
Patterns of fossil distributions within their environmental context from the Middle Triassic in South Canyon, Central Nevada, USA
2014-09-27 08:05PedroMonarrezNicoleBonuso
Journal of Palaeogeography 2014年1期
Pedro M.Monarrez, Nicole Bonuso
Department of Geological Sciences, California State University, Fullerton, CA 92834-6850, USA
1 Introduction*
The Middle Triassic, particularly the Anisian, records an interesting and important evolutionary phase for marine biota.During this time survivors of the end-Permian mass extinction, as well as several lineages of Lazarus taxa,began to diversify and obtain their more modern adaptive characteristics (Vermeij, 1977; Miller and Sepkoski,1988; McRoberts, 2001; Sepkoski, 2002).This radiation is thought to mark the onset of substantial reorganization of marine communities leading up to the marine Mesozoic Revolution (MMR)— the establishment of a marine fauna with modern characteristics (McRoberts, 2001).Previous work derived their diversif i cation patterns from regional and global scales; however, to fully understand how this reorganization initiates, we must examine communities on the local scale.
Recent work focusing on the local scale examines the faunal abundance patterns throughout the Middle and Late Triassic with a primary focus on Europe and China (Pálfy,1990; Fenget al., 2001; Komatsuet al., 2004; Payneetal., 2006; Bonuso and Bottjer, 2008; Posenato, 2008; Diedrich, 2009; Chenet al., 2010).Few local scale studies exist in western North America perhaps due to limited Middle to Upper Triassic marine rock exposures as a consequence of their origination on an active plate margin.Although sparse, Middle to Upper Triassic sections exist in Central Nevada.Researchers primarily studied these sections for their biostratigraphic signif i cance (Silberling and Tozer, 1968; Silberling and Nichols, 1982; Baliniet al., 2007)and their taxonomic signif i cance (Roniewicz and Stanley, 1998; Hopkin and McRoberts, 2005; Waller and Stanley, 2005).However, few comprehensive studies of faunal patterns within a stratigraphic framework exist.This research focuses on South Canyon, Central Nevada— a field location that exposes Middle to Upper Triassic rocks (Figure 1).
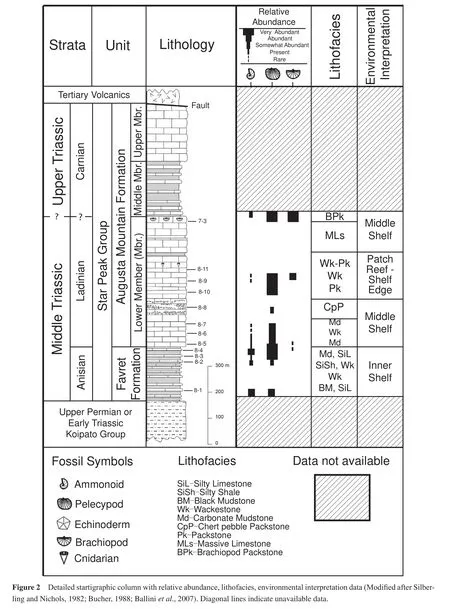
The goal of this project is to document and compare the fossil distributions within their environmental parameters to reveal any underlying associations and determine if or when the switch to more modern characteristics (e.g.,infaunal life style)occurred in this section.Based on preliminary observations, our hypothesis states that within our area, Paleozoic taxa and ecologies (e.g., epifaunal life style)dominate Middle Triassic communities.We test this hypothesis by collecting petrographic and abundance data from the Favret Formation and Lower Member, Augusta Mountain Formation in South Canyon, Nevada (Figure 2).
2 Background
2.1 Geology
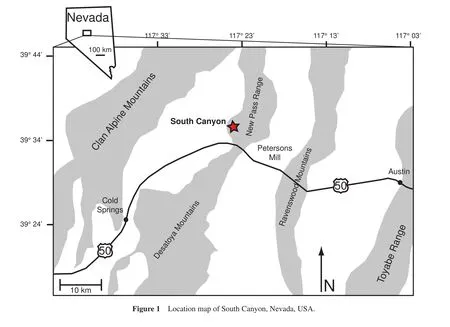
This study focuses on the Triassic section at South Canyon, located in the southern portion of the western front of the New Pass Range, in North-Central Nevada (Baliniet al., 2007; Figure 1).Our research focuses on the Favret Formation and Lower Member of the Augusta Mountain Formation, both within the Star Peak Group (Figure 2).The Star Peak Group consists of units of mostly carbonate marine rocks of Middle to Late Triassic age exposed throughout North-Central Nevada (Emmons, 1877; King,1878; MacMillian, 1972; Nichols and Silberling, 1977).The Star Peak Group overlies the Koipato Group; the Koipato Group consists primarily of conglomerates and silt-stones along with volcanic and volcaniclastic rocks (Baliniet al., 2007)and lacks marine fossils (MacMillian, 1972).The Koipato Group contains abundant scour marks, ripple marks, and cross-bedding, which are indicative of a f l uvialdeltaic system (MacMillian, 1972).The Favret Formation conformably overlies the Koipato Group and it is the basal unit of the Star Peak Group (MacMillian, 1972; Baliniet al., 2007)(Figure 2).The Favret Formation consists of interbedded thin-bedded, dark organic-rich limestones with calcareous shales and siltstones, representing a near-shore environment (MacMillian, 1972).Much of the Favret Formation is comprised of thin-shelled bivalve biosparite(MacMillian, 1972), with a shallow benthic fauna consisting primarily of brachiopods, crinoids and gastropods (Bucher, 1992a).The Favret Formation at South Canyon has been correlated with the Fossil Hill Member of the Favret Formation exposed elsewhere in the New Pass Range (Mc-Coy and Wildhorse mines)and from various localities in North-Central Nevada: in the Augusta Mountains (Favret and Muller Canyons), in the southern Tobin Range, and with the Prida Formation in the Humboldt Range (Bucher,1992b).The lower portion of the Favret Formation is not present at South Canyon as it is in other sites due to subsidence resulting from synsedimentary block faulting following a period of uplift and erosion prior to deposition of the upper portion (MacMillian, 1972).
The Augusta Mountain Formation overlies the Favret Formation and is divided into three members: Lower, Middle, and Upper (Figure 2).The Lower Member consists mostly of thick-bedded to massive limestone, with intercalated shales and siltstones in the lower portion, cherty conglomerates/breccias in the middle portion and massive limestones with corals in areas and a few shale intervals in the upper portion (Baliniet al., 2007).The uppermost portion of the Lower Member consists of brachiopod-rich limestone layers, which have a “coquina-like” appearance(Johnston, 1941; Balini and Jenks, 2007; Baliniet al.,2007)(Figure 2).MacMillian (1972)interpreted the Lower Member as a carbonate shelf environment, with the presence of shales and massive limestones indicating a lower energy, offshore environment relative to the near shore environment of the Favret Formation.However, the presence of the cherty conglomerates/breccias indicates brief regressional periods due to regional uplift that resulted in increased input of terrigenous sediments deposited over the carbonate shelf (MacMillian, 1972).The Middle Member consists of alternating marls and limestones (Baliniet al.,2007)and is indicative of a further offshore environment due to the increased amount of clay (MacMillian, 1972).The Upper Member consists of massive gray limestones(Baliniet al., 2007)and represents a similar depositional environment as the Middle Member (MacMillian, 1972).The Star Peak Group is unconformably overlain by Tertiary ash-f l ow tuffs with volcaniclastic deposits (Baliniet al., 2007; Figure 2).
2.2 Paleontology
Numerous studies have described the ammonoid fauna at South Canyon and have been instrumental in developing North American Triassic biostratigraphy (Johnston, 1941;Silberling, 1956; Silberling and Tozer, 1968; Balini and Jenks, 2007; Baliniet al., 2007).Due to the nearly complete ammonoid succession across the Ladinian-Carnian boundary at South Canyon, attempts have been made to define the Global Boundary Stratotype Section and Point(GSSP)for the base of the Carnian (Waller and Stanley,2005; Balini and Jenks, 2007; Baliniet al., 2007).In addition to ammonoids, the Triassic succession at South Canyon yields several other groups of fossils, including:cnidarians, foraminifera, conodonts, bivalves, brachiopods and gastropods.Several studies have been carried out on the various taxonomic groups within the South Canyon section.These studies include descriptions of the foraminifera (Gazdzicki and Stanley, 1983), cnidarians (Roniewicz and Stanley, 1998), bivalves (Waller and Stanley,2005)and brachiopods (Smith, 1914).The aforementioned studies were mostly reported from the Lower Member of the Augusta Mountain Formation.However, conodonts were also reported from the Middle Member, Augusta Mountain Formation (Mosher, 1968; Baliniet al., 2007).Overall, substantial paleontological studies have been conducted in South Canyon.However, none have conducted a quantitative abundance analysis of taxa and their ecologies within an environmental context.
3 Methods
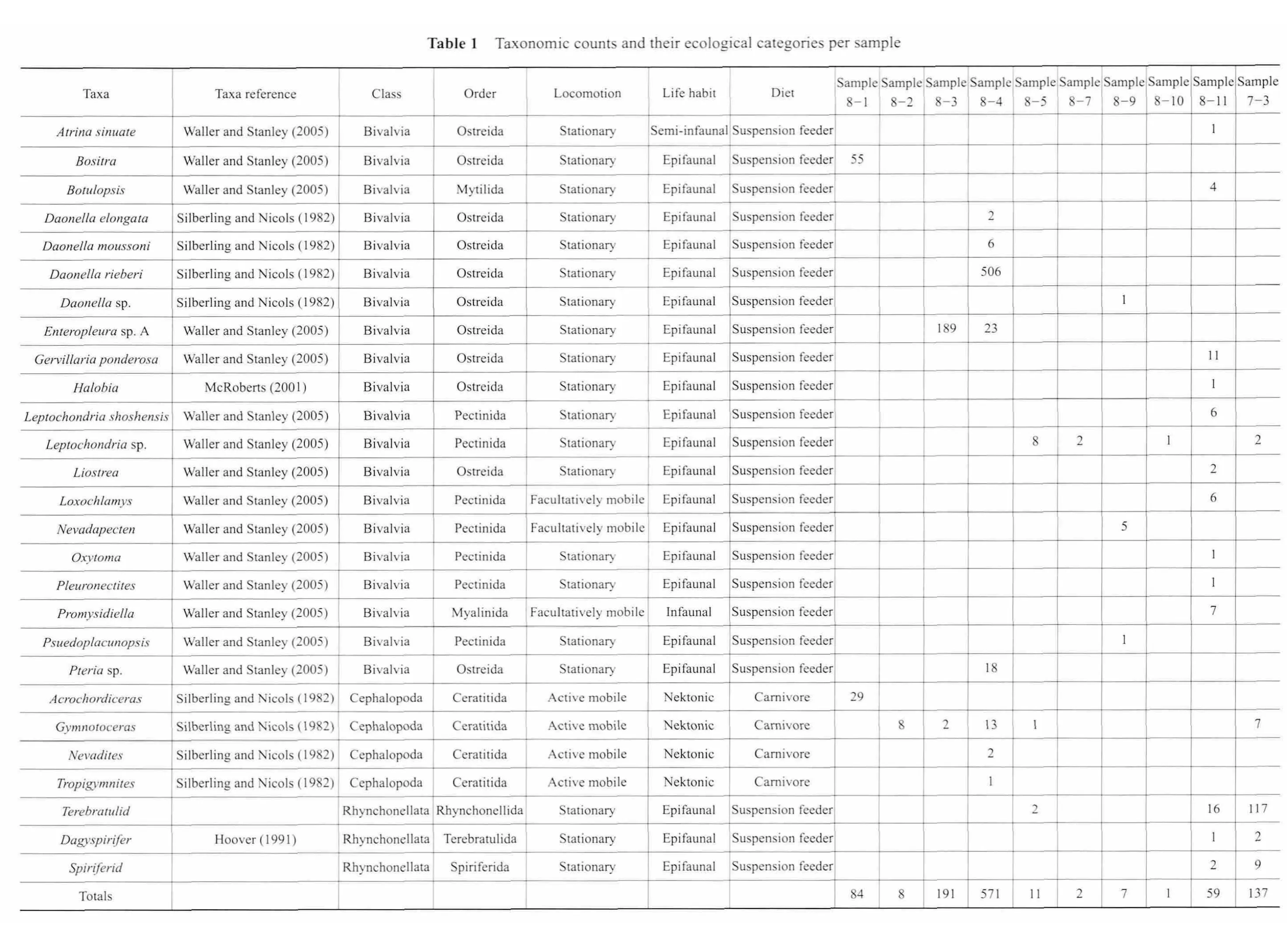
This study aims to document Middle Triassic faunal rediversif i cation, thus samples were collected from the Favret Formation and Lower Member, Augusta Mountain Formation (Figure 2).The formations were measured via compass and Jacob staff method and described and sampled as measured.Samples were collected as we walked the canyon f l oor.Good exposures were limited resulting in a coarse sampling scheme of one sample approximately every 50 meters.Twelve bulk samples were collected and each bulk sample contains three to eight slabs of fossiliferous rock.Visible fossils in hand samples were identif i ed to the genus level and to the species level when it was possible (Figure 3).Abundance data were tallied and ecological categories were determined using the Paleobiology Database (www.pbdb.org; Table 1).Seventy petrographic thinsections were created for sedimentological analyses.The section comprises mixed siliciclastic-carbonates hence we used Dunham’s (1962)carbonate classification scheme to categorize our rocks.
4 Samples and results
The samples collected for the study span the Upper Anisian to Ladinian of the Middle Triassic rocks at South Canyon (Figure 2).The bulk samples consist of primarily carbonate rocks.However, slabs from silt and shale beds were also collected.The following will review the overall sedimentology and fossil content of each sample both in hand sample and thin-section (See Table 1 for complete lists of taxonomic and ecologic data).
4.1 Sample 8-1: Favret Formation
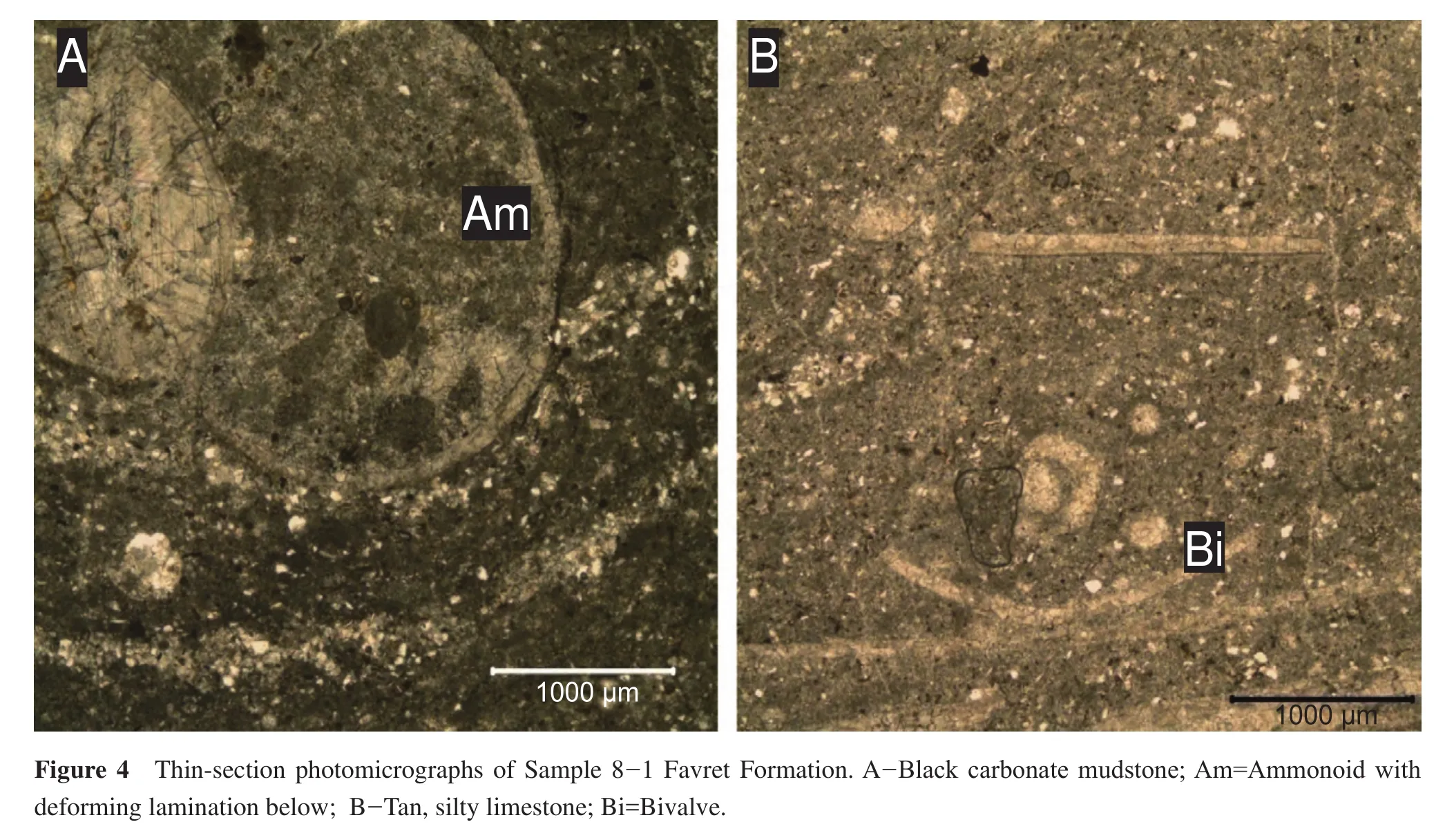
This sample consists of black, organic-rich,carbonate mudstone and tan, laminated silty limestone.Most of the fossils in this sample come from the tan colored silty limestone, which lies parallel to bedding with little or no bioturbation.The fossils in this sample consist of abundant ammonoids and flat-clam bivalves.Thin-section analysis reveals that the black carbonate mudstone has a peloidal mud matrix (Figure 4A).The tan colored rock consists of micrite matrix with high abundance of siliciclastics,which occur in layers (Figure 4B).Evidence of laminations giving way under the weight of an ammonoid suggests that the sediment at the time of deposition consisted of soft mud.The tan rock also contains a large amount of hematite and siliciclastics (Figure 4B).The organicrich content, evidence of soft-sediment deformation and the mudstone grain size indicate deposition took place in a quiet water environment.In terms of ecology, mobile nektonic carnivores dominate the community (Figure 4;Table 1).
4.2 Sample 8-2: Favret Formation

This sample consists of a medium gray wackestone with several ammonoid impressions.In terms of bulk,this sample was the smallest and therefore a small number of fossils were extracted (Table 1).This sample contains mostly ammonoids — mobile nektonic carnivores (Figure 5); bivalve fragments can only be seen in thin-section (Figure 6).As with the previous sample, this sample contains abundant hematite with some siliciclastics.Weak laminations are present, indicative of a quiet water environment(Figure 6).
4.3 Sample 8-3: Favret Formation
This sample consists of tan siltstones and brown silty wackestone with high amounts of hematite.The siltstones are very fossiliferous consisting almost entirely of the Halobiid bivalveEnteropleurasp.A — stationary epifaunal suspension feeders (Figures 3H, 5).The silty wackestone contains wavy brown to orange laminations of predominately hematite and other siliciclastics and is devoid of fossils (Figure 7).The grains within the silty wackestone are coarse silt and sand size (< 1 mm).
4.4 Sample 8-4: Favret Formation
This part of the section is comprised of thin-bedded limestones interbedded with siltstone.The rock consists of a dark gray mudstone with a considerable amount of silt and siliciclastics.AbundantDaonellaspecimens of various species are present throughout — all of which are stationary epifaunal suspension feeders (Figure 5).In addition, ammonoids, foraminifera and ostracodes are present (Figure 8A, 8B).The presence of flat clams such asDaonellaand the overall mudstone grain size suggest a quiet depositional environment.
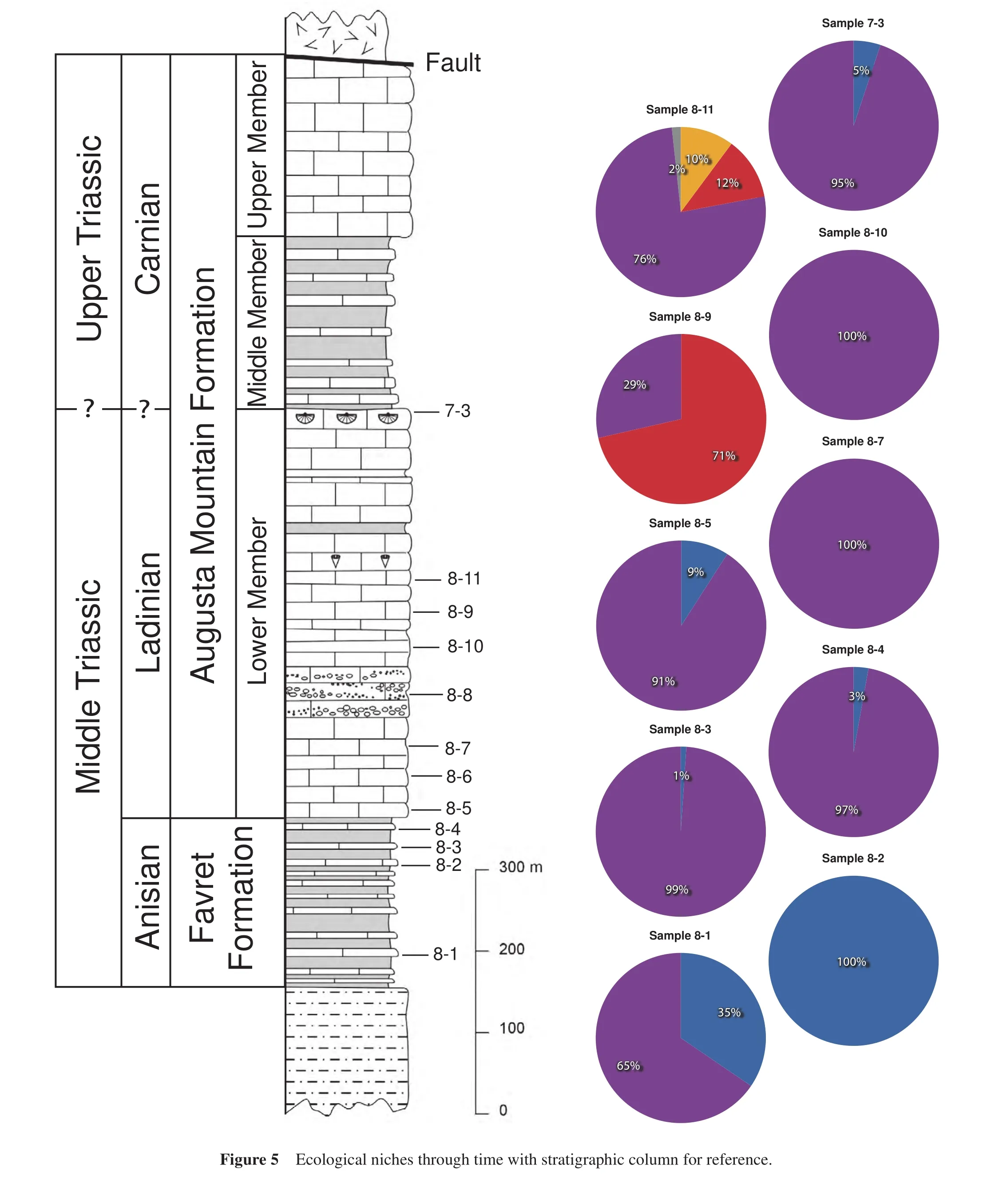
4.5 Sample 8-5: Lower Member, Augusta Mountain Formation
This sample comprises a dark gray mudstone with iron oxide staining.In hand sample, brachiopods and bivalves,both stationary epifaunal suspension feeders, dominate while ammonoids — mobile nektonic carnivores comprise 9% of the counted specimens (Figure 5).An even higher abundance of fossils exists in thin-section including ostracodes, bivalves, ammonoids, gastropods, and possible sponges (Figure 9A, 9B).
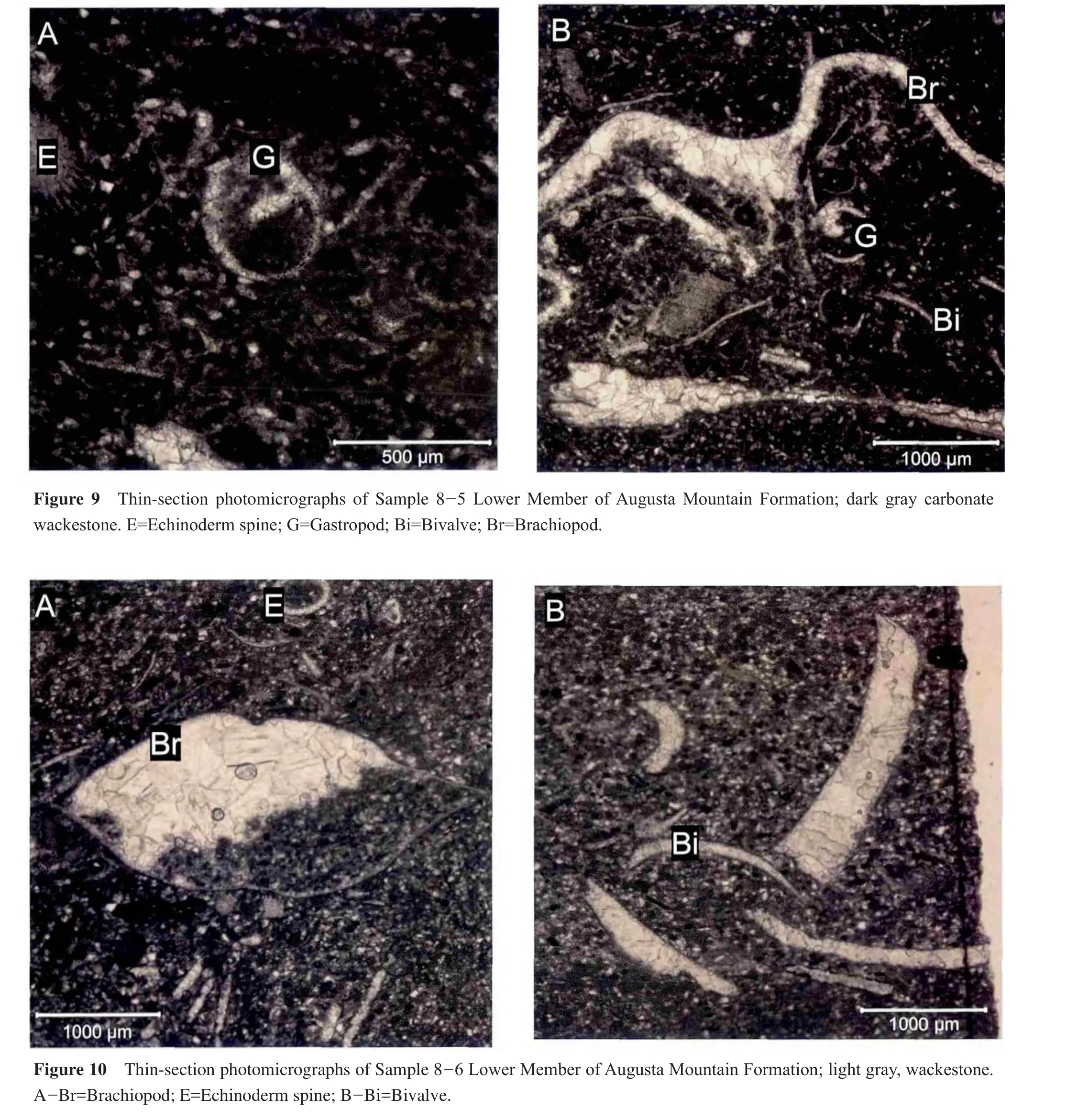
4.6 Sample 8-6: Lower Member, Augusta Mountain Formation
This sample consists of light gray wackestone with large, pinkish iron oxide stains.The exterior of the hand samples lacks fossils; however, thin-sections indicate a high abundance of fossils including brachiopods, echinoderms, foraminifera, and ostracodes (Figure 10A, 10B).
4.7 Sample 8-7: Lower Member, Augusta Mountain Formation
3 7-2 7 1 1 7 2 9 7 1 3 S a m p l e 1 1 8-1 4 1 1 1 6 2 6 1 1 7 1 6 1 2 5 9 S a m p l e l e 8-1 1 S a m p 1 0 l e S a m p 9 8-1 5 1 7 S a m p l e 7 8-2 2 5 8-8 1 2 1 1 4 S a m p l e 8-S a m p l e 2 6 6 5 0 2 3 1 8 1 3 2 1 5 7 1 l e S a m p 3 8-1 8 9 2 1 9 1 l e p l e 8-S a m p 2 8 8 g i c a l c a t e g o r i e s p e r s a m 8-1 5 5 2 9 8 4 S a m p l e D i e t e n s i o n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r n f e e d e r o r e o r e o r e o r e n f e e d e r n f e e d e r s p u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o n f e e d e r u s p e n s i o u s p e n s i o u s p e n s i o u s p e n s i o e n s i o n f e e d e r s p u s p e n s i o u s p e n s i o C a r n i v C a r n i v C a r n i v C a r n i v u s p e n s i o u s p e n s i o u s p e n s i o n f e e d e r S u n t s a n d t h e i r e c o l o i t L i f e h a b E p i f a u n a l S E p i f a u n a l S E p i f a u n a l S E p i f a u n a l S E p i f a u n a l S n a l S E p i f a u n a l S E p i f a u n a l S E p i f a u n a l S E p i f a u i f a u n a l S i c i f a u n a l S i c i c i f a u n a l S i c n a l S i f a u n a l S n a l S E p E p E p E p i f a u n a l S E p i f a u n a l S E p E p i f a u n a l S f a u n a l I n i f a u n a l S E p E p i f a u n a l S N e k t o n N e k t o n N e k t o n N e k t o n E p i f a u E p i f a u i f a u n a l S E p i l e i l e o b S e m i-i n f a u n a l S u o b i l e o b i l e i l e i l e i l e i c c o u m o t i o n n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y n a r y o b o b o b o b n a r y n a r y n a r y a x o n o m L o c o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o S t a t i o l t a t i v e l y m F a c u l t a t i v e l y m F a c u S t a t i o S t a t i o l t a t i v e l y m F a c u S t a t i o S t a t i o A c t i v e m A c t i v e m A c t i v e m A c t i v e m S t a t i o S t a t i o S t a t i o a T a b l e 1 T O r d e r O s t r e i d a O s t r e i d a M y t i l i d a O s t r e i d a O s t r e i d a O s t r e i d a O s t r e i d a O s t r e i d a O s t r e i d a O s t r e i d a a a P e c t i n i d P e c t i n i d O s t r e i d a a P e c t i n i d a P e c t i n i d a P e c t i n i d a P e c t i n i d a a l i n i d a e l l i d a o n M y P e c t i n i d O s t r e i d a C e r a t i t i d a C e r a t i t i d a C e r a t i t i d a C e r a t i t i d a T e r e b r a t u l i d i r i f e r i d a S p R h y n c h B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a B i v a l v i a d a d a d a d a C l a s s h a l o p o o n e l l a t a o n e l l a t a o n e l l a t a e p h a l o p o e p e p h a l o p o e p h a l o p o R h y n c h R h y n c h R h y n c h 5)0 5)0 5)0 5)0 5)c e (2 0 0)T a x a r e f e r e n S t a n l e y (2 0 0 5)t a n l e y (2 0 0 5)t a n l e y (2 0 0 5)i c o l s (1 9 8 2)i c o l s (1 9 8 2)i c o l s (1 9 8 2)i c o l s (1 9 8 2)t a n l e y (2 0 0 5)t a n l e y (2 0 0 5)t a n l e y t a n l e y (2 0 0 5)t a n l e y (2 0 0 5)t a n l e y (2 0 0 5)i c o l s (1 9 8 2)C i c o l s (1 9 8 2)C i c o l s (1 9 8 2)C i c o l s (1 9 8 2)C W a l l e r a n d W a l l e r a n d S W a l l e r a n d S S i l b e r l i n g a n d N S i l b e r l i n g a n d N S i l b e r l i n g a n d N e r l i n g a n d N b e r t s (2 0 0 1)S t a n l e y (2 0 S t a n l e y (2 0 S t a n l e y (2 0 S t a n l e y (2 0 S t a n l e y (2 0 0 5)S t a n l e y (2 0 0 5)e r (1 9 9 1 H o o v S i l b W a l l e r a n d S W a l l e r a n d S M c R o a l l e r a n d S W a l l e r a n d W a l l e r a n d W a l l e r a n d W a l l e r a n d W a l l e r a n d W a l l e r a n d S W a l l e r a n d S W a l l e r a n d W a l l e r a n d S e r l i n g a n d N S i l b e r l i n g a n d N S i l b S i l b e r l i n g a n d N S i l b e r l i n g a n d N s s o n i s i s W s a d e r o A t r i n a s i n u a t e s h e n s i s s T a x a s i t r a e l o n g a t a o u t u l o p s i s r i e b e r i e l l a s p.s p.A r a o n r i a s p.m y s a i e l l a n i t e s i r i f e r B o n d o t o c e r a s d i t e s B o e l l a d a p e c t e n r o n e c t i t e s y s i d D a o n D a o n e l l a m e l l a o n r i a p H a l o b i a n d r i a s h o c h o r d i c e r a T o t a l s D a D a o n t e r o p l e u L i o s t r e a O x y t o m P t e r i a s p.N e v a S p i r i f e r i d E n G e r v i l l a L e p t o c h o L e p t o c h o L o x o c h l a N e v a P l e u P r o m P s u e d o p l a c u n o p A c r o G y m n T r o p i g y m T e r e b r a t u l i d D a g y s p

These rocks consist of dark gray mudstone suggestive of an organic-rich environment.Very little fossils are seen on the exterior of the hand sample but a higher abundance is seen in thin-section including: echinoderms, bivalves,and ostracodes (Figure 11).Worth noting is the overall reduction of siliciclastics in comparison to previous samples.This might suggest that the depositional environment was more offshore.Only two bivalve specimens were identifi ed for the entire sample thus ecological interpretations are unreliable (Figure 5).
4.8 Sample 8-8: Lower Member, Augusta Mountain Formation
This sample represents the onset of a cherty, conglomerate/breccia.The sample consists of red chert clasts in brown, silty, packstone.The chert pebbles are poorly sorted and range from subrounded to subangular.There are no fossil specimens visible in hand samples, however several echinoderm and bivalve fragments are visible in thin-section (Figure 12).
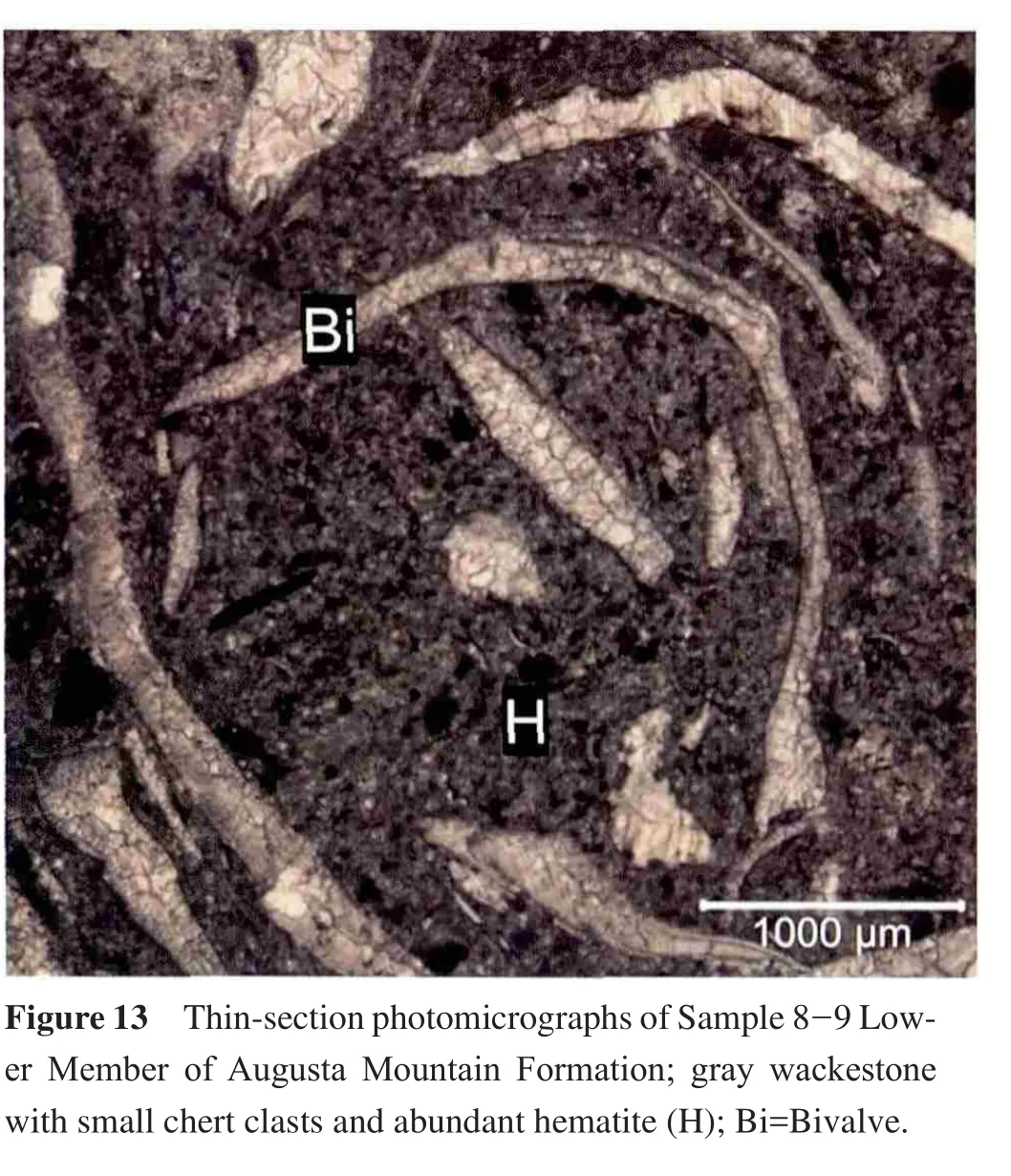
4.9 Sample 8-9: Lower Member, Augusta Mountain Formation
This sample consists of gray wackestone with small chert clasts and abundant hematite crystals.Stationary epifaunal suspension feeding bivalves dominate this sample(Figure 5; Table 1).Similar to previous samples, the majority of the fossils are seen in thin-section.These fossils include bivalves, ammonoids, and ostracodes (Figure 13).
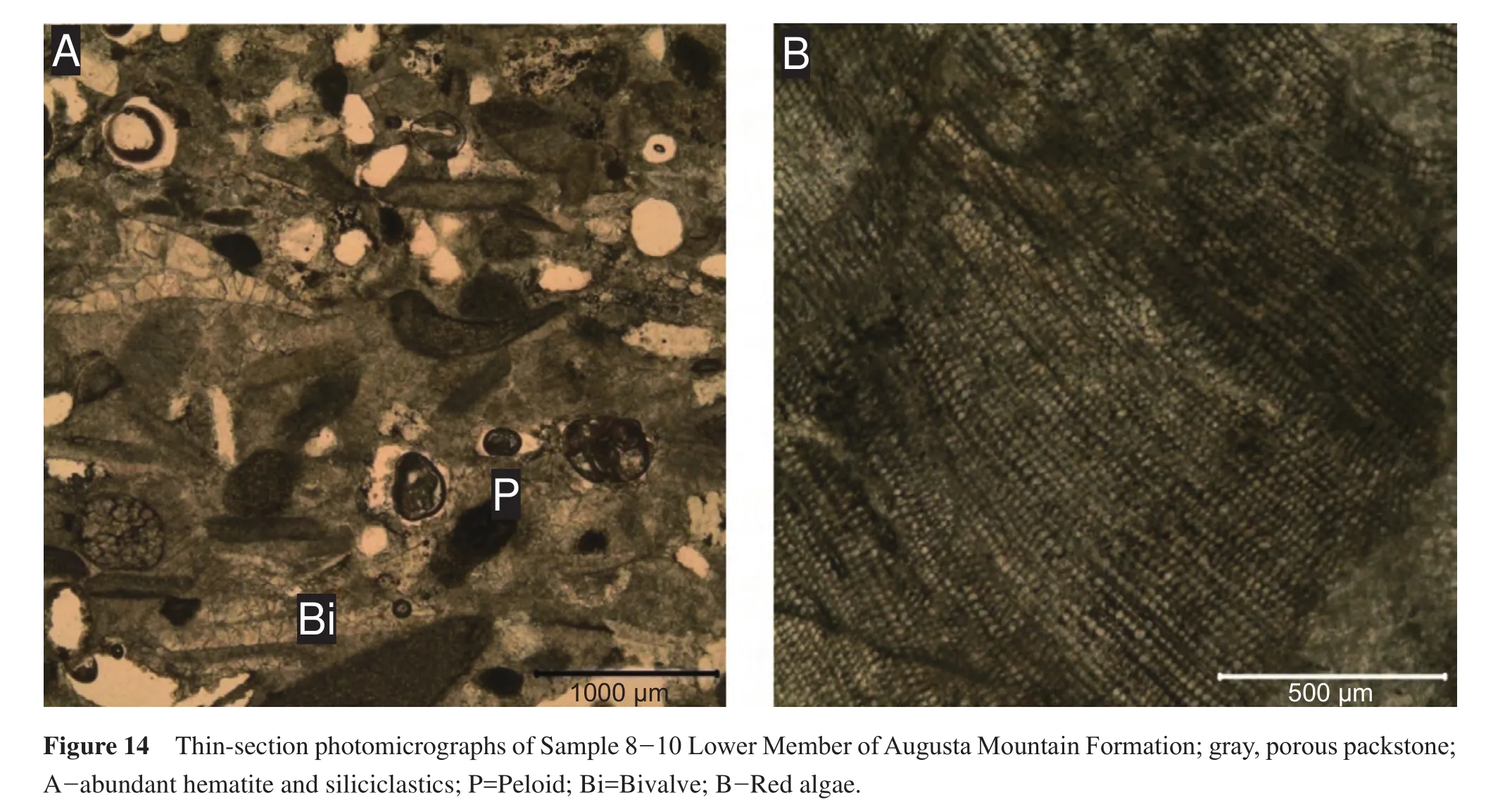
4.10 Sample 8-10: Lower Member, Augusta Mountain Formation

This sample consists of a gray, porous packstone, which is composed entirely of chert at the base.The only fossils visible on the exterior of the sample are fragments of red algae and one stationary epifaunal suspension-feeding bivalve (Figure 5; Table 1).In thin-section, algae are present as well as echinoderm fragments, and few bivalves (Figure 14).The non-chert portions of this sample are comprised of abundant, poorly sorted siliciclastics, with abundant hematite and peloids; these observations in combination with the presence of red algae suggest a high-energy depositional environment.
4.11 Sample 8-11: Lower Member, Augusta Mountain Formation
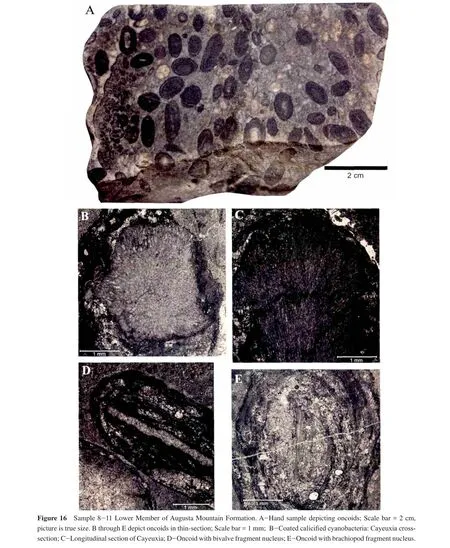
This sample is the most fossiliferous sample and the most ecologically diverse.It is composed of a dark wackestone with abundant Pteriomorphian bivalves, ammonoids,brachiopods, echinoderms, algae fragments and well-preserved crinoid ossicles and echinoderm spines (Figures 15,16; Table 1).Although not abundant, it is important to note the appearance ofGervillaria(Baryvellia)ponderosa— a large, thick-shelled, bivalve.Stationary epifaunal suspension feeder organisms comprise 76% of the same, while 11% are facultatively mobile infaunal suspension feeders,10% are facultatively mobile epifaunal suspension feeders, 2% are facultatively mobile deep infaunal chemosymbiotic, and 2% are stationary, semi-infaunal suspension feeders (Figure 5).In thin-section, we see oncoids with algal envelopes around fossil fragments and abundant siliciclastics (Figures 15, 16).This sample was collected below well-known coral beds (Gazdzicki and Stanley, 1983).Based on its close stratigraphic position to the coral beds and the presence of coated grains, we interpret a moderate to high-energy patch reef environment.
4.12 Sample 7-3: Lower Member, Augusta Mountain Formation
A dark gray, wackestone to packstone with signif i cant iron oxide staining comprises this sample (Figure 17).This sample consists of abundant terebratulid and spiriferid brachiopods and rare bivalves and ammonoids with the sample consisting of 94% stationary epifaunal suspension feeders (Figure 5).
5 Environmental interpretations
Based on our preliminary sedimentological and paleontological data we suggest that during the Middle Triassic an accretionary rimmed shelf margin existed in this area— beginning with the quiet, inner shore environment that eventually transitions to shelf edge and patch reefs environments with time (Figure 2).
Beginning with the base of the Favret Formation we record evidence of a silty limestone with abundant hematite and siliciclastics, which we interpret as a near-shore, low to moderate energy environment, most likely an intertidal zone — near-coast zone between normal high and low-tide levels (Flügel, 2004).MacMillan (1972)noted the presence of ripple-marks in this portion of the section, which is in agreement with our interpretation.The community represented by this sample is dominated by nektonic, carnivorous ammonoids, which constitute 85% of the counted specimens with the other 15% made up of bivalves.The lack of sessile specimens is likely due to sampling bias since the total number of specimens counted equals thirtyfour.The lack of sessile organisms could also be explained by low oxygen conditions, which can result in a black mudstone like the ones observed in the lower part of the section.


As the Favret Formation continued to be deposited,siliciclastics f l owed into the system and resulted in the periodic deposition of siltstones interbedded within the predominant carbonate system.Evidence of ammonoid specimens depressing lamination adds to this interpretation of a muddy, quiet environment, further supporting the inference of an inner shelf depositional setting.Through time we see evidence of a higher abundance of siliciclastics, which could indicate a subtle drop in sea level change or an increase in local tectonics.
The Augusta Mountain Formation marks a lithological change from carbonate mudstone interbedded with siltstone to more massive carbonate wackestones and packstones.Overall we see a progression of decreased terrigenous siliciclastics in the system compared to the Favret Formation — except where chert pebble carbonates exist.This shift towards wackestone and packstones, including the abundance of echinoderm fragments, peloids, oncoids and algal envelopes around fossil fragments, as well as an increase in abundance of bivalves, brachiopods, gastropods, foraminifera and ostracodes, and lastly the reported presence of reef-building fossils (i.e., corals), lead to the interpretation of a deeper intertidal to shallow subtidal,mid- to high-energy environment.We suggest that as sea level rose progressively, the environment changed to a middle shelf environment, eventually patch reefs formed on the shelf edge and returned back to a middle shelf environment again (Figure 2).This overall transgression was interrupted by intervals of regional uplift introducing chert into the system resulting in the chert pebble limestone conglomerate/breccia seen in Sample 8-8.MacMillan (1972)reported a similar trend of gradual transgression with cycles of regression due to regional uplift within the Ladinian portion of the South Canyon section.
6 Taxonomic and ecological changes through time
The Early Mesozoic is thought to record the switch from brachiopod-dominated benthic communities to gastropod-bivalve dominated benthic communities (McRoberts, 2001).In addition to this major reorganization of benthic communities, researchers document an increase in infaunalization, shell thickness and intricate shell ornamentation (McRoberts, 2001).These changes are generally thought to have been a product of increased predatory pressures that forced benthic communities to take part in an evolutionary “arms race” with predators (Vermeij,1977).However, McRoberts (2001)gathered data from the Middle Triassic of Europe and concluded that during the Triassic, evidence for elevated predation was lacking,particularly drill holes created by predatory gastropods.McRoberts (2001)suggests that differential environmental stresses between epifaunal and infaunal habitats, coupled with high competition, may have given the opportunity for infaunal organisms to take over.He also suggests that infaunalization may have been initially coordinated with the end-Permian extinction with the elimination of the epifaunal communities later in the Triassic.
Taxonomic and ecologic patterns within our data indicate that benthic communities begin as ammonoid and flat,epifaunal bivalve dominated (within the Favret Formation)and progress to a benthic community still dominated by epifaunal organisms but infaunalization begins to increase with time.These observed ecological changes can result from the lithologic changes between the Favret Formation and Lower Member of Augusta Mountain Formation; however, when we focus on ecological patterns just within the Lower Member, Augusta Mountain Formation— a unit interpreted as being deposited along the middle shelf and shelf edge, we observe similar trends.The Lower Member of Augusta Mountain Formation begins with epifaunal bivalves dominating followed by an increase in infaunalization and shell thickness up-section (as observed by the presence ofGervillaria)(Figure 3; Table 1).Thus, more modern ecologies begin to occur in the middle of the Ladinian; that is, toward the end of the Middle Triassic.The data from this study is also consistent with a gradual increase in the richness of bivalves throughout the Middle Triassic (McRoberts, 2001).
Further up-section we observe a taxonomic change while the general epifaunal dominant ecology remains the same — epifaunal bivalves give way to epifaunal brachiopods.The overall lack of observed predators (e.g., carnivorous ammonoids and gastropods)up-section suggests that this taxonomic change did not occur due to predatory stresses.Instead, we believe these taxonomic changes might be a result of changes in overall substrate; that is, a change from muddy, soft bottom substrate to a f i rmer substrate.However, further investigation must be completed to conf i rm this hypothesis.
Overall, our section records an interesting time when benthic communities are in, what we would like to call“faunal limbo”; they are neither fully Paleozoic nor fully modernized.Instead, we see modern taxa (i.e., bivalves)with Paleozoic ecologies (i.e., epifaunal suspension feeders)dominating the Middle Triassic with glimpse of a more fully modernized community beginning to take hold in the middle Ladinian stage only to be replaced by a typical Paleozoic Fauna assemblage — that is, dominated by epifaunal brachiopods.As mentioned previously, further investigation needs to be completed in order to fully document this transition.Respectively, a more detailed faunal and sedimentological sampling scheme, including recording evidence of substrate changes, needs to occur to confi rm the overall patterns observed in our study.
7 Conclusions
We present new taxonomic and ecologic data within an environmental context, from the Middle Triassic within the eastern Panthalassa realm.Beginning in the Anisian and continuing through the Ladinian, we observe a transgressional event over a rimmed carbonate platform that periodically experienced local uplift.Within this carbonate platform, we record faunal changes beginning in the Anisian with an ammonoid dominated to flat-clam, epifaunal dominated benthic community that progressively increased in ecological complexity via the introduction of infaunalization within the middle Ladinian stage.By the end of the Ladinian, epifaunal brachiopods dominate benthic communities.We attribute these taxonomic and ecologic changes to changes in substrate rather than predator/prey interactions because of the lack of observed predators through time; however, further investigation is needed to conf i rm our substrate hypothesis.And, overall we report that in our section, Paleozoic-style communities (i.e., epifaunal bivalves and brachiopods)dominate with the introduction of modern-style communities occurring in the middle Ladinian.
Acknowledgements
This research was supported by grants from California State University to Dr.Bonuso.We would like to thank Dr.Nathaniel J.Lorentz for thoughtful comments and overall fi eld support, and Dr.Frank Corsetti for support with microfacies analyses.
Balini, M., Jenks, J.F., 2007.The Trachyceratidae from South Canyon (Central Nevada): Record, taxonomic problems and stratigraphic signif i cance for the definition of the Ladinian-Carnian boundary.In: Lucas, S.G., Spielmann, J.A., (eds).The Global Triassic.New Mexico of Natural History and Science Bulletin,41: 14-22.
Balini, M., Jenks, J.F., McRoberts, C.A., Orchard, M.J., 2007.The Ladinian-Carnian boundary succession at South Canyon (New Pass Range, Central Nevada).In: Lucas, S.G., Spielmann, J.A.,(eds).Triassic of the American West.New Mexico of Natural History and Science Bulletin, 40: 127-138.
Bonuso, N., Bottjer, D.J., 2008.Testing biogeographic, environmen-tal, and taxonomic effects on abundance patterns of Triassic brachiopods and bivalves.Palaios, 23: 43-54.
Bucher, H., 1988.A new Middle Anisian (Middle Triassic)ammonoid zone from northwestern Nevada (USA).Eclogae Geologicae Helvetiae, 81: 723-762.
Bucher, H., 1992a.Ammonoids of the Hyatti Zone (Middle Anisian,Middle Triassic)and the Anisian transgression in the Star Peak Group (northwestern Nevada).Paleontographica Abt.A, 223:137-166.
Bucher, H., 1992b.Ammonoids of the Shoshonensis Zone (Middle Anisian, Middle Triassic)from northwestern Nevada (USA).Palaeontographica A (Paleozoology, Stratigraphy).
Chen, J., Chen, Z.Q., Tong, J.N., 2010.Palaeoecology and taphonomy of two brachiopod shell beds from the Anisian (Middle Triassic)of Guizhou, southwest China: Recovery of benthic communities from the end-Permian mass extinction.Global and Planetary Change, 73: 149-160.
Diedrich, C., 2009.Vertebrates of the Anisian/Ladinian boundary(Middle Triassic)from Bissendorf (NW Germany)and their contribution to the anatomy, palaeoecology, and palaeobiogeography of the Germanic Basin reptiles.Palaeogeography, Palaeoclimatology, Palaeoecology, 273: 1-16.
Dunham, R.J., 1962.classification of carbonate rocks according to depositional texture.AAPG Mem., 1: 108-121.
Emmons, S.F., 1877.Desatoya Mountains.U.S.Geological Exploration.40thParallel Report, 2: 644-649.
Feng, Q., Zhang, Z., Ye, M., 2001.Middle Triassic radiolarian fauna from southwest Yunnan, China.Micropaleontology, 47: 173-204.
Flügel, E., 2004.Microfacies of carbonate racks: Analysis, interpretation and application.Springer: Berlin-Germany, 976.
Gazdzicki, A., Stanley, G.D., 1983.First report of Involutinidae (Foraminifera)in marine Triassic rocks of North America.Jahrbuch für Geologie und Paläontologie Monatshefte, 2: 80-90.
Hoover, P.R., 1991.Late Triassic cyrtinoid spiriferinacean brachiopods from western North America and their biostratigraphic and biogeographic implications.Bulletins of America Paleontology,100: 63-109.
Hopkin, E.K., McRoberts, C.A., 2005.Middle Triassic flat clams(Pterioida: Halobiidae)from the Middle Triassic Anisian of North-Central Nevada.Jounral of Paleontology, 79: 796-800.
Johnston, F.N., 1941.Trias at New Pass, Nevada (New Lower Karnic ammonoids).Journal of Paleontology, 15: 447-491.
King, C., 1878.Systematic Geology.U.S.Geological Exploration.40thParallel Report, 1-803.
Komatsu, T., Jinhua, C., Meizhen, C., Stiller, F., Naruse, H., 2004.Middle Triassic (Anisian)diversif i ed bivalves; depositional environments and bivalve assemblages in the Leidapo Member of the Qingyan Formation, southern China.Palaeogeography, Palaeoclimatology, Palaeoecology, 208: 207-223.
MacMillian, J.R., 1972.Late Paleozoic and Mesozoic tectonic events in West Central Nevada.Chicago, IL, Northwestern University.
McRoberts, C.A., 2001.Triassic bivalves and the initial marine Mesozoic revolution: A role for predators? Geology, 29: 359-362.
Miller, A.I., Sepkoski, J.J.J., 1988.Modeling bivalve diversif i cation: The effect of interaction on a macroevolutionary system.Paleobiology, 14: 364-369.
Mosher, L.C., 1968.Triassic conodonts from western North America and Europe and their correlation.Journal of Paleontology, 42:895-946.
Nichols, K.M., Silberling, N.J., 1977.Stratigraphy and depositional history of the Star Peak Group (Triassic), Northwestern Nevada.Geological Society of America Special Paper, 178: 1-73.
Pálfy, J., 1990.Paleoecological signif i cance of Anisian (Middle Triassic)brachiopod assemblages from the Balaton Highland, Hungary.In: MacKinnon, D.I., Lee, D.E., Campbell, J.D., (eds).Second Annual International Brachiopod Congress.Rotterdam,A.A.Balkema, 2: 241-246.
Payne, J.L., Lehrmann, D.J., Christensen, S., Wei, J., Knoll, A.H.,2006.Environmental and biological controls on the initiaion and growth of a Middle Triassic (Anisian)reef complex on the great bank of Guizhou, Guizhou Province, China.Palaios, 21: 325-343.
Posenato, R., 2008.Patterns of bivalve biodiversity from Early to Middle Triassic in the Southern Alps (Italy): Regional vs.global events.Palaeogeography, Palaeoclimatology, Palaeoecology,261: 145-159.
Roniewicz, E., Stanley, G.D.Jr., 1998.Middle Triassic cnidarians from the New Pass Range, central Nevada.Journal of Paleontology, 72: 246-256.
Sepkoski, J.J.Jr., 2002.A compendium of fossil marine animal genera.Bulletins of American Paleontology, 363: 1-560.
Silberling, N.J., 1956.“Trachyceras Zone” in the Upper Triassic of the Western United States.Journal of Paleontology, 30: 1147-1153.
Silberling, N.J., Nichols, K.M., 1982.Middle Triassic molluscan fossils of biostratigraphic signif i cance from the Humboldt Range,northwestern Nevada.Prof.Pap.U.S.geol.Surv., 1207: 1-77.
Silberling, N.J., Tozer, E.T., 1968.Biostratigraphic classification of marine Triassic in North America.Geological Society of America Special Paper, 110: 1-63.
Smith, J.P., 1914.The Middle Triassic marine Invertebrate Faunas of North America.United States Geological Survey, Professional Paper, 83: 1-254.
Vermeij, G.J., 1977.The Mesozoic marine revolution: Evidence from snails, predators, and grazers.Paleobiology, 3: 245-258.
Waller, T.R., Stanley, J.G.D., 2005.Middle Triassic pteriomorphian bivalvia (Mollusca)from the New Pass Range, West-Central Nevada: Systematics, biostratigraphy, paleoecology, and paleobiogeography.Journal of Paleontology, 79: 1-58.
 Journal of Palaeogeography2014年1期
Journal of Palaeogeography2014年1期
- Journal of Palaeogeography的其它文章
- Congratulations on success of the 1st International Palaeogeography Conference
- 19th International Sedimentological Congress 2014 in Geneva: Theme 12—Palaeogeography
- Quantitative lithofacies palaeogeography
- Diversity patterns and palaeobiogeographical relationships of latest Devonian-Lower Carboniferous foraminifers from South China: What is global,what is local?
- The ichnofacies and ichnoassemblages in terrestrial deposits of China
- Foraminiferal biostratigraphy of lignite mines of Kutch, India: Age of lignite and fossil vertebrates