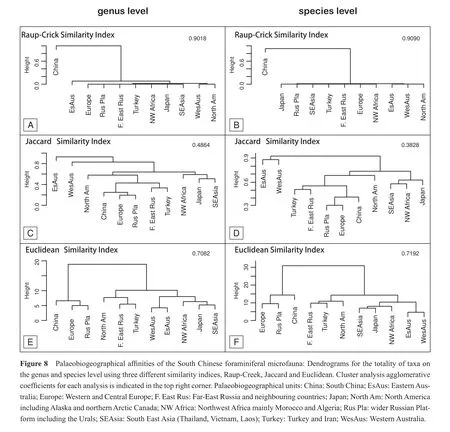
Clusters have been calculated on the genus and species level for the entire study interval (Figure 8)and individual substages (not shown herein)to obtain an overview on possible palaeogeographical affinities of the South Chinese fauna.To evaluate the inf l uence of the calculation method, the clusters have been calculated with three commonly used similarity indices (Raup-Crick, Jaccard and Euclidean similarity indices).It is important to note that these analyses show palaeogeographical affinities of the south Chinese fauna and do not show the palaeobiogeographical relations between the eleven regions, because taxa known only outside South China were not included into the analysis.
Independently from the taxonomic level, the two dendrograms for the same index are similar (Figures 8-10).On the dendrograms calculated with the Raup-Crick index, South China is situated at the opposite corner of the dendrogram isolated from all other palaeobiogeographical units, which are all connected to each other at the base of the dendrogram (Figure 8A, 8B).It is visually diff i cult to differentiate distinctive clusters, with an exception for Eastern Australian and for Western Australian associated to the North American genera.The shape of the dendrograms results from one particularity when applying the Raup-Crick index to our data matrix.In contrast to the usual palaeobiogeographical analyses, there is one unit (South China)with 100% taxa present.This unit dominates all calculations using the Raup-Crick algorithm.
The dendrograms calculated with Jaccard and Euclidean indices show better resolved clusters, which resemble an uneven bar-bell (Figure 8C-8F).Common to both is a stable cluster of South China, Europe and the Russian Platform.With the Euclidean index, this cluster is well separated from a second large digitated cluster comprising all other units (Figure 8E, 8F).In the dendrogram using the Jaccard index the cluster containing China is part of a larger digitated cluster.Stable on both taxonomic levels is the order of cluster branching.The Far-East Russia and Turkey cluster connects to North America on one side of the dendrogram and the cluster SE Asia, Japan and NW Africa constitues the second side of the dendrogram (Figure 8C, 8D).Depending on the similarity matric used, the Australian units from a cluster clearly separated from all other units or as a cluster connected to the second side of the dendrogram.
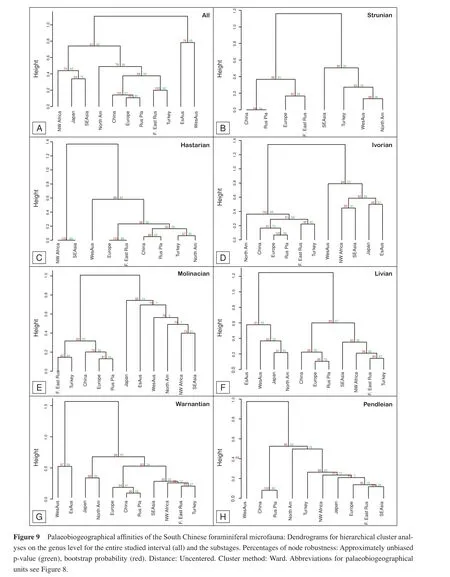
Hierarchical clusters and non-metric multivariate ordination analysis (NMDS)were calculated to foster and question the results of the previously presented cluster analyses (Figures 9-11).The two hierarchical clusters on the genus and species level for each studied time interval(Figures 9, 10)show the same pattern, which is overall comparable to the Jaccard clusters (Figure 8).As in the Jaccard and Euclidean clusters, South China is closest to a bundle formed by Western and Central Europe and the wider Russian Platform.To this cluster is attached the cluster of Far-East Russia and Turkey/Iran, thus the two palaeobiogeographical units situated between South China and the Russian Platform.This clearly indicates the good faunal interconnection in the northern Palaeotethyan Realm.The large distance in the cluster analysis between South China and its southward neighbouring palaeogeographical units (SE Asia, Western Australia)indicates less important faunal interchange between these regions.However, on the species and genus level, the Australian faunas are the regions with the least interchange with China.This was expected for Eastern Australia, since its faunal exchange with the Palaeotethyan Realm is hampered by ocean circulation patterns (Webb, 2000; Aretzet al., 2013).The poor exchange with Western Australia may be partly rooted in oceanic current patterns in the Palaeotethyan Realm (Aretzet al., 2013), but the analysis is certainly affected by the integration of data for Western Australia.The periods of high diversity in South China are often poorly known or less diverse in Western Australia (e.g., Readet al., 1973).On the other hand, the rich late Visean assemblages from the Bonaparte Basin (Mamet and Belford, 1968)falls in a period when data from South China are less abundant, and thus the palaeobiogeographical affinities of the Bonaparte assemblages are not expressed by the comparison with the Chinese data.The relative closeness of North America to South China is documented on the species and genus levels; in fact North America is directly attached to the northern Palaeotethyan cluster.
To detect possible changing palaeogeographical affinities of southern China through time, hierarchical clusters(Figures 9, 10)and the above-mentioned Raup-Crick, Jaccard and Euclidean indices clusters (not shown here)have been calculated for every substage.There are two principal patterns for the position of South China in those dendrograms.The first pattern is that found in the general dendrogram in which South China is attached to a cluster of Europe and Russian Platform.This pattern is found in the Ivorian, Molinacian and Livian substages (Figures 9D-9F; 10D-10F).Before and after, China forms a cluster with the Russian Platform.The position of this cluster and thus the closeness to the other regions varies between the different substages, but also on the taxonomic level.
This is an expression of the important differences between the dendrograms of the different substages and taxonomical levels.On the genus level the units representing the northern Palaeotethys are mostly part of a larger cluster from the Strunian to the Livian (Figures 9B-9F;10B-10F).During several substages, North America is part of this larger cluster.The Warnantian and Pendleian dendrograms (Figures 9G-9H; 10G-10H)show the gradual disappearance of this Palaeotethyan constant.Especially, the Pendleian dendrogram shows a clustering of palaeobiogeographical units (Figures 9H, 10H), which do not correspond to palaeogeographical proximities of these units.The Palaeotethyan cluster can also be identif i ed on the species level.It can be a little more pronounced as in the Strunian and Livian or more diluted as in the Ivorian.

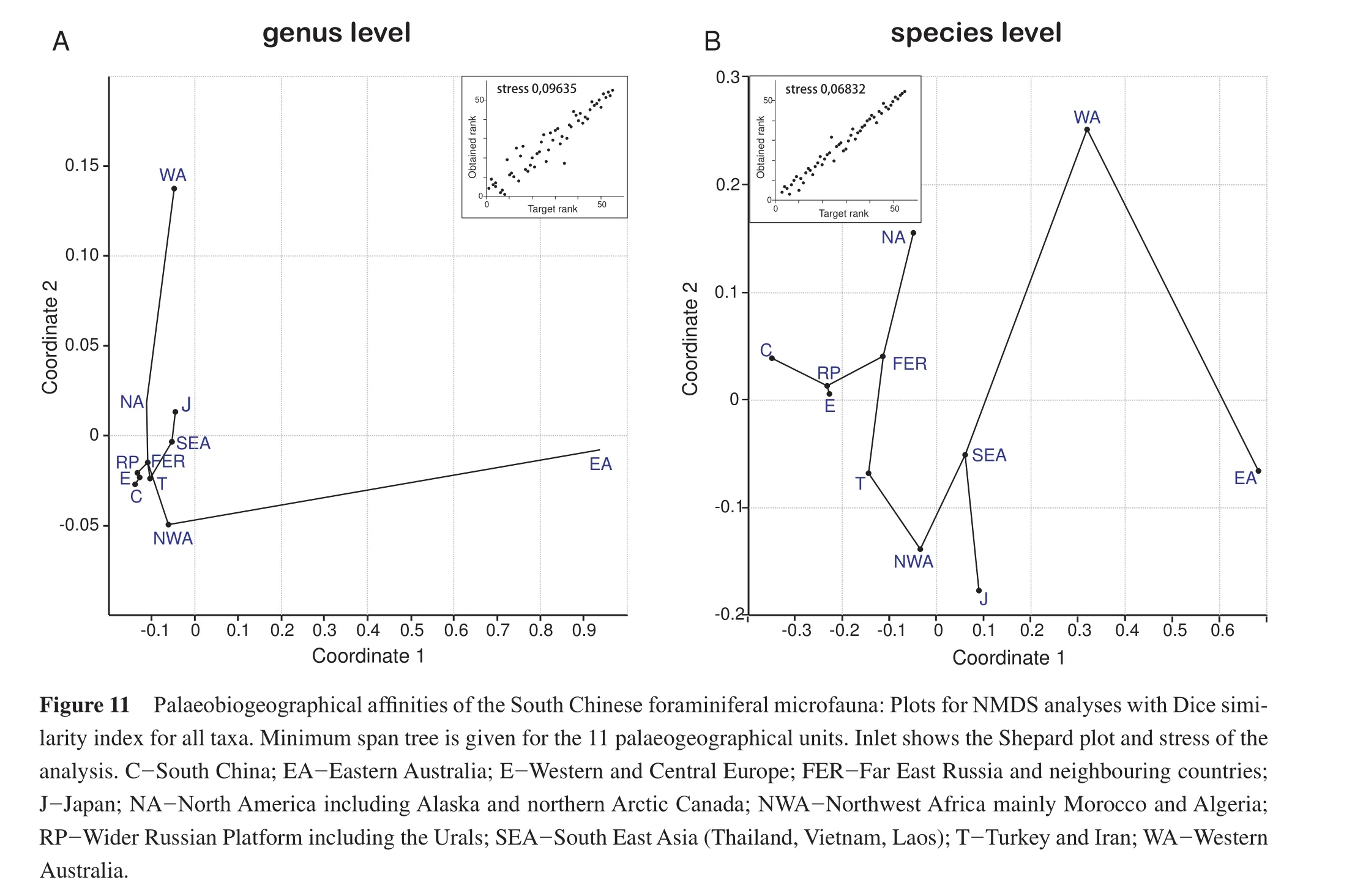
The NMDS analyses (Figure 11)show a similar pattern.Overall, the analysis on the genus level results in a denser cluster (Figure 11A), with Eastern Australia being the most isolated entity on both taxonomic levels.The cluster of South China, Europe and Russian Platform is well identif i ed and this cluster is nearest to Far-East Russia and Turkey.Differences exist between the genus and species for the position and attachment of the other units.North America is situated not too far from the core cluster, but while the minimum span tree indicates on the genus level a connection to Western Australia (Figure 11A), it forms a dead end at the species level (Figure 11B).The position of the other units changes between species and genus level,but the two Australian units are always the most distanced and isolated units, conf i rming the cluster analyses.The minimum span tree for the genus level shows a peculiar relationship of North-West Africa, which is attached to Eastern Australia (Figure 11A)in contrast to the cluster analyses and the NMDS on the species level, where it clusters with SE Asia and Japan (Figure 11B).
5.3 Discussion
The highest mean values for the geographical spread of foraminifers have been observed in the lower Hastarian and lower Warnantian, which are intervals characterised by low total diversity and dominance of small, simple (often unilocular or bilocular)forms.These forms are often called disaster forms or ecological opportunists and their wide geographically distribution or even cosmopolitan character seems to be evident.In the South Chinese dataset, simple forms are among those taxa known from at least 9 palaeobiogeographical units, but there are also taxa with a more reduced geographical extent, indicating that they are not necessarily cosmopolitan.The dominance of these simple forms in the lowermost Hastarian can be linked to the aftermath of a global extinction event (‘Lilliput-Effect’), but their dominance is not necessarily correlated to a possible extinction event, as evidenced in the case of the lower Warnantian.Indeed, a global bioevent is unknown at the Livian/Warnantian boundary, and the dominance of simple forms in South China is the effect of a deterioration in the regional palaeoenvironmental conditions (see above).Moreover, the wide geographical distribution of simple forms can be also explained by their simple test,which do not offer many morphological characters to differentiate taxa (e.g., Krainer and Vachard, 2011).One may ask if several genotypes are lumped together, but it also helps to prevent taxonomic over-splitting.
Increasing endemicity in southern China goes hand in hand with the development of morphologically complex forms (e.g., appearance and evolution of Dainellidae).This does not mean that large, complex forms are mainly endemic, because the database contains numerous examples of large, complex, cosmopolitan taxa.However, some of these complex forms seem to be adapted to specif i c environments (see discussions in Vachardet al., 2010), thus increasing the potential for oligotrophy and possible endemism.
In South China, possible endemicity shows a peak in the Livian (MFZ 12)on the species level (Figure 7B).Taxonomic over-splitting can be ruled out as an explanation.The fauna is taxonomically diversif i ed, but does not show extreme values.Endemic forms are widely distributed among species and genera.The Livian endemism points to the establishment of palaeobiogeographical barriers between South China and neighbouring regions, probably connected to a lower global sea-level.It is interesting to note that in several platform systems and regions, the Livian is diff i cult to identify or is unknown (e.g., North Africa)or characterised by major advances of terrigenous clastic facies (e.g., North-West Carboniferous Basin, Ireland)(Aretzet al., 2010), Northumberland Trough (Waterset al., 2011).It cannot be ruled out that the poorer knowledge and documentation of the Livian in other regions could contribute to accentuated endemicity in South China where the interval is well-studied.This points to the more general problem of a lack of quantitative data from other well-studied regions.The Belgian Namur-Dinant Basin is a standard for the study of Carboniferous biostratigraphy and calcareous foraminifers and a wealth of data is accessible (see for references Potyet al., 2006 and in press), but no synthesis has been published so far.
Poty (2007)showed the global break-down of biogeographical barriers and intensive faunal exchange in the uppermost Ivorian.The most important Mississippian sealevel drop (Hanceet al., 2001; Poty, 2007)follows this so-called ‘Avins Event’ resulting in considerably reduced and geographically isolated platform areas.This development can be seen in South China in the endemicity peak of MFZ 9, and its decrease during MFZ 10, probably due to sea-level rise causing the break-down of barriers.Hence,the global sea-level curve highly inf l uences the faunal interchange, and endemicity and biodiversity are thus consequences of regional origination patterns.Using the 3rd-order sequence model of Hanceet al.(2001), the lowstand and transgressive system tracts should correspond to times of elevated endemicity and regional origination rates, whereas the highstand system tracts should correspond to wide geographical spread and low endemicity.This causality is diff i cult to see in the Chinese data.It is most evident for the interval MFZ 8-MFZ 10, which can be roughly correlated to the HST (MFZ 8), LST and TST (MFZ 9)and HST (MFZ 10)of sequences 4 and 5 of Hanceet al.(2001).A more detailed temporal resolution of the data would be needed to test in detail the relations of 3rd-order sequences, biodiversity and palaeobiogeographical patterns.
The analyses of palaeobiogeographical affinities clearly show that the fauna of the northern Palaeotethyan Realm is well interconnected.This result was expected, based on the qualitative palaeobiogeographical models (Kalvoda,1990; 2002).However, the better connection between South China and North America than between South China and palaeobiogeographical units of the southern Palaeotethyan Realm (e.g., NW Africa)raises some questions.They partly originate from the analyses themselves,because only species present in China are analysed.Thus,the relative poorness of late Visean foraminifers in South China excludes the full integration of the diversif i ed late Visean assemblages of Western Australia and North Africa (see references above).The Palaeotethyan affinity of Alaska and northern Canada included in North America is certainly one crucial element for the position of North America in our palaeobiogeographical analyses (Figures 8-11).These two regions are in other analyses included into the Palaeotethyan Realm (e.g., Groves and Wang,2009).On the other hand, the foraminiferal assemblages of North America are more diverse and better studied than regions like NW Africa or Western Australia, supposedly better connected to South China.The separate analyses for the Warnantian substage, where the datasheet contains more African than North American taxa, does not change the general pattern.Thus, the conclusion that the closeness between South China and North America could (partly)be the expression of work effort and not of palaeobiogeographical connections is not supported.However, it is conspicuous that the well-studied palaeobiogeographical units form a stable cluster in the analyses (Figures 10, 11), and that those regions with less well-known or more discontinuous records are more remote.
The clustering of the palaeobiogeographical affinities in the Serpukhovian has been relatively different than those in earlier substages (Figures 8-11).This reorganisation cannot be connected to the closing of the Rheic Ocean,which is often the origin for changes in palaeobiogeographical models in late Visean-Serpukhovian times (e.g.,Kornet al., 2012)and the establishment of well-separated North American and Palaeotethyan Realms.The pattern is random and it does not follow the positioning of palaeogeographical units or closeness of palaeobiogeographical affinities/units.It must be suspected that, as for the biodiversity, the analyses suffer in this time slice from poor data.The same problem may exist for the Warnantian.There are changes in the clustering compared to the older Visean substages, which could be seen as a prelude to the situation in the Serpukhovian.
Dispersal of foraminifers in the northern Palaeotethyan Realm was eff i cient, as seen in the closeness of the f i ve palaeobiogeographical units in the clusters, hierarchical clusters and NMDS, and the overall high values of geographical spread.The present study cannot answer the question where the biogeographical centre of origination is situated.South China would be an interesting candidate along with other regions like the Urals, Namur-Dinant Basin or Siberia.Westward oceanic currents have been proposed (e.g., Webb, 2000)and also modelled (Aretzet al.,2013)for this part of the Palaeotethyan Ocean.However,our study lacks the temporal resolution to show important migration f l ux of calcareous foraminifers from South China to other regions.The comparison of the Strunian foraminifers showed the higher abundance and earlier appearances of quasiendothyrids in the Urals than in South China (see 4.2).It seems to be reasonable to propose a migration of these forms into South China.The absence of any signif i cant endemicity may also be used as an argument that South China was more likely to import taxa,because it would be very unusual that all indigenous taxa would be exported.Moreover, the eastward migration has also been found among Mississippian rugose corals (Potyet al., 2011; Aretzet al., 2013), although the dispersion of corals is often much slower than for calcareous foraminifers.This does not minimize the possibilities of westward migration or dispersal starting in South China.It further questions a single biogeographical centre, but in any case the high degree of geographical spread and abundance of even cosmopolitan taxa in calcareous foraminifers occurring in the same biozones advocates for very eff i cient and fast distribution.It may be questionable if the temporal resolution in deep time records enables the identif i cation of these fast processes.
6 Conclusions
The analysis of the dataset compiled for South China based on recent taxonomic and biostratigraphic work in several key sections in Guangxi, Guizhou and Hunan is one of the first attempts to quantify regional diversity and palaeobiogeographical affinities of latest Famennian to Serpukhovian calcareous foraminifers.
The diversity curve for the Strunian to Pendleian of southern China has a bell-shaped form, with a double peak in the Ivorian.Its shape is partly inf l uenced by regional palaeoenvironmental conditions, which hamper the development of rich foraminiferal associations (MFZ 2, 13, 14).Diversity in the Strunian and basal Hastarian is rather low.The extinction event of the Hangenberg crises in the latest Devonian is documented in a biodiversity drop at the DCB and by the composition of the basal Mississippian biozone.The latter is characterised by abundant small unilocular and bilocular foraminifers (mainly,Bisphaeraspp.andEarlandiaspp.)and the absence of complex, large forms,which became extinct during the crises.Hence, MFZ 1 exhibits the classical characteristics of a ‘Lilliput-Effect’ in the aftermath of a global mass extinction.In South China the onset of the diversif i cation in the Hastarian is delayed by one biozone compared to other regions due to unsuitable facies for foraminifers (MFZ 2).The highest diversities are found around the Tournaisian-Visean boundary and can be directly related to the very detailed work done for the GSSP at Pengchong.The impact of this work can be directly seen in an exceptionally high value.In the last Tournaisian biozone, 87% of all known species are found in southern China (mean for other biozones with medium to high biodiversity is 30%-40%).The opposite is seen in the late Visean-Serpukhovian.In South China, this time interval is not so well exposed and less studied than the older substages.In consequence, South China is completely decoupled from the global trend.In all other subtropical and tropical palaeobiogeographical provinces the late Visean-Serpukhovian shows the highest diversity of the entire Mississippian; in South China it is rather low.
Hence, there is a clear link between work effort and biodiversity data, at least on the regional scale of South China.It can be expected that this is also true for other regions, and thus our current knowledge of quantitative biodiversity data on regional and global scales may be largely biased.The idea that the Palaeotethyan Realm was the centre of origination (Mamet, 1977)to explain differences between diversity curves for the North American and Palaeotethyan Realm (Groves and Wang, 2009)is not conf i rmed here.The non-normalisation of the potential habitat surface in these two realms simply favours a higher diversity in the larger realm, which is the case for the Palaeotethys.Thus, the size relation of the realms has to be corrected, more data from regions in the central and southern Palaeotethys should be included (regions which have largely been excluded in the dataset of Groves and Wang, 2009), and finally a tripolar or quadrupolar palaeobiogeographical model should be applied.
Palaeobiogeographical affinities of the south Chinese calcareous foraminifers are stable through time, and only minor differences exist between the genus and species level.They are closest to two regions, the wider Russian Platform including the Urals and Western and Central Europe.These three regions form with very few exceptions a stable cluster in most analyses (cluster, hierarchical cluster and NMDS).The two palaeobiogeographical units situated between the Russian Platform and South China are often closely connected to the basic tripolar cluster, and thus characterise important faunal exchange in the northern Palaeotethyan Realm.For the moment, it is impossible to determine the amount of migration into and out of South China.Although oceanic currents may favour the migration from South China towards the west, there are increasing evidences that eastward migration was possible as well.Strunian quasiendothyrids are one possible example of eastward migration; from the Urals (West)into South China (East).
An unresolved problem is the closer palaeobiogeographical af finity of South China to North America compared to neighbouring palaeo(bio)geographical units in the southern Palaeotethys (SE Asia, Western Australia).The contour for North America chosen in this study may infl uence the analyses because the northern regions of the continent have been included although they show af finities to the Palaeotethyan Realm.However, even in Warnantian times, when more species are found in the southern Palaeotethys than in North America, the latter is still closer clustered to South China.This is one example when sample size may not in fl uence the analyses, but overall the well-studied regions cluster closer than the regions with poorer data.
Previous studies have already demonstrated that cosmopolitan calcareous foraminifers are common in the Strunian to Pendleian.This is con fi rmed by the large geographical spread found in all studied biozones.It is curious to see that endemicity is almost absent in the Strunian and basal Hastarian.It first developed with the re-appearance of large, plurilocular forms in the upper Hastarian.Endemicity remains on a relatively stable level in the Ivorian,Molinacian and Livian before it drops to lower values in the Warnantian.Thus, endemicity roughly parallels biodiversity.
The Tournaisian-Visean boundary interval is the only example in South China, where causality between changes in endemicity, origination rates and total biodiversity and 3rd-order sea-level fl uctuations could possibly be documented.Good faunal exchange should be related to high relative sea-level, whereas important local and regional origination should occur during times of geographical isolation.However, the temporal resolution of the South Chinese data is too large to work on this problem.
Acknowledgements
We thank Ian Somerville (Dublin)for his constructive review and Yuan Wang, Min Liu and Xiu-Fang Hu for editorial support.The authors would like to acknowledge their Chinese and Belgian colleagues who generated the data used in this study.
Alve, E., 1999.Colonization of new habitats by benthic foraminifera: A review.Earth-Science Reviews, 46: 167-185.
Aretz, M., Dera, G., Lefebvre, V., Donnadieu, Y., Goddéris, Y., Macouin, M., Nardin, E., 2013.The spatial and temporal distribution of Mississippian rugose corals: Contribution of modelled oceanic currents and temperature data to this problem.In: Nardin, E.,Aretz, M., (eds).Pre-Cenozoic climates international workshop.When data and modelling meet.Strata, Série 1, 14: 8-9.
Aretz, M., Herbig, H.-G., Somerville, I.D., Cózar, P., 2010.Rugose coral biostromes in the late Visean (Mississippian)of NW Ireland: Bioevents on an extensive carbonate platform.Palaeogeography, Palaeoclimatology, Palaeoecology, 292: 488-506.
Aretz, M., Poty, E., Devuyst, F.-X., Hance, L., Hou, H., 2012.Late Tournaisian Waulsortian-like carbonate mud banks from South China (Longdianshan Hill, central Guangxi): Preliminary investigations.Geological Journal, 47: 450-461.
Blakey, R.C., 2007.Carboniferous-Permian paleogeography of the assembly of Pangea.In: Wong, Th.E., (ed).Proceedings of the XVth International Congress on Carboniferous and Permian Stratigraphy.Utrecht, the Netherlands, 10-16 August 2003.Royal Netherlands Academy of Arts and Sciences, 443-465.
Buggisch, W., Joachimski, M.M., Sevastopulo, G., Morrow, J.R.,2008.Mississippian δ13C carb and conodont apatite δ18O records— Their relation to the Late Palaeozoic Glaciation.Palaeogeography, Palaeoclimatology, Palaeoecology, 268: 273-292.
Cascales-Miñana, B., Cleal, C.J., Diez, J.B., 2013.What is the best way to measure extinction? A ref l ection from the palaeobotanical record.Earth-Science Reviews, 124: 126-147.
Clarke, K.R., 1993.Non-parametric multivariate analyses of changes in community structure.Australian Journal of Ecology, 18:117-143.
Coen, M., Hance, L., Hou, H., 1996.Papers on the Devonian-Carboniferous transition beds of central Hunan, South China.Mémoires de l’Institut Géologique de l’Université de Louvain, 36:229.
Conil, R., De Putter, T., Hou, H., Wei, J.Y., Wu, X.H., 1988.Contribution a l’étude des Foraminifères du Strunien et du Dinantien de la Chine sud-orientale.Bulletin de la Société Belge de Géologie,97: 47-61.
Conil, R., Lys, M., 1964.Matériaux pour l’étude micropaléontologique du Dinantien de la Belgique et de la France (Avesnois):Partie 1, algues et foraminifères, et partie 2, foraminifères.Mémoires de l’Institut géologique de l’Université de Louvain, 23:1-296.
Cooper, A., 2004.Measures of Biodiversity.In: Webby, B.D., Paris,F., Droser, M.L., Percival, I.G., (eds).The Great Ordovician Biodiversif i cation Event.New York: Columbia University Press,52-57.
Cózar, P., 2003.Foraminiferal fauna and zonation from the Lower Carboniferous of the Guadiato area SW Spain: Comparison with other European and North African foraminiferal zonation schemes and their paleobiogeographical implications.In: Ahr, W.M., Harris, P.M., Morgan, W.A., Somerville, I.D., (eds).Permo-Carboniferous Carbonate Platforms and Reefs.SEPM Special Publication, 78 and AAPG Memoir, 83: 155-170.
Cózar, P., Medina-Varea, P., Somerville, I.D., Vachard, D., Rodríguez, S., Said, I., 2013.Foraminifers and conodonts from the late Visean to early Bashkirian succession in the Saharan Tindouf Basin (southern Morocco): Biostratigraphic refinements and implications for correlations in the western Palaeotethys.Geological Journal, on line, doi: 10.1002/gj.2519.
Cózar, P., Said, I., Somerville, I.D., Vachard, D., Medina-Varea, P.,Rodríguez, S., Berkhli, M., 2011.Potential foraminiferal markers for the Visean-Serpukhovian and Serpukhovian-Bashkirian boundaries — A case-study from Central Morocco.Journal of Paleontology, 85: 1105-1127.
Cózar, P., Somerville, I.D., 2004.New algal/foraminiferal evidence for the recognition of the Asbian/Brigantian boundary in northern England.Proceedings of the Yorkshire Geological Society, 55:43-65.
Devuyst, F.-X., 2006.The Tournaisian-Visean boundary in Eurasia.Definition, biostratigraphy, sedimentology and early evolution of the genus Eoparastaffella (foraminifer).Thèse doctorale, Université catholique de Louvain, 430.
Devuyst, F.-X., Hance, L., Hou, H., Wu, X., Tian, S., Coen, M.,Sevastopulo, G., 2003.A proposed global stratotype section and point for the base of the Visean Stage (Carboniferous): The Pengchong section, Guangxi, South China.Episodes, 26: 105-115.
Devuyst, F.-X., Kalvoda, J., 2007.Early evolution of the genusEoparastaffella(Foraminifera)in Eurasia: The ‘interiectagroup’ and related forms, late Tournaisian to early Visean (Mississippian).Journal of Foraminiferal Research, 37: 69-89.
Fielding, C.R., Frank, T.D., Birgenheier, L.P., Rygel, M.C., Jones,A.T., Roberts, J., 2008.Stratigraphic imprint of the Late Palaeozoic Ice Age in eastern Australia: A record of alternating glacial and non-glacial climate regime.Journal Geological Society London, 165: 129-140.
Foote, M., 2000a.Origination and extinction components of taxonomic diversity: General problems.In: Erwin, D.H., Wing, S.L., (eds).Deep Time: Paleobiology’s Perspective.Paleobiology,26 (Suppl.): 74-102.
Foote, M., 2000b.Origination and extinction components of taxonomic diversity: Paleozoic and post-Paleozoic dynamics.Paleobiology, 26 (Suppl.): 578-605.
Gallagher, S.J., 1998.Controls on the distribution of calcareous Foraminifera in the Lower Carboniferous of Ireland.Marine Micropalaeontology, 34: 187-211.
Gradstein, F.M., Ogg, J.G., Smith, A.G., Bleeker, W., Lourens,L., 2004.A New Geologic Time Scale, with special reference to Precambrian and Neogene.Episodes, 27: 83-100.
Gradstein, F.M, Ogg, J.G., Schmitz, M.D., Ogg, G., 2012.The Geologic Time Scale 2012.Oxford: Elsevier, 1176.
Groves, J.R., Lee, A., 2008.Accelerated rates of foraminiferal origination and extinction during the Late Paleozoic ice age.Journal of Foraminiferal Research, 38: 74-84.
Groves, J., Wang, Y., 2009.Foraminiferal diversif i cation during the Late Paleozoic ice age.Paleobiology, 35: 367-392.
Groves, J., Wang, Y., Qi, Y., Richards, B.C., Ueno, K., Wang, X.D.,2012.Foraminiferal biostratigraphy of the Visean-Serpukhovian(Mississippian)boundary interval at slope and platform sections in southern Guizhou (South China).Journal of Paleontology, 86:753-774.
Hance, L., Hou, H., Vachard, D., 2011.Upper Famennian to Visean Foraminifers and some carbonate Microproblematica from South China.Beijing: Geological Publishing House, 330.
Hance, L., Muchez, P., Hou, H., Wu, X., 1997.Biostratigraphy, sedimentology and sequence stratigraphy of the Tournaisian-Visean transitional strata in South China (Guangxi).Geological Journal,32: 337-357.
Hance, L., Poty, E., 2006.Hastarian.Geologica Belgica, 9: 111-116.
Hance, L., Poty, E., Devuyst, F.-X., 2001.Stratigraphie séquentielle du Dinantien type (Belgique)et corrélation avec le Nord de la France (Boulonnais, Avesnois).Bulletin de la Société Géologique de France, 172, 4: 411-426.
Hou, H., Wu, X., Yin, B., 2011.Correlation of the Tournaisian-Visean Boundary Beds.Acta Geologica Sinica - English Edition,85: 354-365.
Isaacson, P.E., Díaz-Martínez, E., Grader, G.W., Kalvoda, J., Babek,O., Devuyst, F.-X., 2008.Late Devonian-earliest Mississippian glaciation in Gondwanaland and its biogeographic consequences.Palaeogeography, Palaeoclimatology, Palaeoecology, 268: 126-142.
Isbell, J.L., Miller, M.F., Wolfe, K.L., Lenaker, P.A., 2003.Timing of Late Paleozoic glaciation in Gondwana: Was glaciation responsible for the development of northern hemisphere cyclothems? In: Cham, M.A., Archer, A.W., (eds).Extreme Depositional Environments: Mega End Members in Geological Time.Geological Society of America, Special Paper, 370: 5-24.
Jin, X., Devuyst, F.-X., Hance, L., Poty, E., Aretz, M., Yin, B., Hou,H., 2007.Stratigraphy and lithofacies of the Tournaisian and Visean in the Guilin-Liuzhou area, Guangxi, South China.16th International Congress on the Carboniferous and Permian, Nanjing,Guide Book for Field Excursion C2, 41.
Kalvoda, J., 1990.Late Devonian-Lower Carboniferous paleobio-geography of benthic foraminifera and climatic oscillations.In:Kauffman, E.G., Walliser, O.H., (eds).Extinction events in Earth history.Berlin: Springer Verlag, Lecture Notes in Earth Sciences, 30: 183-188.
Kalvoda, J., 2001.Upper Devonian-Lower Carboniferous foraminiferal paleobiogeography and Perigondwana terranes at the Baltica-Gondwana interface.Geologica Carpathica, 52: 205-215.
Kalvoda, J., 2002.Late Devonian-Early Carboniferous foraminiferal fauna: Zonations, evolutionary events, paleobiogeography and tectonic implications.Folia, Geologia, 39: 1-213.
Kalvoda, J., Bábek, O., Aretz, M., Cossey, P., Devuyst, F.-X., Hargreaves, S., Nudds, J., 2012.High resolution biostratigraphy of the T-V boundary interval in the North Staffordshire Basin and correlation with the South Wales-Mendip shelf.Bulletin of Geosciences, 87: 497-541.
Kalvoda, J., Devuyst, F.-X., Bábek, O., Dvořák, L., Rak, Š., Rez, J.,2010.High-resolution biostratigraphy of the Tournaisian-Visean(Carboniferous)boundary interval, Mokrá quarry, Czech Republic.Geobios, 43: 317-331.
Kalvoda, J., Kumpan, T., Bábek, O.Upper Famennian and Lower Tournaisian sections of the Moravian Karst (Moravo-Silesian Zone, Czech Republic): A proposed key area for correlation of the conodont and foraminiferal zonations.Geological Journal, in press, doi: 10.1002/gj.2523.
Kaminski, M.A., Setoyama, E., Cetean, C.G., 2010.The Phanerozoic Diversity of Agglutinated Foraminifera: Origination and Extinction Rates.Acta Palaeontologica Polonica, 55(3): 529-539.
Korn, D., Titus, A.L., Ebbighausen, V., Mapes, R.H., Sudare, M.N.,2012.Early Carboniferous (Mississippian)ammonoid biogeography.Geobios, 45: 67-77.
Krainer K., Vachard D., 2011.The Lower Triassic Werfen Formation of The Karawanken Mountains (southern Austria)and its disaster survivor microfossils, with emphasis on Postcladella n.gen.(Foraminifera, Miliolata, Cornuspirida).Revue de Micropaléontologie, 54: 59-85.
Kulagina, E.I., 2013.Taxonomic diversity of foraminifers of the Devonian-Carboniferous boundary interval in the South Urals.Bulletin of Geosciences, 88: 265-282.
Leven, E.Y., 1993.Main events in the Permian history of the Tethys and Fusulinids.Moscow: Stratigraphy and Geological Correlation, I: 51-65.
Lipina, O.A., 1973.Zonal stratigraphy and paleogeography based on Tournaisian foraminifers (Zonalnaya st ratigraf i ya i paleogeograf i ya turne po foraminiferam).Voprosy mikropaleontologii,16: 3-34 (In Russian).
Loeblich, A.R.Jr., Tappan, H., 1987.Foraminiferal Genera and their classification.New York: van Nostrand Reinhold Company, 2:970 (Imprinted 1988).
Mamet, B.L., Belford, D., 1968.Carboniferous Foraminifera, Bonaparte Gulf Basin, North-Western Australia.Micropaleontology,14: 339-347.
Mamet, B.L., 1977.Foraminiferal zonation of the Lower Carboniferous: Methods and stratigraphic implications.In: Kauffman, E.G., Hazel, J.E., (eds).Concepts and Methods of Biostratigraphy.New York: Dowden, Hutchinson and Ross, 445-462.
Mamet, B.L., Skipp, B., 1970.Lower Carboniferous calcareous Foraminifera; preliminary zonation and stratigraphic implications for the Mississippian of North America.International Congress of Carboniferous Stratigraphy and Geology, 6th, Sheffield, 1967,Comptes Rendus, 3: 1129-1146.
Marshall, J.E.A., Lakin, J., Finney, S.M., 2013.Terrestrial climate and ecosystem change from the Devonian-Carboniferous boundary to the earliest Visean interval in East Greenland.Documents de l’Institut Scientif i que, Rabat, 26: 81-82.
McCune, B., Grace, J.B., 2002.Chapter 11 Hierarchical clustering.In: McCune, B., Grace, J.B., (eds).Analysis of Ecological Communities.Glenedon, 86-96.
Nardin, E., Lefebvre, B., 2010.Unravelling extrinsic and intrinsic factors of the early Palaeozoic diversif i cation of blastozoan echinoderms.Palaeogeography, Palaeoclimatology, Palaeoecology,294: 142-160.
Okuyucu, C., Vachard, D., Göncüoglu, M.C., 2013.Refinements in biostratigraphy of the foraminiferal zone MFZ11 (late early Visean, Mississippian)in the Cebeciköy Limestone (Ýstanbul Terrane, NW Turkey)and palaeogeographic implications.Bulletin of Geosciences, 88: 621-645.
Poty, E., 2007.The Avins event: A remarkable worldwide spread of corals at the end of the Tournaisian (Lower Carboniferous).In:Hubmann, B., Piller, W.E., (eds).Fossil corals and sponges; Proceedings of the 9th International Symposium on Fossil Cnidaria and Porifera, Graz 2003.Schriftenreihe der Erdwissenschaftlichen Kommissionen, Oesterreichische Akademie der Wissenschaften, 17: 231-249.
Poty, E., Aretz, M., Hance, L.Belgian substages as a basis for an international chronostratigraphic division of the Tournaisian and Visean.Geological Magazine, in press, doi: 10.1017/S0016756813000587.
Poty, E., Aretz, M., Xu, S., 2011.A comparison of Mississippian colonial rugose corals from Western Europe and South China.In: Aretz, M., Delculée, S., Denayer, J., Poty, E., (eds).XIth International Symposium on Fossil Cnidaria and Porifera, Liège,Belgium.Kölner Forum für Geologie und Paläontologie, 19:135-136.
Poty, E., Devuyst, F.-X., Hance, L., 2006.Upper Devonian and Mississippian foraminiferal and rugose coral zonation of Belgium and Northern France: A tool for Eurasian correlations.Geological Magazine, 143: 829-857.
Poty, E., Mottequin, B., Denayer, J., 2013.Orbitally forced sequences in the Lower Carboniferous and the onset of Carboniferous glaciations at the Tournaisian Visean boundary.In: Nardin, E.,Aretz, M., (eds).Pre-Cenozoic climates international workshop.When data and modelling meet.Strata, Série 1, 14: 66.
R.Development Core Team.2009.A Langage and Environment for Statistical Computing.R Foundation for Statistical Computing.Vienna (Austria).
Raup, D.M., Crick, R., 1979.Measurement of faunal similarity in paleontology.Journal of Paleontology, 53: 1213-1227.
Rauser-Chernousova, D.M., Bensh, F.R., Vdovenko, M.V., Gibshman, N.B., Leven, E.Y., Lipina, O.A., Reitlinger, E.A.,Solov’eva, M.N., Chediya, I.O., 1996.Reference book on the systematics of Paleozoic Foraminifera (Endothyroida, Fusulinoida).Rossiiskaya Akademiya Nauk, Geologicheskii Institut,Moskva ‘‘Nauka”, 204 (In Russian).
Read, J.F., Alcock, P.J., Hoseman, P., 1973.Harris Sandstone-Yindagindy Formation relationships and possible position of Permo-Carboniferous boundary, Carnarvon Basin, Western Australia.Journal of the Royal Society of Western Australia, 56: 80-85.
Ross, C.A., 1973.Carboniferous Foraminiferida.In: Hallam, A.,(ed).Atlas of Palaeobiogeography.Amsterdam: Elsevier Scientif i c, 127-132.
Sepkoski, J.J.Jr., 2002.A compendium of fossil marine animal genera.Bulletins of American Paleontology, 363: 1-560.
Shen, J.W., Qing, H., 2010.Mississippian (Early Carboniferous)stromatolite mounds in a fore-reef slope setting, Laibin, Guangxi,South China.International Journal of Earth Sciences (Geologische Rundschau), 99: 443-458.
Siegmund, H., Trappe, J., Oschmann, W., 2002.Sequence stratigraphic and genetic aspects of the Tournaisian “Liegender Alaunschiefer”.Geologische Rundschau, 91: 934-949.
Sokal, R.R., Michener, C.D., 1958.A statistical method for evaluating systematic relationships.University of Michigan Museum of Paleontology Papers on Paleontology, 38: 1409-1438.
Somerville, I.D., 2008.Biostratigraphic zonation and correlation of Mississippian rocks in Western Europe: Some case studies in the late Visean/Serpukhovian.Geological Journal, 43: 209-240.
Somerville, I.D., Cózar, P., Said, I., Vachard, D., Medina-Varea, P.,Rodríguez, S., 2013.Palaeobiogeographical constraints on the distribution of foraminifers and rugose corals in the Carboniferous Tindouf Basin, South Morocco.Journal of Palaeogeography,2(1): 1-18.
Vachard, D., 1977.Etude stratigraphique et micropaléontologique(algues et foraminifères)du Viséen de la Montagne Noire (Hérault, France).Mémoires del’ Institut de Géologique Université de Louvain, 29: 111-195.
Vachard, D., Fadli, D., 1991.Foraminifères, algues et pseudo-algues du Viséen du Massif des Mdakra (Maroc).Annales de la Société géologique du Nord, 109(3/4): 185-191.
Vachard, D., Pille, L., Gaillot, J., 2010.Palaeozoic Foraminifera:Systematics, palaeoecology and responses to global changes.Revue de Micropaléontologie, 53: 209-254.
Vachard, D., Tahiri, A., 1991.Foraminifères, algues et pseudo-algues du Viséen de la région d’Oulmès (Maroc).Géologie Méditerranéenne, 18(1-2): 21-41.
Ward, J.H., 1963.Hierachical grouping to optimize an objective function.Journal of the American Statistical Association, 58:236-244.
Waters, C.N., Dean, M.T., Jones, N.S., Somerville, I.D., 2011.Northumberland Trough and Solway Basin.In: Waters, C., (ed).A revised correlation of Carboniferous rocks in the British Isles.Geological Society of London Special Report, 26: 89-95.
Webb, G.E., 2000.The palaeobiogeography of Eastern Australian Lower Carboniferous corals.Historical Biology, 15: 91-119.
Xu, B., 1987.Biogeographical provincialism and biomigratory barriers during the Permo-Carboniferous in China.GeoJournal, 14(4):429-434.
 Journal of Palaeogeography2014年1期
Journal of Palaeogeography2014年1期
- Journal of Palaeogeography的其它文章
- Congratulations on success of the 1st International Palaeogeography Conference
- 19th International Sedimentological Congress 2014 in Geneva: Theme 12—Palaeogeography
- Quantitative lithofacies palaeogeography
- The ichnofacies and ichnoassemblages in terrestrial deposits of China
- Patterns of fossil distributions within their environmental context from the Middle Triassic in South Canyon, Central Nevada, USA
- Foraminiferal biostratigraphy of lignite mines of Kutch, India: Age of lignite and fossil vertebrates