
放牧胁迫下冰草种群遗传多样性的研究
2014-09-20 06:47赵萌莉参木友
草原与草业 2014年1期
秀 花,赵萌莉,韩 冰,参木友,拉 巴
(1.西藏农牧科学院草业科学研究所,西藏 拉萨市 850009; 2.内蒙古农业大学生态环境学院,内蒙古 呼和浩特市 010019)
放牧胁迫下冰草种群遗传多样性的研究
秀 花1,赵萌莉2,韩 冰2,参木友1,拉 巴1
(1.西藏农牧科学院草业科学研究所,西藏 拉萨市 850009; 2.内蒙古农业大学生态环境学院,内蒙古 呼和浩特市 010019)
本研究采用ISSR分子标记技术对放牧胁迫下冰草的遗传多样性进行了研究。由Shannon’s和Nei’多样性指数检测的研究结果表明,虽然放牧胁迫导致部分位点丢失,但整个种群仍表现出丰富的多态性,ISSR检测的多态性条带比率为91.95%,种群间的遗传差异很小,放牧胁迫并未使冰草种群产生遗传分化。
冰草;ISSR;放牧胁迫;遗传多样性
引言
冰草属(Agrop J.Gaertn)为禾本科早熟禾亚科大麦族小麦亚族植物。是一种放牧和打草兼用型牧草,并已广泛驯化栽培作为草地补播草种。有关冰草遗传多样性方面的研究报道很多。阎贵兴〔1〕(2001)发现冰草属植物不同种间存在核型多型性;云锦凤〔2〕(1990)对蒙古冰草B染色体进行了分析;谢新民〔3〕(2001)对蒙古冰草遗传多样性研究结果表明,蒙古冰草种内居群间及同一居群内的不同个体间存在丰富的多样性;李景欣〔4〕(2004)利用根尖压片法,从细胞水平对来自6个不同居群的冰草的遗传多样性进行研究发现,冰草种内居群间存在一定程度的分化和变异,一些北美国家有关冰草的研究,主要侧重其生态建设、耐牧性及营养物质的运输和储存等。
本文应用ISSR分析技术对不同放牧梯度下的冰草遗传多样性及其分化进行研究,并为探索合理的放牧制度和草地利用方式,防止草原退化和退化草原的恢复提供理论基础,为筛选耐牧基因提供遗传学信息。
1 材料和方法
在植物生长旺盛期按株采集冰草新鲜叶片。株距50m以上,选择株丛径中等、生长正常的植株30株,剪去叶片,用保鲜膜包好放入冰盒带回实验室,贮存于-20℃备用。
1.1实验地自然概况
本实验选用的冰草取自内蒙古锡林郭勒盟白音锡勒牧场中国科学院草地生态定位站,地处东经:116°04′~117°05′,北纬:43°26′~44°08′,海拔高度:1250-1280m,属温带草原区典型草原栗钙土亚区。年均气温-0.4℃,1月平均气温为-23℃,7月平均气温为17.9℃,年降水量350mm,主要集中在6-8月,占年降水量的76.26%。在研究区域内,选择长期无放牧(围封20年以上)和放牧样地,三次重复。
1.2 DNA的提取与检测
称取0.25g新鲜叶片。采用CTAB法提取每个植株的总DNA,用0.7%的琼脂糖电泳检测DNA质量,如图1、图2为冰草的部分样品DNA电泳结果,DNA的纯度和浓度通过紫外吸收法测定。
1.3 ISSR反应条件及程序
PCR反应体系的建立主要通过对DNA浓度、引物等条件进行初步的调整,达到了反应条件的优化,确定了ISSR扩增反应总体积为25ul,反应体系:其中包括10*buffer 2.5ul,2.0mmol/LMgcl22ul,0.1mmol/LdNTPs 0.4ul,0.15mmol/LISSR引物1ul,40ng左右的基因组DNA,1单位TapDNA聚合酶。以上各种加完后用灭菌三蒸水补齐至25ul。
扩增在PE9600型PCR仪上进行。扩增程序:94℃预变性5min,94℃变性45s,54℃退火1min,72℃延伸1.5min,进行44个循环,最后72℃补平5min,4℃停止。扩增产物用2.8%琼脂糖凝胶(内含溴化乙锭)电泳鉴定,电泳1.5-2.0小时后,将凝胶放在自动凝胶成像系统下进行分析。
1.4引物筛选
以无放牧样地的冰草DNA为材料,对30个ISSR引物进行筛选,筛选出12个扩增条带较多、信号强、背景清晰的引物用于ISSR-PCR反应。引物由上海生物工程公司合成,名称和序列见表1。

表1 ISSR所用引物序列与序号
1.5扩增产物和差异带的分析方法
1.5.1 带的记录
电泳图的每一条带(DNA片段),均为一个分子标记(Marker),代表一个引物的结合位点。根据各分子标记的迁移率及其有无统计所有的二元数据;有带(显性)记作1,无带(隐性)记作0,强带和弱带的赋值均为1。
1.5.2 数据的统计分析法
应用POPGEN32软件对冰草种间遗传结构进行分析,计算Nei基因多样性指数(H),Shannon表型信息指数(I),群体分化系数(Gst)等;根据Nei遗传距离用类平均聚类方法(UPGMA)进行系统聚类,分析群体间遗传分化关系。
2 结果与分析
2.1遗传多态性分析
本实验中,PCR反应扩增出的DNA片断大小在200-1500bp之间,各引物检测出的位点总数和单态位点及多态位点不同(表2),其中22893引物检测出的位点数最多,为12个,36542引物检测出位点数最少,为4个。12个ISSR引物在22份材料间共扩增出87个位点,平均每条引物产生7.25条带,在87个位点中有80个为多态位点,多态位点比率为91.95%。不同引物多态位点从1-12个,多态位点比率从50%到100%。其中无放牧冰草种群多态位点比率为77.01%,放牧冰草种群为60.92%(表3)。
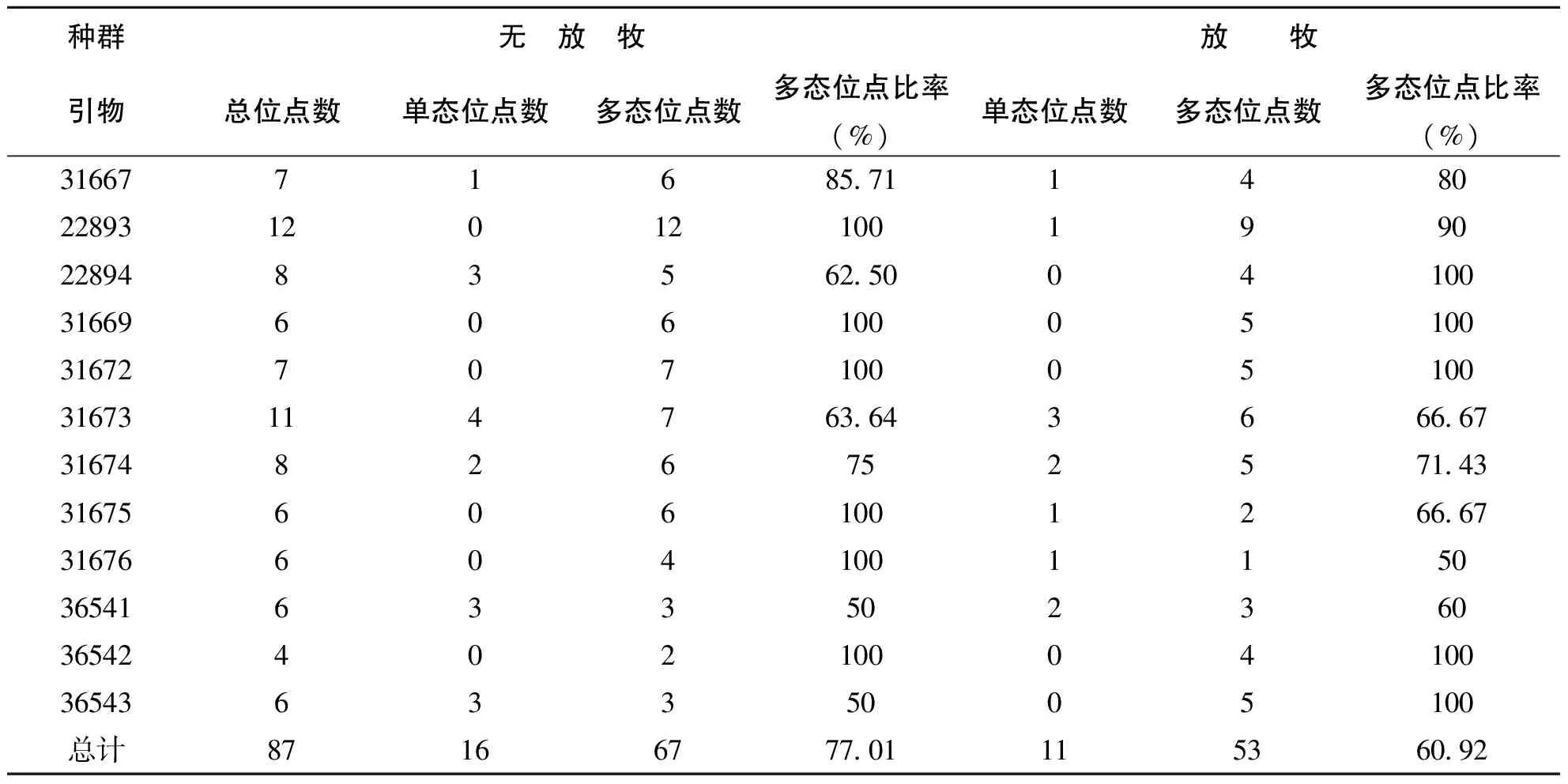
表2 各引物扩增的位点数及多态位点数
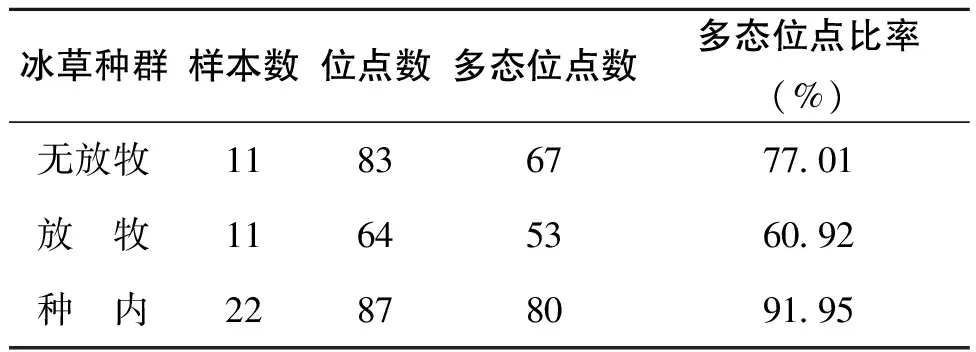
表3 冰草种群的多态位点比较
2.2基因频率
用ISSR分析无放牧与放牧冰草种群的基因频率变化,经G2检验,在引物31673扩增的1200bp位点,引物22893扩增的1000bp、1500bp,引物22894的450bp、550bp、750bp,引物31675的550bp,引物31676的520bp,引物31672的900bp,引物36542的900bp,引物36543的200bp、350bp位点上两个种群的基因频率变化表现出极显著性(p<0.001),在引物22894扩增的1000bp位点、引物31676的420bp、引物31669的700bp、引物31672的1000bp、引物36542的900bp、引物36543的800bp位点均为显著(p<0.01)。在引物22894扩增的450bp、550bp位点,引物36543的200bp位点上无放牧种群基因频率均为1,放牧种群基因频率均为0。而引物31676扩增的520bp位点上无放牧种群的基因频率为0,放牧种群的基因频率为1。可见放牧胁迫不仅使冰草种群的基因频率发生了变化,与无放牧种群相比,放牧种群部分位点缺失,但也有的位点只在放牧种群中出现。
2.3遗传多样性分析
根据表4,引物31669估算的种群内遗传多样性(Hpop)最高(0.53),引物31676估算的种群内遗传多样性最小(0.21),12个引物估算的种群内遗传多样性平均值是0.33;对种内总的遗传多样性(Hsp)的估算,引物31669估算数值最高(0.58),引物36541估算数值最低(0.30),平均值为0.44。由Shannon,s指数估计的种群内遗传多样性在引物31669中最大为91.18%,引物22894最小44.92%,种群内遗传多样性的平均值为75.87%。种群间遗传多样性幅度从8.82%(引物31669)到55.08%(引物22894),种群间遗传分化的平均值为24.13%,说明放牧胁迫对冰草种群的分化程度有影响,由于2个冰草种群所处的地理自然环境相同,种群之间并未产生隔离,仍存在基因交流,因而没有哪个种群拥有特有的位点或等位基因。但是,并非所有等位基因在不同种群均有分布,有的位点在不同的种群上有丢失现象。
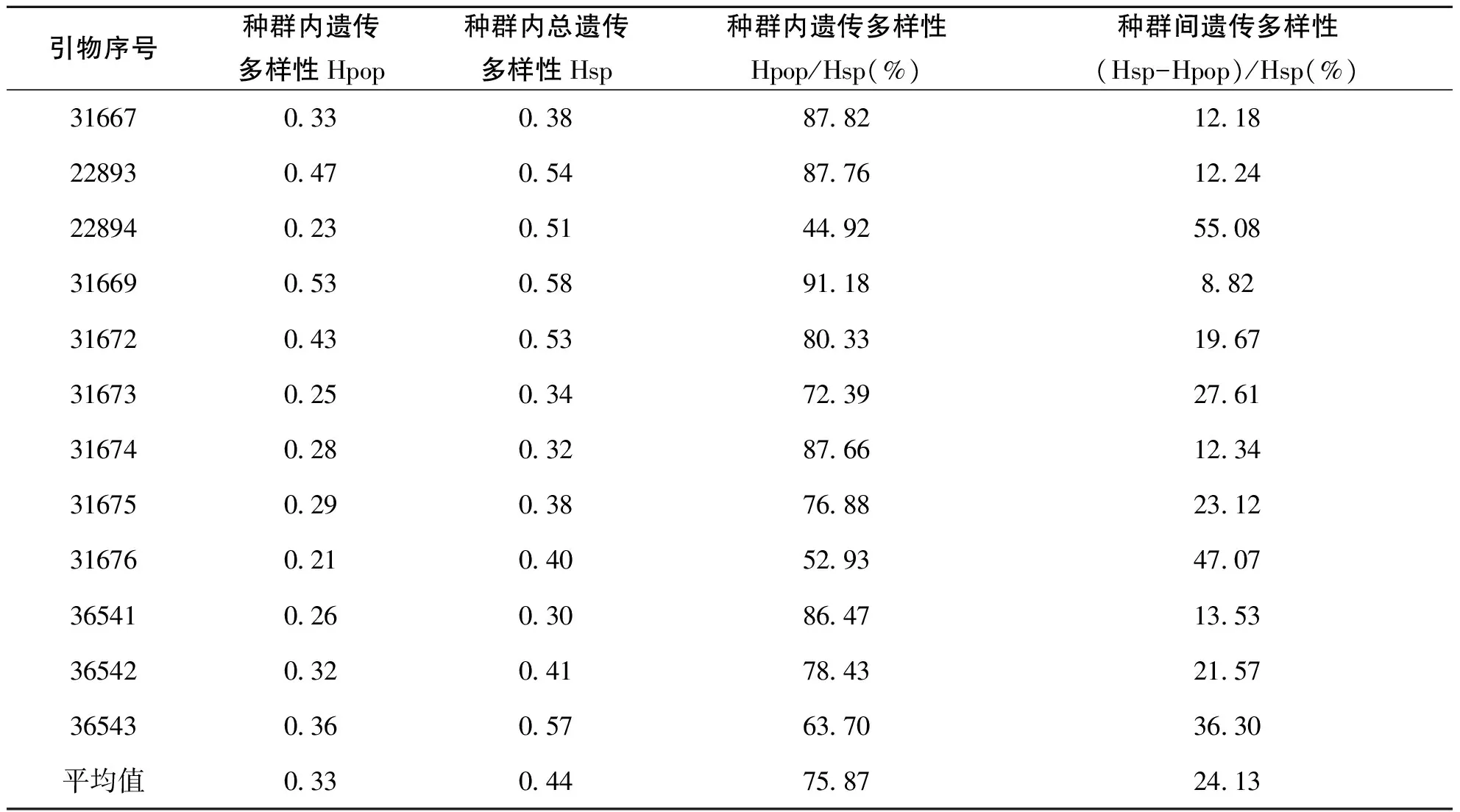
表4 Shannon’s多样性指数估计的冰草种群内、种群间遗传多样性及分化
2.4遗传结构与基因流
Nei(表8)’指数计算的冰草种群内遗传多样性平均值是0.22,种内总遗传多样性的平均值为0.29,种群间的遗传分化为23.66%,说明有23.66%的遗传变异存在于种群间,大部分遗传变异存在于种群内(76.34%),但不同引物所占的比率不同,其中引物22894最高为57.62%,而引物31669最低仅为9.55%。
表5中,各个引物的Nm值从4.74(引物31669)到0.37(引物22894)不等,平均值为1.61。Wrigst(1951)认为〔5〕,当Nm>1时,基因流可以防止遗传漂变引起的居群间的遗传分化,如果Nm<1则由于遗传漂变可以导致种群间明显的遗传分化。因此,当Nm值生高时,Gst值就降低。无放牧与放牧冰草种群的基因流的存在说明种群间有基因的交流,且遗传分化不是由遗传漂变产生的,通过对冰草种群生长环境的分析研究,冰草种群相似的基因频率可能是由于相同的生境选择压力所引起的。
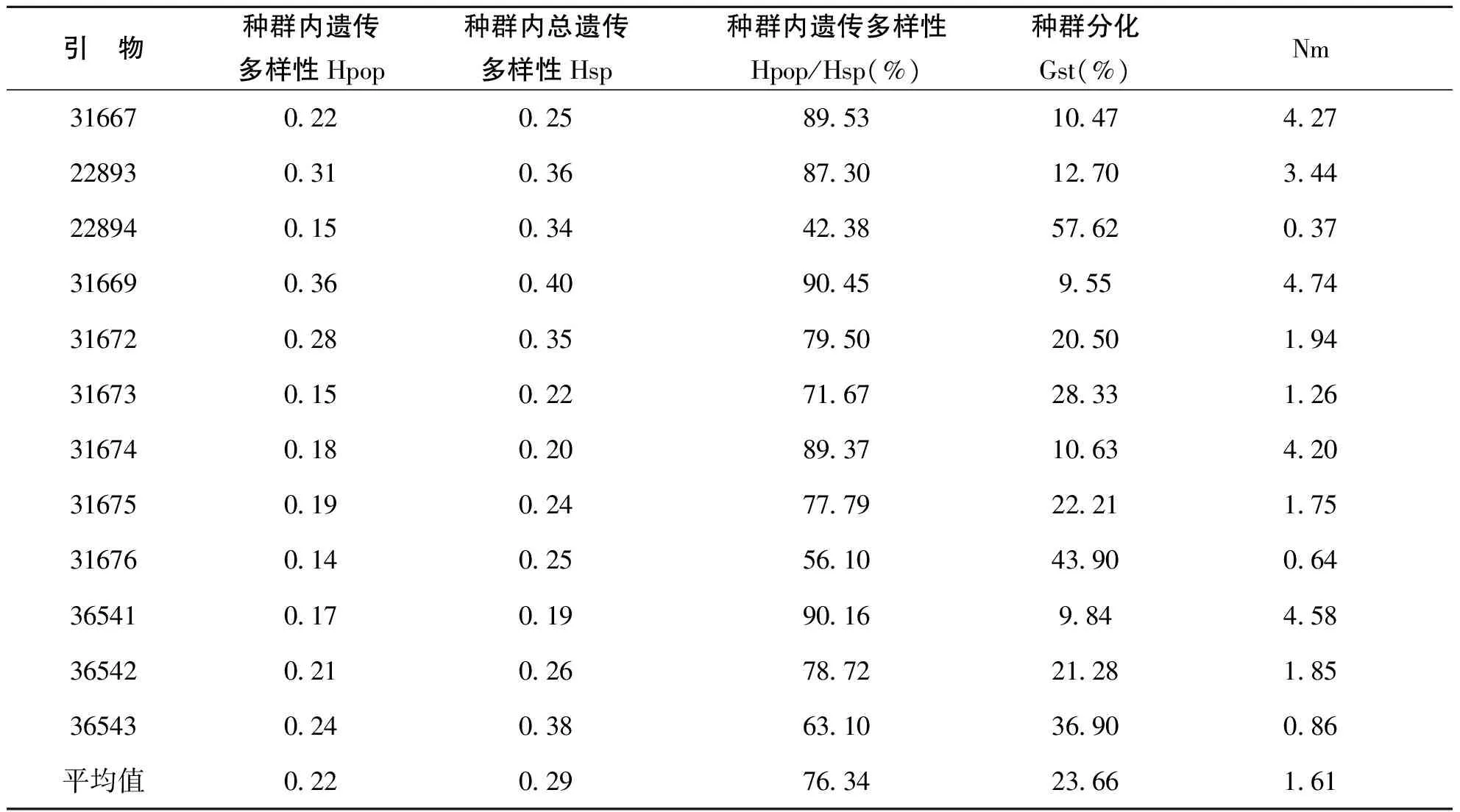
表5 Nei,指数估计的冰草种群内、种群间遗传多样性及分化
Nm=estimate of gene flow Gst or Gcs. E. g., Nm=0.5(1-Gst)/Gst
2.5遗传距离与遗传相似度分析
遗传距离和遗传相似度是评价种群内和种群间遗传变异水平的重要指标。根据12个引物ISSR扩增产物的电泳结果,按Nei的方法计算出无放牧与放牧冰草种群的遗传距离和遗传相似度。遗传距离越大,亲缘关系越远;遗传相似度越大,亲缘关系越近。无放牧冰草种群和放牧冰草种群遗传距离为0.18;遗传相似度为0.83。
3 讨论与结论
本研究对无放牧冰草种群与放牧冰草种群进行ISSR分析的结果,由Shannon’s多样性指数估计的各个引物在2个冰草种群内的遗传多样性显示无放牧冰草种群大于放牧冰草种群,这与Nei’指数估计的遗传多样性结果一致,这说明放牧胁迫在一定程度上降低了冰草种群的遗传多样性。2个冰草种群总的种内多态位点百分率为91.95%,高于高等植物平均遗传多样性水平为70%〔6〕。Grant和Millar等都认为一个物种的进化潜力和抵御不良环境的能力取决于遗传多样性水平的高低〔7、8〕。因此,冰草如此高的遗传多样性水平是其在放牧胁迫等不良环境条件下仍能够生存的遗传基础。在放牧胁迫下,冰草种群ISSR扩增的位点数下降,多态位点数也由无牧的67个下降到放牧的53个,同时随着多态位点数的下降遗传多样性也明显降低。
对无放牧冰草种群和放牧冰草种群进行ISSR分析的结果,放牧胁迫下冰草种群的部分位点丢失,但也有新的位点出现。由Shannon’s和Nei’多样性指数检测的遗传多样性,无放牧冰草种群大于放牧冰草种群,而且无放牧冰草种群的多态位点比率也大于放牧冰草种群,这说明放牧胁迫使冰草种群的遗传多样性降低,但无放牧与放牧冰草种群多态位点比率分别为77.01%和60.92%,种内总多态位点比率为91.95%,可见基因组仍表现出丰富的多态性。
〔1〕阎贵兴.中国草地饲用植物染色体研究.呼和浩特:内蒙古人民出版社,2001
〔2〕云锦凤,斯琴高娃.蒙古冰草B染色体的研究.内蒙古农牧学院学报,1996,17(1):14-17
〔3〕解新民.蒙古冰草的遗传多样性研究.呼和浩特:内蒙古农业大学博士学位论文,2001
〔4〕李景欣,云锦凤,阿拉坦苏布道.冰草的遗传多样性研究.中国草地,2004(6):12-15
〔5〕Wright,s..Genetic structure of population. Ann Eugen,1951,324-354
〔6〕Zhou Y P, Ge S, Wang X D. Molecular marker in systematic and evolutional Botany. Beijing: Science Press, 2001.43-149
〔7〕Grant V. T he Evolutionary Process: A Critical Study of Evolutionary Theory. New York: Columbia University Press, 1991.55-160
〔8〕Millar C I, W J Libby. Strategies for conserving clinical, ecotype, and disjunct population diversity in wide spread species, In: Falk D A ,K E Ho lsinger eds. Genetics and conservation of rare plants. New York: Oxford University Press, 1991: 149-170.
〔9〕NeiM. 分子群体遗传学与进化论.王家玉. 北京: 农业出版社,1983.116-242
〔10〕刘友良.植物水分逆境生理〔M〕.北京:农业出版社,1992.56-118
〔11〕魏伟,王洪新,胡志昂等.毛乌素沙地柠条群体分子生态学初步研究RAPD 证据〔J〕.生态学报,1999,19(1):16-22.
〔12〕赵利峰,李珊,潘莹等.华山新麦草自然居群沿海拔梯度的遗传分化〔J〕.西北植物学报,2001,21(3):391-400.
附图
冰草种群ISSR扩增图版
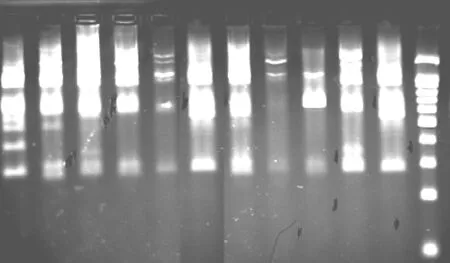
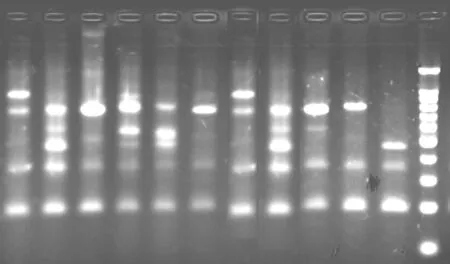
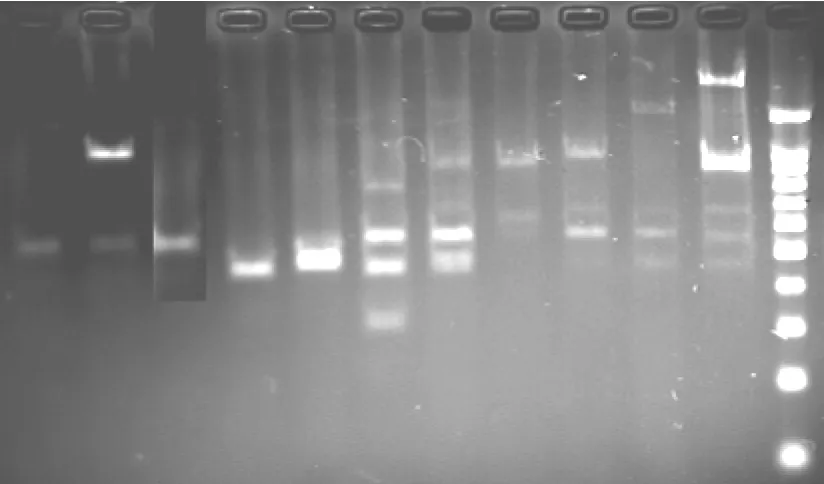
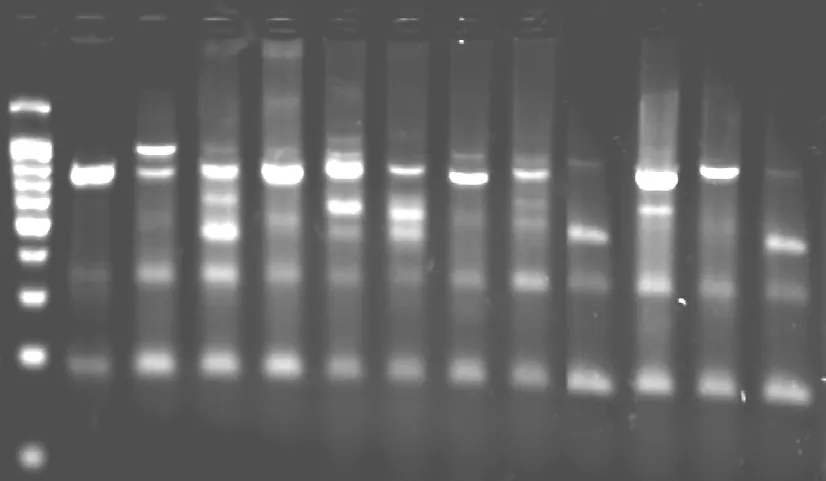
S543+.9
A
2095—5952(2014)01—0030—06
2013-10-22
秀 花(1977-),女,蒙古族,助理研究员,从事草地生态和管理研究工作。
猜你喜欢
今日农业(2022年15期)2022-09-20
新世纪智能(高一语文)(2020年12期)2020-06-01
食品与生活(2019年11期)2019-05-15
红土地(2018年7期)2018-09-26
草原与草业(2018年4期)2018-03-20
科技视界(2016年27期)2017-03-14
中国工程咨询(2016年1期)2016-02-14
食品与健康(2015年8期)2015-08-11
中国医药导报(2015年27期)2015-02-28
当代畜禽养殖业(2014年10期)2014-02-27
