
The ichnofacies and ichnoassemblages in terrestrial deposits of China
2014-09-27 08:05:22BinHuYuanYuanWangHuiBoSong
Journal of Palaeogeography 2014年1期
Bin Hu , Yuan‑Yuan Wang Hui‑Bo Song
1.Institute of Resources and Environment, Henan Polytechnic University, Jiaozuo 454003, China
2.Key Laboratory of Biogenic Traces and Sedimentary Minerals of Henan Province, Jiaozuo 454003, China
1 Introduction
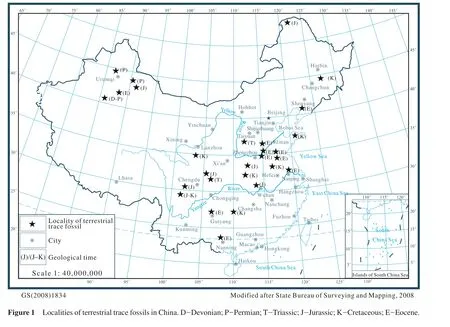
Mesozoic-Cenozoic terrestrial deposits of China are extensively exposed.Large- and middle-scale depositional basins are distributed in the northeast, north,northwest, southwest and southeast of China.It is well known that the Mesozoic-Cenozoic basins are very important source areas of the fossil-fuel and mineral resources in China.A lot of petroleum, natural gas and coal resources developed in the Songliao Basin, North China Basin, Jianghan Basin, Sichuan Basin, Erdos Basin, Tarim Basin and Qaidam Basin.Within the last 30 years,there has been great progress in terrestrial ichnological research in China, with large numbers of trace fossils in fl uvial or lacustrine sedimentary environments found in these basins (Figure 1).Until now, the research achievements mainly come from the Jiyuan Basin in the western Henan Province (Wu, 1985; Bromley, 1996; Buatoiset al., 1996; Huet al., 2004; Zhanget al., 2005; Qiet al., 2007a), the Hefei Basin of Anhui Province (Jin,1987), the Mesozoic Basin of Sichuan Province (Huet al., 1991; Hu and Wu, 1993), the Jiyang Depression of Shandong Province (Wang, 1996; Lüet al., 2003; Huet al., 2005, 2006), the Dongpu Sag of eastern Henan Province (Wuet al., 1999), the Liaohe Basin of northeastern China (Liet al., 1997; Luet al., 2003b), the Tuha Basin in Xinjiang Uygur Autonomous Region (Zhanget al., 2000), the Erdos Mesozoic Basin (Luet al., 2003a),Xixia Basin of southwestern Henan Province (Zhanget al., 2004; Songet al., 2008)and Miyang Sag of southern Henan Province (Qiet al., 2007b).These studies primarily concentrate on the genera and species description of trace fossils, the ichnoassemblage and ichnofabric analysis, paleoenvironmental reconstruction and trace fossil application in energy exploration (Huet al., 1997, 2002).From these studies, 36 kinds of ichnoassemblages and 22 ichnofabrics have been identif i ed and established,especially, the ichnoassemblages in deep-water lake facies of China (Huet al., 2007)are different from those in other areas of the world.In addition, the terrestrial trace fossils include vertebrate footprints and dinosaur egg fossils (Duet al., 2002; Wang and Feng, 2008); fish swimming traces (Luet al., 2003a)and insect trace fossils in the wood substrate (Guo, 1991).Nowadays, the widely accepted terrestrial ichnofacies are theScoyeniaichnofacies,Termitichnusichnofacies andMermiaichnofacies (Buatois and Mángano, 1995).The terrestrial trace fossils found in China mostly belong to these three ichnofacies.
1.1 Scoyenia ichnofacies
Previous studies indicate that the representative trace fossils in theScoyeniaichnofacies areScoyenia,Beaconites,Ancorichnus,Taenidium,Palaeophycus,Cruziana,Rusophycus,Diplichnites,Umfolozia,Merostomichnus,Acripes,Siskemiaand various footprints and crawling traces.At present, this ichnofacies is distinguished by ichnoassemblages withScoyenia gracilisandAncorichnus coronusprimarily, or other burrows of similar ethology.It is characterized by some burrows with crescent f i lling structures, and arthropod footprints are an important composition too.It is developed in the transitional zone between land and shallow water or temporarily submerged,which are characterized by nonmarine low energy environment periodically exposed or extremely shallow water,specif i cally the f l uvial f l oodplain, lake shore zone, dunes within a temporary lake and the supratidal zone.This ichnofacies is always associated with mud cracks, rain prints and rhizoliths.
1.2 Termitichnus ichnofacies
Based on the study of trace fossils in Late Pleistocene fl uvial deposits of Namibia (Smithet al., 1993), this ichnofacies is represented by ichnoassemblages in terrestrial settings, especially paleosol deposits.Initially, this ichnofacies was proposed as an ichnosubfacies ofScoyeniaichnofacies.Generally, trace fossils of this ichnofacies include dwelling, feeding traces, reproduction structures,rhizoliths, footprints and coprolites, and the main ichnogenera areTermitichnus,Edaphichnium,Scaphichnum,Celliforma,Macanopsis,Ichnogyrus,Coprinisphaera,Pallichnus,Daimonelix,Chubutoithes,Uruguay,AttaichnusandSyntermesichnus, exhibiting high to middle diversity.TheTermitichnusichnofacies is always distributed in an entirely terrestrial environment, and includes all kinds of paleosols, erosion surfaces, fl uvial plain, abandoned channel, fl oodplain, and dunes on beach with soft sandy or muddy substrates.
1.3 Mermia ichnofacies
On the basis of ichnoassemblage characteristics in turbidites of the Carboniferous lacustrine deposits in Argentina of South America, and integrated with ichnoassemblages in lacustrine deposits of different ages from other areas in the world, Buatois and Mángano (1995)established theMermiaichnofacies to represent a kind of ichnocoenoses in an entirely subaqueous lacustrine environment.The ichnofacies mainly consists of horizontal grazing or feeding traces made by benthic deposit feeders, while crawling traces are less prominent.The trace fossils of the ichnofacies show medium to high diversity and abundance, and the typical ichnogenera areMermia,Gordia,Helminthopsis,Helminthoidichnites,Vagorichnus,Cochlichnus,Tuberculichnus,Planolites,Treptichnus,Maculichnus,UndichnaandPalaeophycus.Generally, the trace fossils of this ichnofacies are preserved in pre- and post-event deposits and developed in soft substrate associated with fine-grained sediments in oxygen-rich and low-energy lacustrine environment.It represents a stable condition with low depositional rates, which is disturbed by episodic turbidity currents and underf l ows.In event deposits, parallel and graded beds, and tool marks are common.
It should be noted that there are some similar trace fossils between terrestrial and marine facies.This phenomenon may ref l ect similarities in trace makers and trace making behaviors in the two different realms.For example,Cruzianais produced by trilobites in marine environments, whereasIsopodichnusis produced byConchostrachansin fresh water environments.Both are produced by organisms with a symmetrical structure.Because of their similar trace morphology, some scholars equateIsopodichnuswithCruziana.Hasiotis (1993)identif i ed lobster burrows after analyzing the trace fossils in the Triassic fl uvial and lacustrine deposits.The morphology of lobster burrows is similar toPsilonichnusmade by arthropods or crabs on a beach.However, lobster burrows show some unique characteristics generated in non-marine environments.Therefore, a new ichnogenus namedCamborygmais established for lobster burrows.
Compared with marine environments, the composition of trace fossils in terrestrial deposits is obviously different, with different ichnoassemblages yielded in different lithofacies.There are vast contrasts in environmental factors between marine and terrestrial environments.Most of the terrestrial fauna are affected by temperature, precipitation, soil humidity and water depth, and the ethology of trace makers change with subenvironments.Terrestrial environments are signif i cantly inf l uenced by climatic conditions, and water level, clarity, salinity, energy and oxygen content generally f l uctuate with seasonal change.Therefore, trace fossil and ichnoassemblage characteristics are affected by environmental parameters such as the water level, soil or substrate properties, food webs, nutrient levels and environmental energy with a terrestrial imprint.
2 lchnoassemblages in f l uvial deposits in China
The f l uvial depositional system is an important constituent of terrestrial facies, which includes straight channel, braided, meandering and anastomosed stream.The fl uvial sedimentary facies includes four kinds of subfacies,namely, channel subfacies (including channel lag sedimentary microfacies and point bar sedimentary or heart bar sedimentary microfacies), embankment subfacies(composed of natural levee and crevasse splay microfacies), f l oodplain (f l ood basin)subfacies (made up of alluvial flat, overbank lake and back-swamp microfacies)and oxbow lake subfacies.All these subfacies generate paleosols that are disturbed by trace makers and plants,and are inf l uenced by f l uctuations of water level.In these deposits, some of the most complex trace fossils are produced by plants and insects.Each subfacies is characterized by a unique water level, ecological environment, and biotic community.Generally, trace makers of different biotic communities have a different ethology, resulting in different ichnoassemblages.On the other hand, some trace makers from different biotic communities have a similar ethology in the same environment, producing similar ichnoassemblages.For example, the burrows made by insects living in aquatic and terrestrial environments may possess similar morphologic characteristics.
There are many types of trace makers in terrestrial facies.Chamberlain (1975)and Ratcliffe and Fagerstrom(1980)summarized different kinds of invertebrate dwelling in the f l uvial and related environments.These organisms include four types as follows: (1)arthropods, like spiders, ants, termites, wasps, bees, crickets, centipedes,caddisf l ies, f l ies, and other kinds of insects in terrestrial and aquatic environments; (2)worms, like earthworms,leeches and nematodes in terrestrial and aquatic environments; (3)molluscs, like gastropods and mussels in terrestrial and aquatic environments; (4)crustaceans in terrestrial and aquatic environments, including crayfish, shrimps,crabs, woodlouses, scorpions, ostracods and amphipods.In addition, some plant roots are closely related to many terrestrial and aquatic invertebrates, which may provide shelter and food for these animals and help to build burrows.
Some complex burrow systems of social and solitary insects are often generated in dehydrated deposits (low water level).The social termites can make elaborate nests which are often developed in dry channel, f l oodplain and distal f l oodplain environments, and most of these burrows commonly occur in fully dehydrated soil.Changes in nest structure are dependent on moisture in soil and air.
Beetles always live in wet to dry environments and produce shelter nests for larvae and dwelling nests.Their nests can be useful for the determination of water level in the past.For example, beetles prefer to burrow in mud.They produce horizontal burrows near land-water interface; however, scarabs prefer to burrow in dehydrated soils and deciduous grounds (Chamberlain, 1975).
The fossil nests of social or solitary hornets and bees found in the Cretaceous of North America, South America and Africa (Bown, 1982; Bown and Ratcliffe, 1988;Genise and Bown, 1994, 1996)are similar to modern ones.These insects prefer to burrow in fully dehydrated soils or sediments.
Worms, mussels and gastropods can also indicate the change of water level.Earthworms live in low- to medium-humid soils above the water level.After rain, many earthworms moves to grass or road to avoid drowning.Other small worms, such as annelids and nematodes, are wholly aquatic and always appear in high water level environments such as stream, river and pond.Mussels and gastropods are aquatic animals, and often burrow during the dry season, afterwards, while the water is subsiding,a series of directivity creeping traces are produced on the ground or sediment surface (Chamberlain, 1975).
Aquatic and terrestrial crustaceans, especially the lobsters, can be used to identify water depth and indicate paleoenvironment.The earliest lobster fossils are found in the Triassic.According to how long burrows are occupied, complexity of burrow structures and the relative water level of modern lobsters, the lobsters burrows can be divided into three types: (1)the most complex burrows, where lobsters live in and is not directly joined to open water; (2)the secondary complex burrows, where the living time in burrows is reduced and these burrows are directly joined to open water; (3)the simple burrows,where most living time of lobsters is in open water and only simple burrows to escape and breed are produced.The morphology of lobster burrows could ref l ect the depth of water as a modern analogy (shrimps and crabs)in transitional zone.The morphology of burrows and their distance below the sediment-water interface also ref l ect the types of lobster in speci fi ed areas in a sedimentary system.For example, lobster burrows in the upper member of the Triassic Chinle Formation in northeast Utah and west Colorado re fl ect these distribution characters (Hasiotis, 1993, 2002).
Above-mentioned research indicates that there are aquatic arthropods, worms, mollusks and plant roots in fl uvial depositional system and the types of trace makers and ichnoassemblages have obvious and important differences in different fl uvial sedimentary subfacies.In the channel subfacies (mainly in point bar and heart bar deposits), the main organisms are aquatic invertebrates, including gastropods, bivalves, nematodes, lobsters, crabs,insect larvae, mud-like beetles and aquatic oligochaete worms and so on.Their burrows and surface traces are distributed in deposits of point bar, heart bar or abandoned channel.In pro fi le, burrows are always distributed near or beneath water table and are nearly horizontal.Therefore, these burrows can indicate the position of the water table.In the embankment subfacies (mainly in natural levee, crevasse splay and near channel fl oodplain deposits), organisms are mostly lobsters, beetles, insect larvae, worms and plant roots.They fl uctuate with water table and intermittent fl ooding.During fl ood season, water table is high, sediment is water-rich, and environment condition is bene fi cial for aquatic or semi-aquatic organisms such as mud-like beetles, insect larvae, and aquatic worms.The crawling traces on sediment surfaces and horizontal or U-shaped burrows are produced.However,during the dry season, the water table will fall, and the environment will become conducive to the breeding of terrestrial organisms, like ants, bees, hornets and rodents,and tend to host vertical burrow construction.Traces in these two kinds of habitats can overlap or alternate vertically.In the fl oodplain subfacies (mainly distal channel and fl oodplain deposits above the water table), terrestrial organisms are common, for example, earthworms and annelids, crustaceans such as lobsters and crabs; insects such as termites, ants, bees, hornets and beetles; arthropods such as spiders; and terrestrial plants.
Up to now, a lot of fl uvial trace fossils have been found in the Mesozoic and Cenozoic terrestrial deposits of China,at least 24 ichnogenera.Most of these are feeding, grazing, dwelling, resting traces and rhizoliths.The common ichnogenera areAncorichnus,Beaconites,Cystichnium,Muensteria,Palaeophycus,Planolites,Psammichnites,Scoyenia,Steinichnus,Taenidium,Gordia,Paradidymaulichnus,Monomorphichnus,Oniscoidichnus,Pelecypodichnus,Rusophycus,Arenicolites,Cylindricum,Gastruichnus,Polykladichnus,Skolithos,Stipsellus,Thalassinoides, andTrichichnus.These trace fossils are preserved mainly as endichnia, epichnia and hypichnia in fine sandstone, siltstone, sandy mudstone and mudstone.This paper lists 6 ichnoassemblages found commonly in fl uvial sedimentary environments from the Upper Cretaceous of the Sichuan Basin (Figure 2)and Xixia Basin of western Henan Province (Figures 3, 4, 5).
2.1 Scoyenia-Rusophycus ichnoassemblage
This ichnoassemblage is characterized by grazing and crawling traces on bedding planes and endichnial feeding traces.The main trace fossils areScoyenia,Steinichnus,Cystichnium,Planolites,Gordia,Monomorphichnus,Oniscoidichnus,Paradidymaulichnus,PelecypodichnusandRusophycus.These trace fossils are mainly developed in the red-violet, thin- to medium-bedded fine sandstones and thin-bedded muddy siltstone or sandy mudstone, with medium- to small-scale cross-beddings in fine sandstone.Climbing ripple lamination, wavy bedding and horizontal bedding are common in siltstone and sandy mudstone.The associated inorganic sedimentary structures with trace fossils on bedding planes (hypichnia and epichnia)are asymmetrical ripple marks, rill marks,mud cracks, raindrop impressions and so on.With regard to the relationship of ripple marks and mud cracks, almost all the samples ref l ect the former is earlier than the latter.Some traces are distributed on the ripple marks or across the mud cracks.From the above sedimentary characters, it reveals that: (1)the paleoclimate was arid at that time; (2)the active zones of trace makers are f l oodplain along channel and shallow water swales or oxbow lakes;(3)the active substrate of trace makers is semi-consolidated, with muddy sands that were alternately dry and wet.When the substrate is wet, there are large amount of organismal activities.When the substrate is dry and mud cracks are developed, the organisms escape or burrow deep into the substrate.Obviously, the alternation is closely related with the seasonality in precipitation and fl ooding.This ichnoassemblage is commonly generated in f l oodplains along channel and shallow water swales or oxbow lakes (Figure 2B, 2C).
2.2 Skolithos-Arenicolites ichnoassemblage
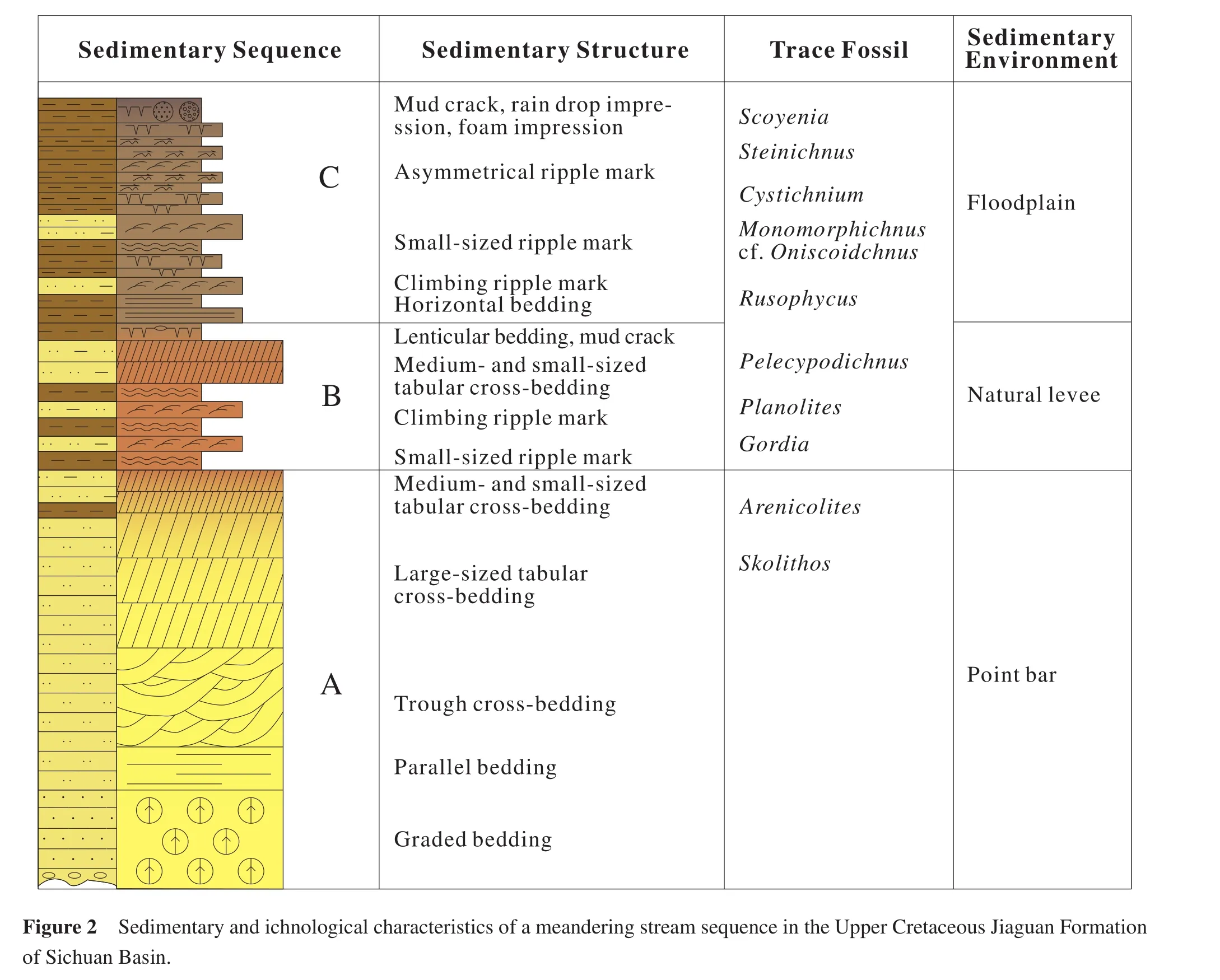
This ichnoassemblage is characterized by vertical dwelling burrows in high abundance but with low diversity.The main trace fossils areArenicolitesandSkolithos.A small amount of irregularThalassinoidesand boring traces may be found locally.This ichnoassemblage is commonly developed in medium- to thick-bedded fine sandstones with parallel bedding or large-scale tabular cross-bedding.A few muddy conglomerates can be found in the middle to lower part of sandstone beds, even the lag beds, scouring structures such as f l ute casts and tool marks are present at the bottom of sandstone beds.The monotonous vertical dwelling burrows mainly appear in the high-energy sandy bar deposits with consolidated substrates.The trace makers are suspension-feeders.It ref l ects a high energy sandy bar (heart bar or point bar)environments in the channel(Figure 2A).
2.3 Gastruichnus-Palaeophycus ichnoassemblage
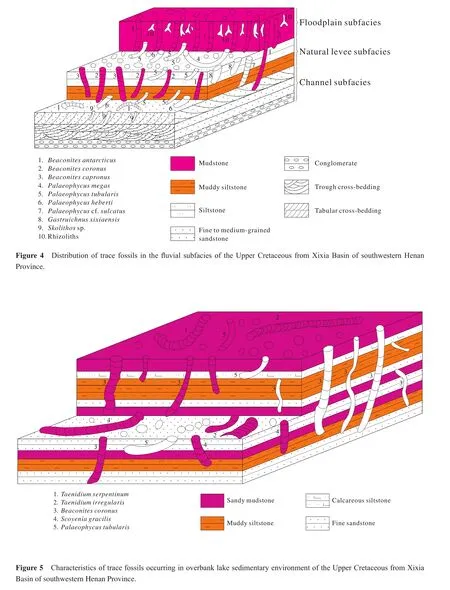
This ichnoassemblage consists of trace fossils with higher abundance and diversity, with common composition such asGastruichnus xixiaensis,Palaeophycus tubularis,Palaeophycus megas,Skolithosisp.,Beaconites capronusandBeaconites coronusmainly developed in the medium- to thick-bedded fine sandstone.It indicates the high oxygen content in water.Among these trace fossils,Gastruichnus xixiaensisandBeaconites coronusare dwelling (associated with feeding activity)burrows, arranged almost perpendicular (high angle)to the bedding plane, revealing a medium-energy water current.The ichnogenusPalaeophycusis a kind of large feeding burrows distributed along the bedding planes of fine sandstone with parallel bedding and gentle wavy bedding, which indicates the water body is stable and beneficial for the trace maker’s activity.This ichnoassemblage is commonly developed in channel-levee environment (Figure 4).
2.4 Beaconites-Taenidium ichnoassemblage
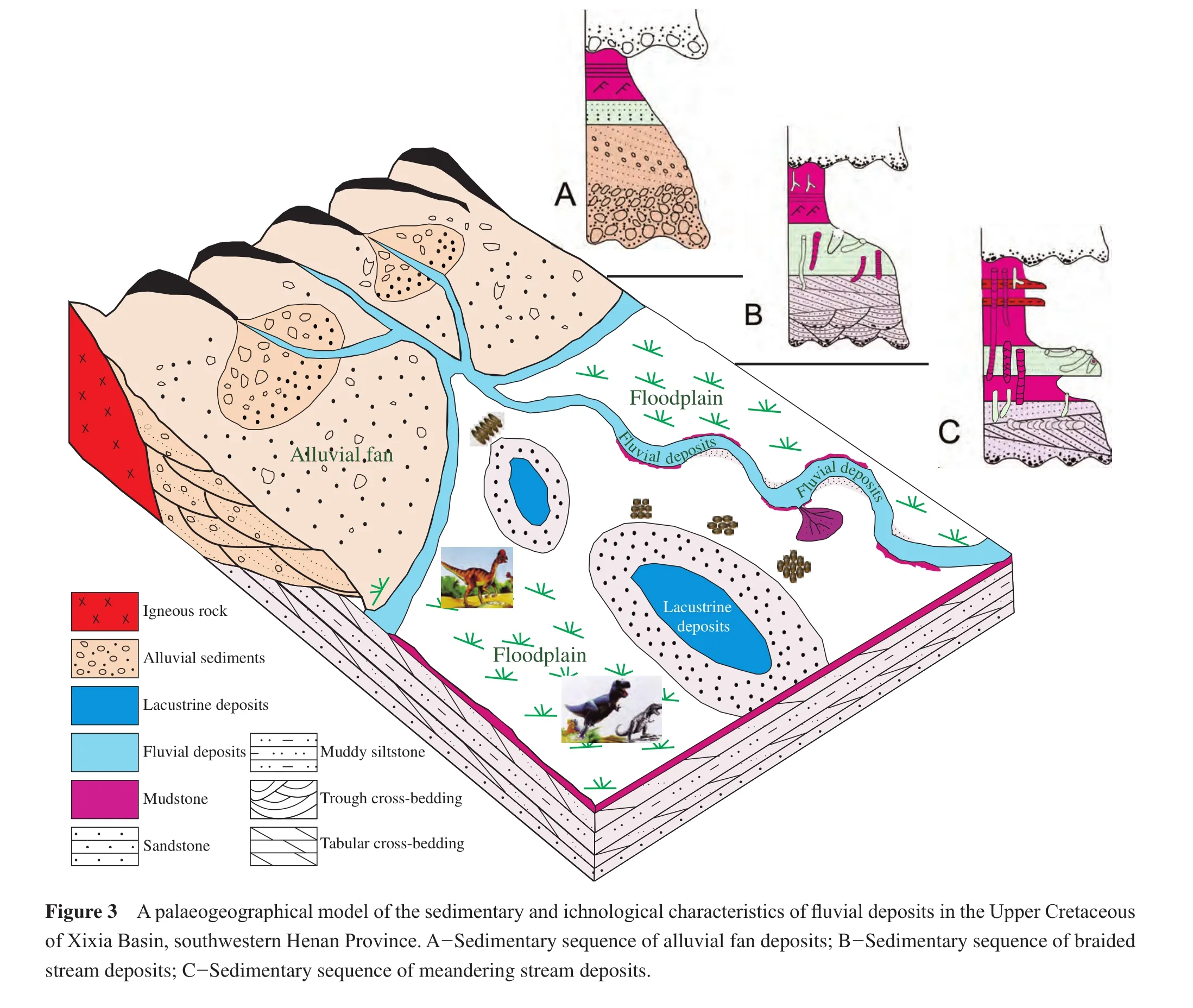
The ichnoassemblage is marked byBeaconites coronus,Beaconites antarcticus,Taenidium irregularisandPalaeophycus tubularis.These trace fossils, with high abundance and low diversity, are mostly burrows with meniscus backf i ll structures and are preserved in purplish thin muddy siltstone intercalated with reddish mudstone bands associated with wavy or discontinuously wavy bedding.Based on these characters, this ichnoassemblage is thought to occur in embankment (including crevasse splay)sedimentary environments (Figure 4).
2.5 Beaconites-Rhizoliths ichnoassemblage
This ichnoassemblage is mainly composed ofBeaconites coronus,Steinichnuscf.largusand rhizoliths.These trace fossils have a lower abundance and diversity and occur in the red-violet silty mudstone locally containing ginger stones, which is interpreted as an arid climatic environment.The rhizoliths indicate a terrestrial environment and temporary or extremely shallow water conditions.Therefore, this ichnoassemblage is hypothesized to form in a f l oodplain sedimentary environment (Figure 4).
2.6 Beaconites-Scoyenia ichnoassemblage
The main trace fossils in this ichnoassemblage are feeding traces likeBeaconites coronus,Beaconites antarcticus,Taenidium irregularis,Palaeophycus tubularis,Palaeophycuscf.sulcatusandScoyenia gracilis.Generally, the trace fossils with high abundance and medium diversity are arranged along or inclined at high angle to the bedding plane.This ichnoassemblage is mainly developed in thinbedded grayish green siltstone, silty mudstone or muddy siltstone, and is always associated with ripple marks and wavy or gentle wavy bedding, which ref l ects low energy conditions of water.Thus, this ichnoassemblage is commonly thought to appear in the periodically exposed overbank lake (extremely shallow lacustrine)sedimentary en-vironment (Figure 5).
3 Ichnoassemblages in lacustrine deposits in China
The distribution of ichnocoenoses in lacustrine facies is closely related to the position occupied by trace makers within the lake.Generally, according to the size and open degree of a lake basin, f l uctuation of water table and nutrient levels of lake water, the distribution of trace makers in lacustrine deposits are divided into three ecological environment zones,i.e., eulittoral (lakeshore), littoral (lake beach)and inner lake (permanently lacustrine).
The eulittoral area is located on the land side of the lakeshore zone.In the deposit area, when water table declines to very low levels, the substrate will be exposed and desiccated, simultaneously, the plants, terrestrial-like and hydrophilic organisms will grow, whereas the deposit area will also be affected by winds and waves when a strong storm comes.The main traces include burrows and grazing traces produced by tiger beetles, mud-like beetles, mole crickets, crickets, lobsters and bivalves.On the lakeshore, footprints and resting traces may be produced by birds, mammals and reptiles, and the morphology of burrows there is often affected by the water table.
The littoral area is located between the shore saturated zone and the lowest water table of lake basins.The trace makers are affected by the waves, duration of desiccation and nutrient level of lake water.Many organisms occur in high water table zone, like deposit feeding beetles, bivalves, oligochaetae worms, lobsters and crabs.These animals can produce large, shallow and complex burrows and may migrate between eulittoral and littoral areas, which tend to form overlapping ichnoassemblages with the inf l uence of seasonal f l uctuations.
The inner lake (permanently lacustrine)area is located in the deeper lake area rarely affected by f l uctuations of the water table.Many benthonic fauna live in the deeper lake.Common organisms include larvae of aquatic insects,worms, arthropods and mollusks.Based on the distribution of organisms in modern lakes, Chamberlain (1975)discovered their distribution is related to bathymetry.Most organisms live at 10-20 m depth, like leeches, platyhelminthes, polychaetes, isopods, gastropods and larvae of dragonf l ies and trichoptera.The larvae of bivalves and midges can live at depths of dozens of meters in water.Meanwhile, amphipods and oligochaete worms may be able to live at any lake depth.
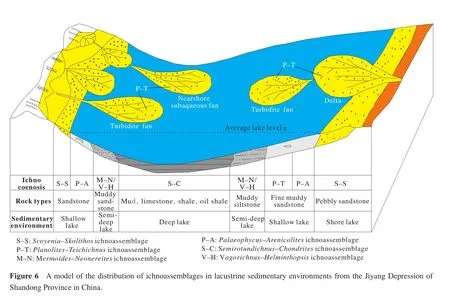
Lacustrine ichnology has been much improved with on-going terrestrial resource exploration.Since the 1990s,the study of reservoir sedimentology in the Mesozoic-Cenozoic terrestrial basins has been developing rapidly in China, especially in some large basins containing petroleum and natural gas such as the Jiyang Depression of Shandong Province, Liaohe Basin of northeastern China,Erdos Basin and Tuha Basin of northwestern China, and the Dongpu Depression of Henan Province.Meanwhile,the sedimentological research greatly promoted the ichnological studies, with a large number of lacustrine trace fossils having been discovered in the Jiyang Depression of Shandong Province, Liaohe Basin of northeastern China, Jiyuan Basin and Puyang Sag of Henan Province.There have been at least 59 ichnogenera identif i ed.The major trace makers are arthropods, mollusks and worms.The common ichnogenera include feeding tracesAncorichnus,Beaconites,Chondrites,Cystichnium,Fuersichnus,Glockeria,Muensteria,Palaeophycus,Phycodes,Paracanthorhaphe,Planolites,Psammichnites,Scoyenia,Steinichnus,Taenidium,Terebellina,Teichichnus,Tuberculichnus; grazing tracesCochlichnus,Helminthopsis,Mermoides,Gordia,Helminthoidichnites,Multilagueichnus,Neonereites,Paradidymaulichnus,Pilichnus,Patellarisichnus,Semirotundichnus,Vagorichnus; crawling tracesCruziana,Diplichnites,Monomorphichnus,Oniscoidichnus; resting tracesLiaohusichnus,Rusophycus,Pelecypodichnus,Sagittichnus;and dwelling tracesArenicolites,Bifungites,Brevitubus,Cylindricum,Linearis,Margaritichnus,Monocraterion,Arenicolites,Diplocraterion,Gastruichnus,Ophiomorpha,Polykladichnus,Skolithos,Stipsellus,Thalassinoides,Tigillites,Trichichnus, coproliteFarreina,Lumbricariaand rhizoliths.These trace fossils are preserved as endichnia, epichnia and hypichnia, and generated in fine sandstone, siltstone, sandy mudstone and mudstone.Based on the research of lacustrine trace fossils in these terrestrial sedimentary basins, six types of ichnoassemblages are considered to be particularly common.Their composition and environmental distribution are described as follows (Figure 6).
3.1 Scoyenia-Skolithos ichnoassemblage
The ichnoassemblage is made up ofScoyenia,Beaconites,Taenidium,Planolites,Skolithos,Thalassinoides,Palaeophycus,Cystichnium,Gordia,Sagittichnus,Favreinaand rhizoliths.They are mainly feeding, dwelling and crawling traces preserved epichnia and endichnia on bedding planes of sandstone, siltstone and sandy mudstone.The ichnoassemblage is always developed in periodically exposed or extremely shallow lakeshore and interdistributary bays of lake deltaic plain under drought or semiarid climate conditions.
3.2 Palaeophycus-Arenicolites ichnoassemblage
The common ichnogenera in this ichnoassemblage arePalaeophycus,Arenicolites,Skolithos,Cylindricum,Polykladichnus,Thalassinoides,Ophiomorpha,Diplocraterion,Trichichnus,Tigillites,Beaconites,AncorichnusandPlanolites.Most of them are vertical or inclined tubes and appear in groups.The dominant ethology is dwelling traces, secondly feeding traces, and occasional escape traces in sandstones.These trace fossils are preserved as endichnia, and are always produced in fine- to medium-grained sandstone associated with cross-beddings, sometimes in laminated siltstone and sandy mudstone.This ichnoassemblage is commonly distributed from lakeshore to the upper part of shallow lake or lake delta plain to delta front.In profile, it always appears in the interbedded sandstone and mudstone of alternating lakeshore and upper shallow lake environments.
3.3 Planolites-Teichichnus ichnoassemblage
The ichnoassemblage is composed ofPlanolitesandTeichichnus, which is characterized by the dense distribution of smallPlanolites.Therefore, it is commonly referred to as thePlanolitesichnoassemblage.Other associated ichnofossils includeGordia,Helminthopsis,Brevitubus,Cruziana,Palaeophycus,Arenicolites,SagittichnusandMargaritichnus.Moreover, a small amount ofChondritesare found, which indicate oxygen-depleted environments.These trace fossils mostly are feeding and crawling traces (a minor amount of dwelling traces)preserved as endichnia and epichnia in intercalated thinbedded dark mudstone and grey siltstone, muddy siltstone and sandy mudstone.Most burrows are parallel or gently inclined to the bedding plane and the color of burrow f i llings is lighter than surrounding rocks.The ichnoassemblage ref l ects the sedimentary environments of medium to low water energy, locally poorly oxygenated, such as the lower part of a shallow lake, restricted lake bay and the distal delta front.
3.4 Vagorichnus-Helminthopsis ichnoassemblage
This ichnoassemblage is developed in the Lower Jurassic Anyao Formation of the Jiyuan Mesozoic Basin,western Henan Province.It mainly consists of feeding and crawling traces.Common trace fossils areCochlichnus anguineus,Helminthoidichnites tenuis,Helminthop-sis abeli,Helminthopsis hieroglyphia,Monomorphichnus lineatus,Neonereites biserialis,Paracanthorhaphe togwunia,Tuberculichnus vagans, andVagorichnus anyao,most of which are hypichnia preserved in light grey calcilutite and grey yellow thin- to medium- to thick-bedded calcareous siltstone.The rocks belong to deeper lacustrine(profundal)turbidite deposits containing rare body megafossils except for a few plant debris.In addition, the trace fossilsFuersichnus,Glockeria,Gordia,MegagraptonandPlanoliteshave been found in the deeper lacustrine turbidites of the Upper Cretaceous Yaojia Formation from the Songliao Basin, northeastern China (Zhenget al., 2000).This ichnoassemblage ref l ects the deeper lacustrine (profundal)turbidite sedimentary environment.
3.5 Mermoides-Neonereites ichnoassemblage
IchnogenusMermoidesis a tiny simple irregular feeding trace found in the Shahejie Formation of Jiyang Depression, Shandong Province.Mermoidesis abundant in deeper water deposits of the Jiyang Depression and a representative of the ichnoassemblage.It is preserved as an endichnia in dark grey, thin or laminated mudstone.Generally, the burrow is parallel or slightly inclined to the bedding plane.Mermoidesis similar toMermiain size and occurrence.ButMermiashows clear ring-shaped tracks,whereasMermoidesis irregular.Common trace fossils in this ichnoassemblage areMermoides chezhenensis,Mermoides latiusculus,Pilichnus lacustris,Neonereites uniserialis,Helminthoidichnitescf.tenuis,Helminthopsis abeli,Vagorichnuscf.anyao,Multilagueichnus lingpanensis,Chondrites lacustris,Cochlichnus anguineus,Gordia marina,Planolitesichnosp.andFavreinacf.joukowskyi.These trace fossils are distributed along bedding planes,dominated by horizontal or sub-horizontal feeding or grazing traces with a small amount of branched feeding traces and some cubichnia.This ichnoassemblage is mostly developed in dark grey mudstone or shale with horizontal lamination, associated with multilayer carbonates and turbidites and some deeper water body fossils like ostracods and gastropods as well as tiny thin-shelled bivalves, fish and their scales and fin debris.Thus, this ichnoassemblage is thought to form in a quiet deep lacustrine sedimentary environment.
3.6 Semirotundichnus-Chondrites ichnoassemblage
This ichnoassemblage is composed mostly of semicircular and saucer-shaped grazing trails (Pascichnia)and Agrichnia, and is found in the Paleogene Shahejie Formation of Dongying Sag in the Jiyang Depression of Shandong Province and Miyang Sag of southern Henan Province.IchnogenusSemirotundichnusin the ichnoassemblage is a smallhorizontal trail distributed widely in the dark mudstone and silty mudstone of relatively deep lake deposits.It is a representative trace fossil in this ichnoassemblage.Common trace fossils areSemirotundichnusdongyingensis,Patellarisichnus boxingensis,Neonereitesuniserialis,Circinansichnusboxingensis,Chondrites lacustris, andPlanolitesichnosp.These trace fossils are abundant and unique in the Paleogene deep lacustrine sedimentary environments of northern China, and almost all of them are developed in dark grey, dark thin-bedded mudstone or silty mudstone with pronounced horizontal lamination.Compared with theMermoides-Neonereitesichnoassemblage, the characteristics of these trace fossils and their host rock indicate that the ichnoassemblage forms in an even deeper lacustrine sedimentary environment with a low oxygen content.
4 Conclusions
The above-mentioned discussion shows that the terrestrial ichnofossils found in China comprise 83 ichnogenera, of which 24 ichnogenera occur in f l uvial deposits and 59 ichnogenera appear in lacustrine deposits.These ichnofossils are mostly generated in Mesozoic basins (like Sichuan, Erdos, Tarim, Qaidam Basins of western China;Jiyuan and Xixia Basins of western Henan Province)and Cenozoic basins (like Jiyang Depression of eastern China and Liaohe Basin of northeastern China).
Based on composition, occurrence and distribution of ichnofossils in the subfacies of f l uvial and lacustrine facies, 36 ichnoassemblages have been proposed in papers published within the last 30 years in China.The most common groups in f l uvial facies includeScoyenia-Rusophycus,Skolithos-Arenicolites,Gastruichnus-Palaeophycus,Beaconites-Taenidium,Beaconites-Rhizoliths andBeaconites-Scoyeniaichnoassemblage that are mostly generated in the Upper Cretaceous of the Sichuan Basin of western China and Xixia Basin of western Henan Province.The most common groups in the lacustrine facies are theScoyenia-Skolithos,Palaeophycus-Arenicolites,Planolites-Teichichnus,Vagorichnus-Helminthopsis,Mermoides-Neonereites, andSemirotundichnus-Chondritesichnoassemblages that are mainly produced in the Jurassic of Jiyuan Basin of western Henan Province and the Paleogene in Jiyang Depression of Shandong Province and Liaohe Basin of northeastern China.
Acknowledgements
This paper was supported by the National Natural Science Foundation of China (No.41272117 and No.41102002)and the International Cooperation Program of Henan Province (No.124300510039).Thanks also to Prof.Zeng-Zhao Feng, Prof.Yi-Ming Gong and Prof.Shou-Ren Yang for their comments and amendments on various aspects of the manuscript.
Bown, M.T., 1982.Ichnofossils and rhizoliths of the nearshore f l uvial Jebel Quatrani Formation (Oligocene), Fayum Province,Egypt.Palaeogeography, Palaeoclimatology, Palaeoecology, 40:255-309.
Bown, T.M., Ratcliffe, B.C., 1988.The origin ofChubutolithesichnofossils from the Eocene and Oligocene Chubut Province,Argentina.Journal of Paleontology, 62: 163-167.
Bromley, R.G., 1996.Trace Fossils, Biology and Taphonomy.London: Unwin Hyman, 1-361.
Buatois, L.A., Mángano, M.G., 1995.The paleoenvironmental and paleoecological signif i cance of the lacustrineMermiaichnofacies: An archetypical subaqueous nonmarine trace fossil assemblage.Ichnos, 4: 151-161.
Buatois, L.A., Mángano, M.G., Wu, X.T., Zhang, G.C., 1996.Trace fossils from Jurassic lacustrine turbidites of the Anyao Formation (Central China)and their environmental and evolutionary signif i cance.Ichnos, 4: 287-303.
Chamberlain, C.K., 1975.Recent lebensspuren in nonmarine aquatic environments.In: Frey, R.W., (ed).The Study of Trace Fossils: A Synthesis of Principles, Problems and Procedures in Ichnology.New York: Springer-Verlag, 431-458.
Du, Y.S., Li, D.Q., Peng, B.X., Lei, R.L., 2002.Largescale dinosaur footprints of Sauropod from Yanguoxia, Yongjing county,Gansu Province.Earth Science—Journal of China University of Geosciences, 27(4): 367-372 (in Chinese with English abstract).
Genise, J.E., Bown, T.M., 1994.New trace fossils of termites (Insecta: Isoptera)from the late Eocene-early Miocene of Egypt,and the reconstruction of ancient sopteran social behavior.Ichnos, 3: 155-183.
Genise, J.F., Bown, T.M., 1996.Uruguay Roselli1938 andRosellichnus, n.ichnogenus: Two ichnogenera for clusters of fossil bee cells.Ichnos, 4: 199-217.
Guo, S.X., 1991.A Miocene trace fossil of insect from the Shanwang Formation in Linqu, Shandong.Acta Palaeontologica Sinica, 30(6): 739-742 (in Chinese with English abstract).
Hasiotis, S.T., 1993.Ichnology of Triassic and Holocene cambarid crayfish of North America: An overview of burrowing behavior and morphology as ref l ected by their burrow morphologies in the geological record.Freshwater Crayfish, 9: 407-418.
Hasiotis, S.T., 2002.Continental Trace Fossil short course Number 51, SEPM, Tulsa, OK, 134.
Hu, B., Jiang, Z.X., Qi, Y.A., Zhang, G.C., 2006.Trace fossils of deeper lacustrine deposits in the Paleogene Shahejie Formation of Jiyang depression, Shandong Province, eastern China.Acta Palaeontologica Sinica, 45(1): 83-94 (in Chinese with English abstract).
Hu, B., Jiang, Z.X., Qi, Y.A., Zhang, G.C., 2007.Paleogene ichnocoenoses of lacustrine deposits in Jiyang Depression of Shandong Province, Eastern China.Journal of China University of Geosciences, 18: 404-405.
Hu, B., Qi, Y.A., Zhang, G.C., Jiang, Z.X., 2002.Ichnocoenoses in the Mesozoic and Cenozoic continental deposits in China.Acta Sedimentologica Sinica, 20(4): 574-581 (in Chinese with English abstract).
Hu, B., Wang, G.Z., Qi, Y.A., 1997.Theory and Application of Ichnology.Xuzhou (Jiangsu): China University of Mining Press,1-209 (in Chinese).
Hu, B., Wang, H.F., Zhang, H.M., Niu, Y.B., Bai, Z.K., 2005.Ichnocoenoses and their sedimentary environments of the Shahejie Formation of Paleoene in Dongying Sag, Jiyang Depression,Shandong Province.Journal of Henan Polytechnic University,24(3): 186-191 (in Chinese with English abstract).
Hu, B., Wu, X.T., 1993.Ichnocoenoses in the f l uvial deposits of the Late Cretaceous Jiaguan Formation in Emei, western Sichuan.Acta Palaeontologica Sinica, 32(4): 478-489 (in Chinese with English abstract).
Hu, B., Wu, X.T., Pan, L.M., 1991.Ichnocoenoses of the Late Paleozoic and Mesozoic f l uvial deposits of Emei area, western Sichuan, China.Acta Sedimentologica Sinica, 9(4): 128-135 (in Chinese with English abstract).
Hu, B., Zhang, G.C., Guo, W.X., Qi, Y.A., Zhang, L.W., 2004.Stipsellusichnofabric in f l uvial deposits of the Tuanzhuang Formation from Jiyuan, western Henan.Coal Geology & Exploration,32(3): 1-4 (in Chinese with English abstract).
Jin, F.Q., 1987.Continental trace fossilScoyeniafound in the Jurassic red beds of Hefei basin.Journal of Hefei University of Technology, 9: 91-95 (in Chinese with English abstract).
Li, Y.X., Lu, Z.S., Wang, D., 1997.Continental Trace Fossils and Sedimentary Environments in Liaohe Basin.Beijing: Petroleum Industry Press, 1-145 (in Chinese with English abstract).
Lü, X.X., Hu, B., Jiang, Z.X., Chen, S.Y., Li, S.J., 2003.Ichnocoeneses and their sedimentary environments of the Paleogene Shahejie Formation in Chezhen and Zhanhua sags, Jiyang depression, Shandong Province.Journal of Palaeogeography, 5(2):187-196 (in Chinese with English abstract).
Lu, Z.S., Hao, C.K., Chen, B., 2003a.New evidences of Late Triassic fish swimming traces in Hengshan county, Shanxi Province,China.Acta Palaeontologica Sinica, 42(2): 266-273 (in Chinese with English abstract).
Lu, Z.S., Hao, C.K., Ma, H.B., Zhang, X.H., 2003b.Continental ichnofabric types in the Liaohe oilfield and their environmental interpretation, China.Acta Geologica Sinica, 77(1): 9-15 (in Chinese with English abstract).
Qi, Y.A., Hu, B., Zhang, G.C., Gong, Y.M., 2007a.Ichnofabrics and their environmental interpretation from the Middle Triassic Youfangzhuang Formation, Jiyuan Region, western Henan Province.Acta Sedimentologica Sinica, 25(3): 372-379 (in Chinese with English abstract).
Qi, Y.A., Zeng, G.Y., Hu, B., Zhang, G.C., Gong, Y.M., 2007b.Trace fossil assemblages and their environmental signif i cance from Hetaoyuan Formation of Paleogene in Biyang depression of Henan Province.Acta Palaeontologica Sinica, 46(4): 441-452(in Chinese with English abstract).
Ratcliffe, B.C., Fagerstrom, J.A., 1980.Invertebrate lebensspuren of Holocene f l oodplains: Their morphology, origin and paleoecological signif i cance.Journal of Paleontology, 54: 614-630.
Smith, R.M.H., Mason, T.R., Ward, J.D., 1993.Flash-f l ood sediments and ichnofacies of the Late Pleistocene Homeb Silts,Kuiseb River, Namibia.Sedimentary Geology, 85: 559-579.
Song, H.B., Hu, B., Wang, D.Y., Liu, S.X., Gao, Y., Zhong, M.Y., 2008.Sedimentary characteristics and environments of the Gaogou Formation of Upper Cretaceous in Xixia Basin, Henan.Petroleum Geology and Engineering, 22(4): 1-5 (in Chinese with English abstract).
Wang, D.Y., Feng, J.C., 2008.Dinosaur eggs and skeletons from Henan Province in China.Beijing: Geological Publishing House,1-320 (in Chinese with English abstract).
Wang, H.Z., 1996.Semiquantitative analysis of bioturbations from the Shahejie Formation in the Dongying depression, Shandong.Scientia Geologica Sinica, 31(3): 287-295 (in Chinese with English abstract).
Wu, X.T., 1985.Trace fossils from the turbidites in Jiyuan-Yima basin of the western Henan and their environmental signif i cance.Acta Sedimentologica Sinica, 3(3): 23-31 (in Chinese with English abstract).
Wu, X.T., Lin, Y.L., Pan, J.N., 1999.Signif i cance of ichnofacies and its corresponding well log patterns of the Shahejie Formation in Dongpu depression.Journal of Palaeogeography, 1(3): 50-64(in Chinese with English abstract).
Zhang, G.C., Guo, W.X., Zeng, Y.F., 2004.Ichnofabrics from the Upper Cretaceous f l uvial and lacustrine deposits in Xixia basin,Henan.Journal of Palaeogeography, 6(4): 434-441 (in Chinese with English abstract).
Zhang, G.C., Zeng, Y.F., Buatois, L.A., Mangono, M.G., 2005.Lacustrine deposits and associated trace fossils in the upper part of the Tanzhuang Formation (T2-3), Jiyuan Basin, Henan Province.Acta Sedimentologica Sinica, 23(1): 100-107 (in Chinese with English abstract).
Zhang, J.P., Xue, S.H., Yang, S.P., Wu, Y.Y., 2000.Trace fossils made by lacustrine animals found in the Jurassic from Tulufan-Hami basin of Xinjiang and their palaeoenvironmental signif icance.Geoscience, 14(3): 373-378 (in Chinese with English abstract).
Zheng, Y.L., Chen, C.R., Huang, Q.H., 2000.Trace fossils of Yaojia Formation in Songliao Basin and their environmental implication.Daqing Oil Geology and Exploration, 19(6): 12-14 (in Chinese with English abstract).
 Journal of Palaeogeography2014年1期
Journal of Palaeogeography2014年1期
- Journal of Palaeogeography的其它文章
- Provenance records of the North Jiangsu Basin,East China: Zircon U-Pb geochronology and geochemistry from the Paleogene Dainan Formation in the Gaoyou Sag
- Foraminiferal biostratigraphy of lignite mines of Kutch, India: Age of lignite and fossil vertebrates
- Patterns of fossil distributions within their environmental context from the Middle Triassic in South Canyon, Central Nevada, USA
- Diversity patterns and palaeobiogeographical relationships of latest Devonian-Lower Carboniferous foraminifers from South China: What is global,what is local?
- Quantitative lithofacies palaeogeography
- 19th International Sedimentological Congress 2014 in Geneva: Theme 12—Palaeogeography