
冀北山地杨桦次生林不同林层的空间结构特征1)
2014-09-18 11:11张曼
东北林业大学学报 2014年9期
张 曼
(河北农业大学,保定,071000)
高兴九
(河北省丰宁满族自治县国营林场)
孙海静 崔丽红 李永宁 黄选瑞
(河北农业大学)
林分空间结构是指林木在林地上的分布格局及其属性在空间上的排列方式。它决定了树木之间的竞争态势、空间生态位,同时,对林分的稳定性、发展的可能性和经营空间的大小起决定作用[1]。通过研究林分空间结构,可为森林调整和制定经营决策提供理论依据[2]。杨桦次生林作为冀北山地的主要森林类型之一,目前,由于林分结构不合理,森林功能难以充分发挥。因此,调整和改造现有林分结构,人工促进其向健康稳定的群落演替,是次生林经营亟待解决的问题。虽然前人对群落结构进行了研究[3-6],但有关林分不同林层空间结构的研究较少。本研究用以相邻木关系为基础,从林木个体水平分布、林木大小分化、空间隔离程度、林木所处的光环境及结构单元的距离尺度等方面加以量化,分别用角尺度、大小比数、混交度和开阔比数表达林分不同林层的空间结构特征,旨在加深对杨桦次生林群落的科学认识,为该地区次生林的保护和经营管理提供理论参考。
1 研究区概况
研究区设立在河北省木兰围场国有林场管理局管辖的桃山林场。桃山林场位于围场县西部,总经营面积1.58×104hm2,属于燕山山脉余脉。地理坐标为 E116°32'~118°14',N41°35'~42°40'。平均海拔在1250 m,属于大陆性季风型高原山地气候,冬春寒冷干燥多风,夏季炎热多雨。无霜期110 d,年平均气温3℃,年均降水量450 mm,年蒸发量1350 mm。土壤多为砂壤土和黑色棕壤土。杨桦次生林是该地区主要的森林类型。主要乔木树种有:白桦(Betula platyphylla)、山杨(Populus davidiana)、色木槭(Acer mono)、棘皮桦(Betula dahurica)、蒙古栎(Quercus mongolica)、稠李(Padus racemosa)等。
2 研究方法
2.1 样地调查
2012年7月,选择典型的杨桦次生林进行实地调查,布设200 m×50 m的样地,样地海拔为1362 m,坡向为南,坡位为下坡,坡度16°,林分郁闭度0.9。样地调查采用相邻网格法,将样地划分成100个10 m×10 m的小样方,再以每个小样方为调查单元,记录每个样方内所有木本植物的树种名称、坐标、树高(H)、胸径(D)、冠幅等。本文中林层的分级标准是:更新层(胸径小于2.5 cm,树高小于2 m);乔木层(胸径大于2.5 cm),乔木层根据树高分为3个林层:低林层(2 m≤H<12 m);中林层(12 m≤H<16 m);高林层(H≥16 m)。
2.2 数据处理
运用Winklemass、Excel软件分别计算样地内更新层和乔木层的空间结构参数(大小比数、角尺度、混交度及开阔比数),运用Origin8.5科技绘图软件绘图辅助分析林分的空间结构特征。为了消除边缘木的影响,特设置了5 m的缓冲区,实际计算面积为7600 m2。
混交度(Mi)。混交度用于描述混交林中林木之间相互隔离程度的指数,定义为参照树i的4株最近相邻木j中与参照木不属同种的个体所占的比例[1]。

大小比数(Ui)。大小比数用于描述树种或单株林木的生长状态,定义为比参照树i大的相邻木j占4株最近相邻木的比例,可用胸径、树高或冠幅来测度[7]。

角尺度(Wi)。角尺度用来描述相邻木围绕参照木的均匀性,定义为在参照树周围,2个最近相邻木的夹角小于标准角(72°)的个数占所考察的最近相邻木的比例[8-9]。

开阔比数(OPi)。开阔比数用于描述林分空间结构中高大乔木的光环境,定义为空间结构单元中参照树不受相邻木遮蔽的程度[10]。

3 结果与分析
3.1 杨桦次生林树种组成
由表1可知,杨桦次生林更新层包括4个树种,乔木层包括6个树种。乔木层中,按株数居前3位的树种依次是白桦、色木槭和山杨;按断面积居前3位的树种依次是白桦、山杨和棘皮桦。乔木层中白桦和山杨的平均高度最大,分别为 13.5、12.5 m,主要原因在于白桦和山杨作为天然林破坏后的先锋树种,生长优势明显。由此可知,白桦和山杨是乔木层的优势种,也是建群种,主要伴生树种有色木槭和棘皮桦,其他伴生树种所占株数比例较小。更新层中,由于色木槭具有耐荫性,所以其更新良好,分布范围广,株数最多。从断面积比例构成看,该林分的树种组成式为7白桦2山杨1棘皮桦+色木槭-蒙古栎-稠李。样地中林木的分布情况(见图1)。
3.2 混交度
由表2可知,更新层的平均混交度为0.228,零度混交的比例高达56.2%,说明更新层中有较多的同种林木聚集在一起,混交程度低;乔木层的平均混交度为0.520,总体上处于中度混交的水平,分布在中度混交水平以上的频率为64.6%,混交状况良好。乔木层中低林层的平均混交度为0.547,混交程度较好;中林层的平均混交度为0.512,的平均混交度为0.410,混交状况不良。混交度随林层增加降低,说明该混交林处于不稳定的演替阶段,林分结构不稳定。
由于树种在林分中的更新方式、混交比例和优势度不同,其混交度存在差异,进而导致不同林层的混交度存在差异[11]。由表2结合图2(a)可知,更新层中色木槭的株数最多,且多呈簇状聚集生长,其零度混交比例高达68.9%,树种混交度仅为0.145,混交状况差。乔木层中,树种株数比例较大的前3个树种依次是白桦、色木槭和山杨,由于其在不同林层中所占株数比例不同,故其混交度不同。低林层共6个树种,除白桦外,其他树种混交度均在中度混交水平以上,混交状况良好。中林层共4个树种,除色木槭(1株)处于完全混交状态外,其他树种处于中、强度混交水平。中白桦株数较多,其零度和弱度混交比例达54%,混交状况不良。
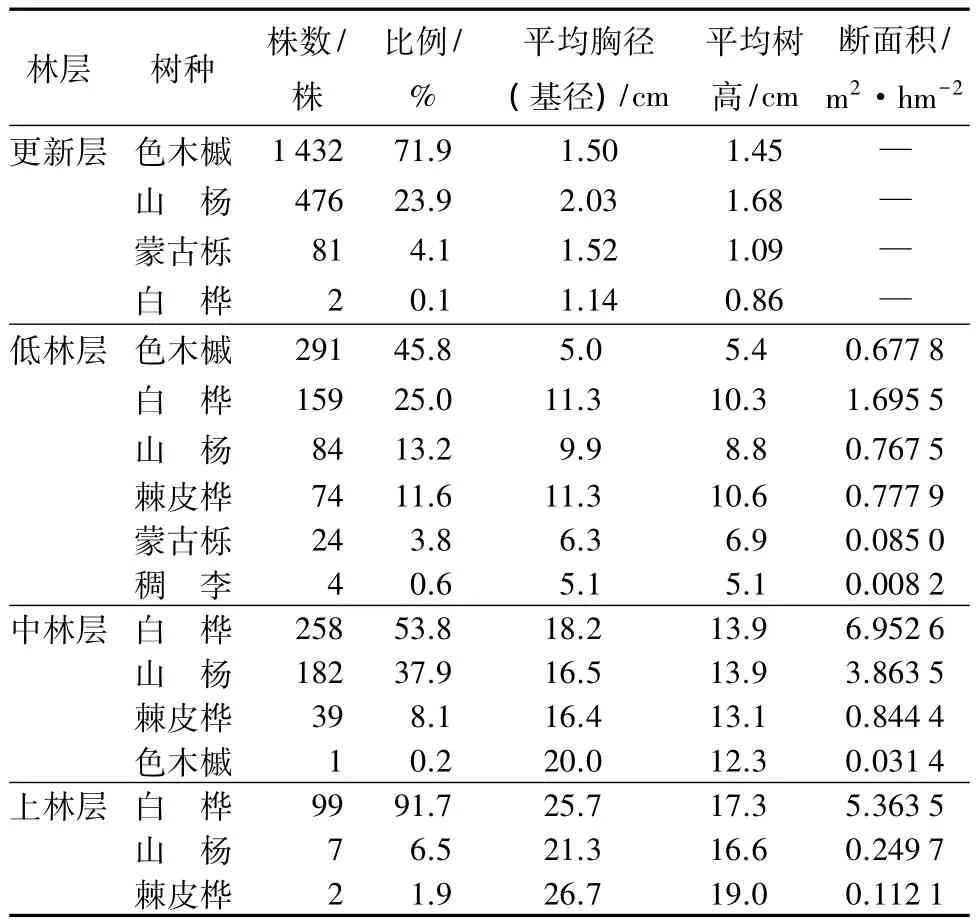
表1 杨桦次生林林分结构特征
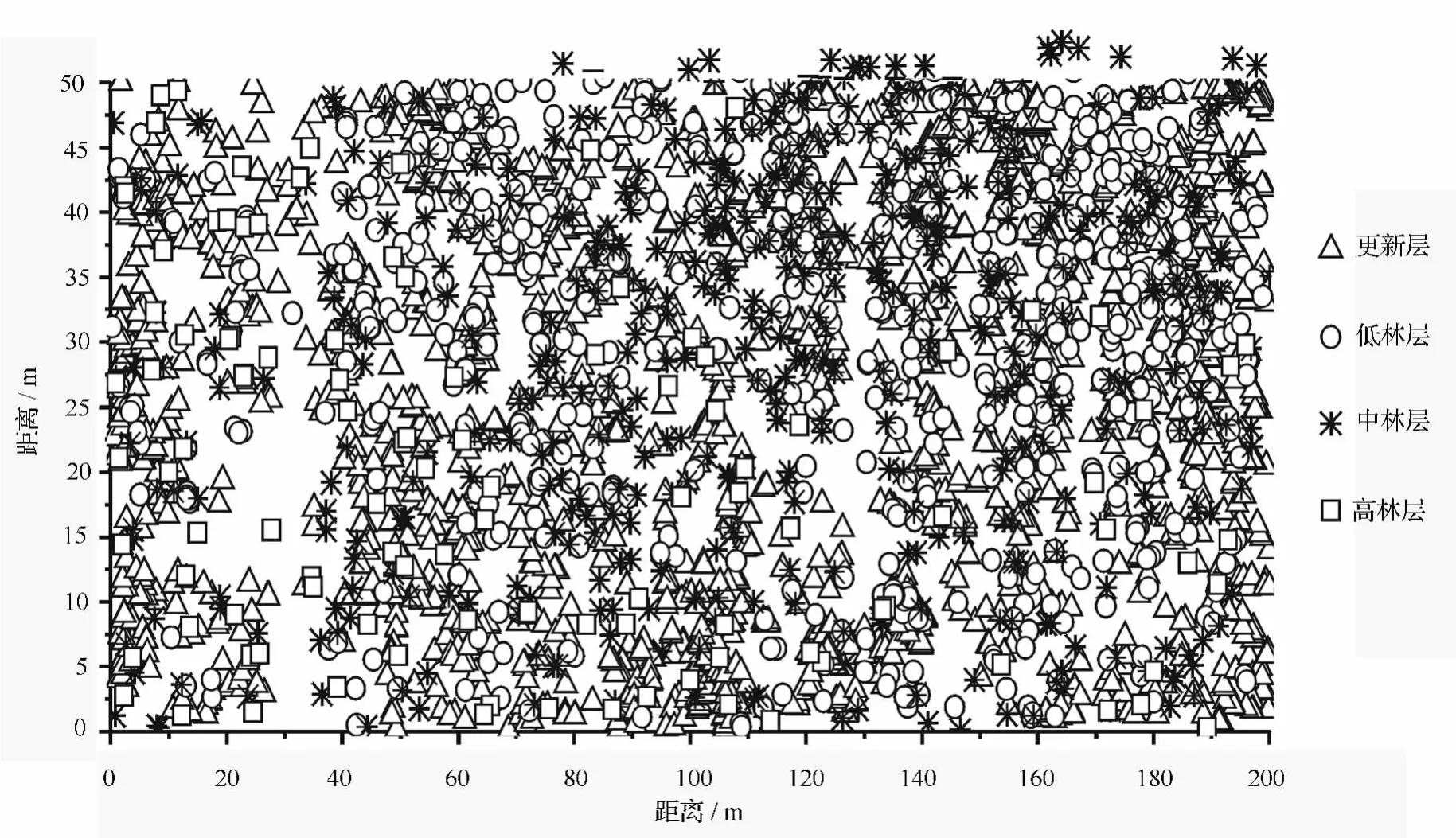
图1 不同林层的林木在样地中的分布
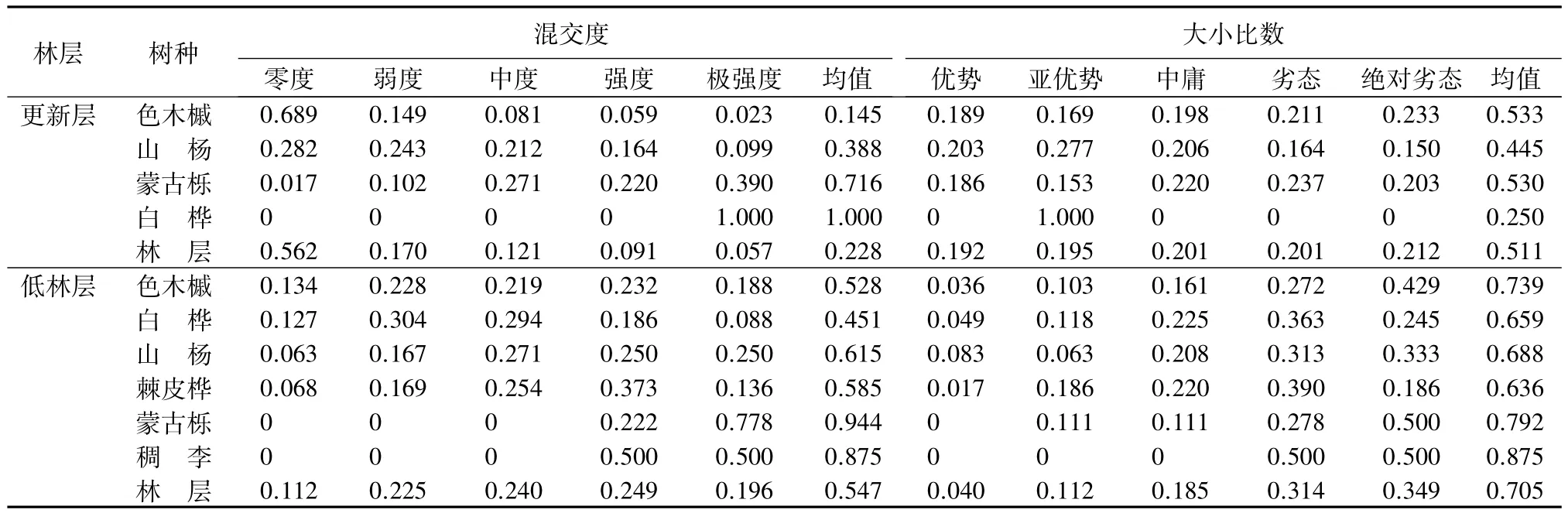
表2 各林层的混交度和大小比数统计
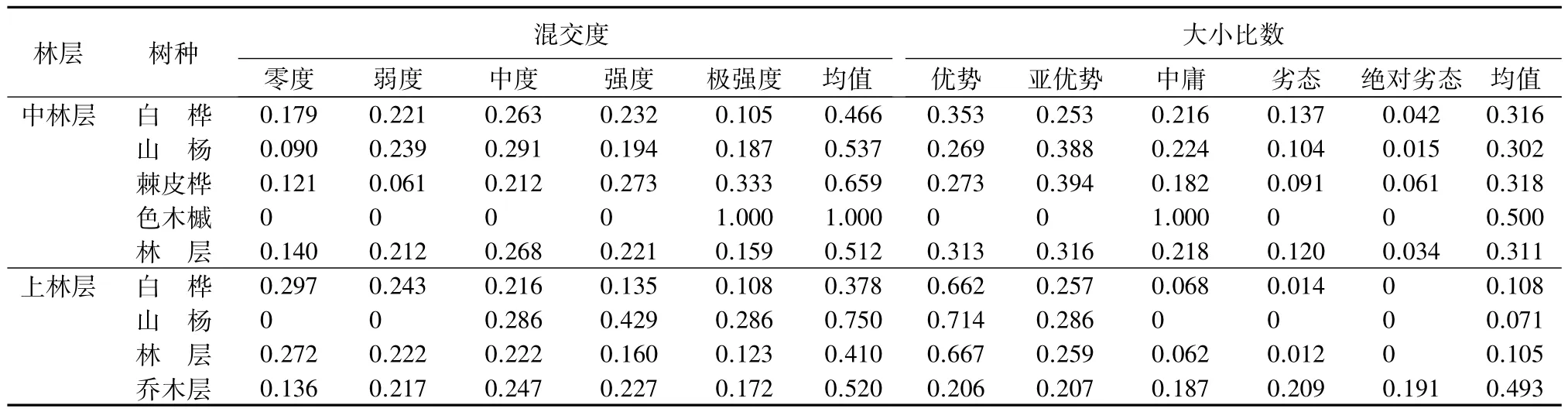
续(表2)
3.3 角尺度
由表3可知,更新层中绝对均匀和均匀分布的结构单元比例较小,分别为0.2%和13.4%,随机分布的结构单元比例最大,为55.5%,不均匀和团状分布结构单元比例为30.8%。更新层平均角尺度为0.575,属于聚集分布,从图2也可看出,更新层密度大,且多呈簇状生长,由于山杨、蒙古栎和白桦的喜光特性,其更新苗大都分布在林木分布较稀疏的地方。乔木层中,随机分布的结构单元比例最大,高达58.3%,绝对均匀和均匀分布的结构单元比例为18.3%,不均匀和团状分布的结构单元比例为23.5%。乔木层的平均角尺度为0.530,属于聚集分布,但聚集程度不大。从乔木层各林层的平均角尺度看,低林层的平均角尺度最大为0.537,其次为中林层和,趋近于随机分布。从图2可看出,白桦种群在低林层,特别是在中林层分布较为密集,这与白桦在幼年阶段为了存活或发挥群体效应常呈簇状聚集生长的特性有关,成林后林木聚集程度由于自然稀疏或人为干扰会有所下降,但林分空间格局总体上仍呈聚集型分布[12]。由于山杨也是喜光树种,所以其多分布在白桦分布较稀疏的地方,色木槭由于耐荫性,分布范围广。
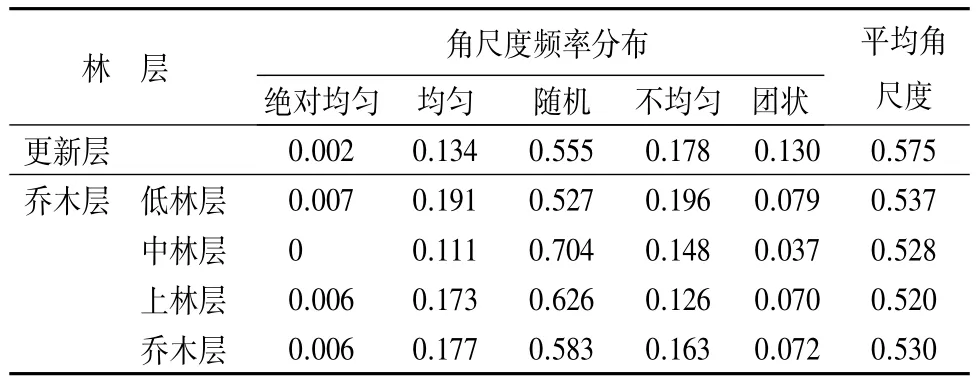
表3 各林层的平均角尺度及频率分布
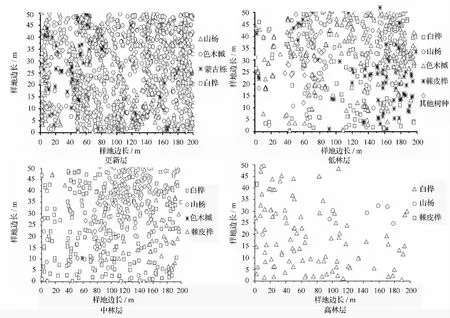
图2 各林层林木在样地中的分布
3.4 大小比数
大小比数反应了森林群落内林木的大小分化程度。由表2可知,更新层的平均大小比数为0.511,乔木层的平均大小比数为0.493,优势、亚优势、中庸、劣态和绝对劣态5种等级林木的分布频率相差不大,说明在空间结构单元内,大部分林木分化程度较高,不同等级的林木分布较为均匀。乔木层中,各林层的平均大小比数随林层的增高而降低,低林层的平均大小比数为0.705,中林层的平均大小比数为0.311,的平均大小比数为 0.105,与林分的实际生长状况相符。
分树种计算大小比数的平均值可以反映林分中树种的优势度。更新层中,竞争木优势等级由高到低的排序为:白桦、山杨、蒙古栎、色木槭;乔木层竞争木优势等由高到低的级排序为:白桦(0.37)、山杨(0.39)、棘皮桦(0.52)、色木槭(0.74)、蒙古栎(0.79)、稠李(0.88)。棘皮桦、白桦和山杨在低林层中占据优势地位,白桦和山杨在中林层中占据优势地位,与乔木层的竞争木优势等级排序基本一致。说明白桦和山杨在林分中占据优势地位,这与构成当地森林群落主要树种的事实相符,即在林分中各个径阶的林木均有分布。结合更新情况分析,白桦是群落演替的先锋树种,经过长期演替,白桦种群将逐渐以中大径木形式存在,又由于其更新不良,即将退出群落;随着演替进展,山杨将最终作为顶级树种占据优势地位;色木槭的大小比数值较高,可能与其更新时间较晚、耐荫性有关,但其更新良好,径阶分布连续,可能发展成为该地区优势树种。
3.5 开阔比数
由表4可知,乔木层的平均开阔比数为0.713,处于开阔和十分开阔状态的林木比例达65.9%,说明在次生林内大多数林木处于较开阔的光环境中,对光环境的利用较为充分。
由于空间结构单元中,参照木开阔比数的大小与相邻木的树高和距离有关。因此,不同林层或同一树种不同林层的的开阔比数大小有所差异。低林层的平均开阔比数为0.508,处于开阔状态以上的林木比例达62.9%,对光环境的利用较为充分;中林层的平均开阔比数为0.915,参照木受到完全遮挡的比例为0,74.6%的林木处于十分开阔的状态,对光环境利用充分;的平均开阔比数为0.975,参照木受到完全遮挡和遮挡的比例为0,90.1%的林木处于十分开阔的状态,对光环境的利用很充分。从树种角度分析,乔木层中,树种开阔比数从大到小依次为:山杨(0.845)、白桦(0.831)、棘皮桦(0.764)、色木槭(0.430)、蒙古栎(0.347)、稠李(0.125),说明山杨和白桦处于光利用和光竞争的优势地位。低林层中,棘皮桦的开阔比数最大,其次为白桦和山杨;中林层中,山杨的开阔比数最大,其次为棘皮桦和白桦;中,山杨和白桦均处于十分开阔的光环境中。与上述研究的大小比数优势度排序基本一致。
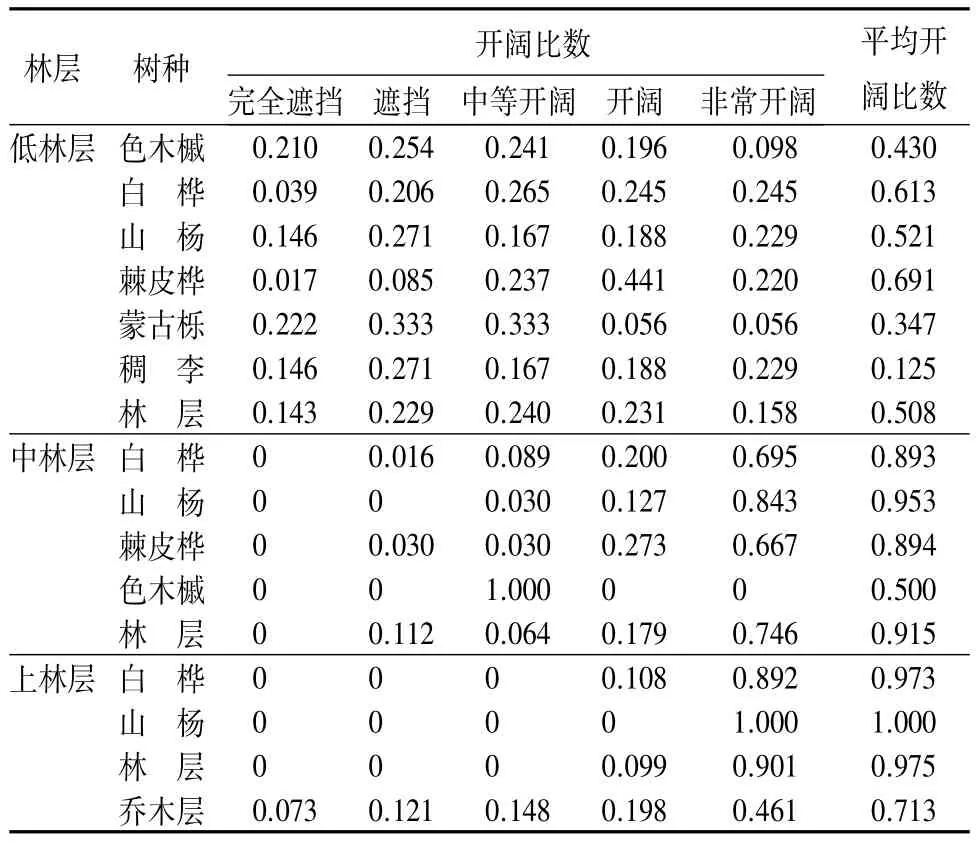
表4 各林层不同树种的开阔比数及频率分布
3.6 大小比数与开阔比数相关性
对不同树种的大小比数及开阔比数进行相关性分析。由表5可知,树种的大小比数与开阔比数之间均为负相关,即林木大小比数增大时,其开阔比数有减小的趋势。白桦、色木槭、山杨、棘皮桦及全林分的大小比数与开阔比数均呈极显著负相关。

表5 不同树种大小比数与开阔比数相关性
3.7 空间结构单元的距离尺度
为了进一步分析杨桦次生林中林木的分布状态,计算各林层由4株相邻木和1株参照木构成的结构单元,参照木与最近邻体的平均距离及参照树与4株最近相邻木的平均距离。由表6可知,随着林层的增加,最近邻体平均距离呈现递增的趋势,从0.709 m增加到1.673 m,空间结构单元平均距离也呈现出相同的趋势,从1.363 m增加到2.878 m。说明林木在生长发育过程中,随着对环境和营养条件需求的增加,竞争加剧,发生自然稀疏现象,加大了林木间距离,随后林木竞争逐渐减弱。
为了分析各林层结构单元中,4株最近相邻木的分布范围。根据调查数据,将参照木与相邻木间距离为5个组::0~1 m,≥1~2 m,≥2~3 m,≥3~4 m,≥4 m。然后统计各林层结构单元中,相邻木在各距离组出现的频率(见表7)。由表7可知,不同的林层,参照木与相邻木的距离在不同距离组的分布频率各不相同。
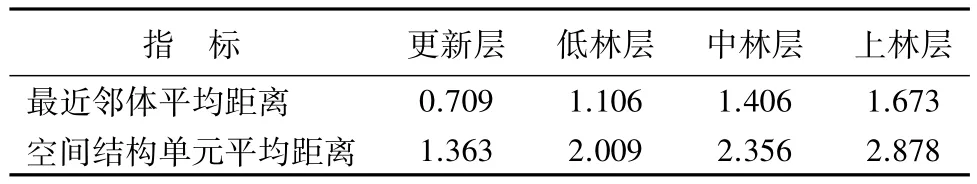
表6 各林层结构单元的距离尺度
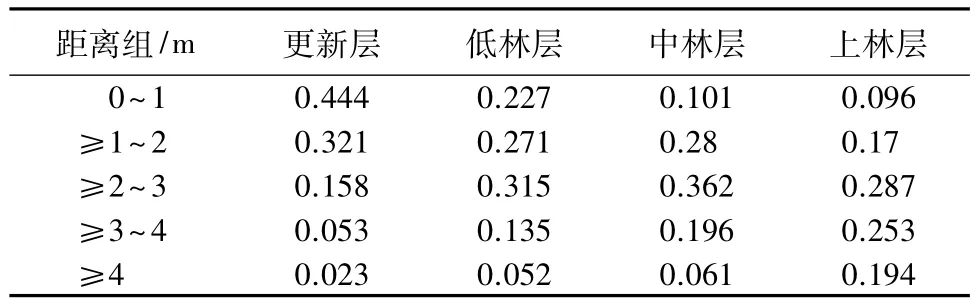
表7 各林层不同距离组相邻木出现的频率
更新层中,0~1 m的距离内分布频率最高,为44.4%,其他距离组的分布频率逐渐下降。
低林层中,从0~1 m到≥1~2 m的分布频率显著下降,其他距离组的分布频率均有不同程度的增加,最高分布频率出现在≥2~3 m,为31.5%。
中林层中,从≥1~2 m到≥2~3 m的分布频率增加,最高分布频率出现在≥2~3 m,为36.2%,从≥3~4 m到≥4 m的分布频率增加为25.7%。
高林层中,最高分布频率仍出现在≥2~3 m,但其频率下降为28.7%,从≥3~4 m到≥4 m的分布频率大幅增加,为44.7%。
因此,应根据各林层林木的生长发育特点结合其他空间结构参数,对结构单元中距离近、混交度低的弱势木进行抚育间伐,促使其形成合理的空间结构。
4 结论与讨论
通过对桃山林场杨桦次生林的空间结构特征分析发现:该地区的混交林分布不合理,林分处于不稳定状态;从树种组成来看,更新层有4个树种,乔木层有6个树种,物种丰富度不高;白桦和山杨为群落的优势种和建群种,棘皮桦和色木槭为主要伴生树种。
从各空间结构参数来看,更新层中由于以营养繁殖为主的色木槭株数比例达71.9%,且多呈丛状聚集分布,零度和弱度混交比例达73.2%,平均混交度仅为 0.228,混交状况差,平均角尺度为 0.575,聚集强度大;乔木层平均混交度为0.520,强度和极强度混交结构单元比例占39.9%,混交程度好;平均角尺度为0.530,属于聚集分布;乔木层中,聚集强度随着林层高度而降低,其原因是:随着树木个体生长对环境营养条件的需求加大,各林木为占据有利的生长空间和获得足够的环境资源空间,自疏作用和它疏作用加强,聚集强度逐渐减弱;乔木层和更新层大小比数分布频率呈现出均匀分布特征,白桦和山杨种群在空间结构单元中优势度明显;乔木层平均开阔比数为0.713,处于开阔的光环境中,但不同树种的光竞争和利用程度差异较大。乔木层中,各林层的平均大小比数随林层的增高而降低,开阔比数随林层的增高而增高,相关性分析表明,各树种及全林分的大小比数与开阔比数均呈极显著负相关。空间结构单元的平均距离随林层的增高而增加,但大部分相邻木分布在3 m以内,林木之间的竞争仍十分剧烈。
从群落的演替趋势来看,该地区的森林处于演替中前期,处于不稳定状态。在垂直结构上,上层乔木比下层乔木在生长上占优势,但优势树种在局部环境中不一定处于优势地位。白桦和山杨作为先期演替树种在一定时期内仍将保持其优势地位,但由于白桦更新不良,在演替的过程中其优势度会逐渐下降,最终退出群落。随着演替进展,山杨将最终作为顶级树种占据优势地位。色木槭径级分布连续,小径级个体较多,更新良好,可能会发展成为该地区的优势树种。其他种群个体数量少,径级分布不连续,更新不良。所以,在进行森林经营过程中,以调整合理的林分空间结构为目的,通过适当的抚育择伐措施,改善群落内生长空间分配不均匀性,增加更新树种,调整林分树种组成、水平分布格局和林内树种竞争关系,提高群落生物多样性,加速演替进行,促使林木个体和整个林分群落朝健康稳定的方向发育。
[1]惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27.
[2]惠刚盈,(德)克劳斯·冯佳多.森林空间结构量化分析方法[M].北京:中国科学技术出版社,2003.
[3]邵方丽,余新晓,吴海龙,等.冀北土石山区天然次生林山杨、白桦种群的空间分布格局[J].林业科学,2012,48(6):12-17.
[4]邵芳丽,余新晓,宋思铭,等.天然杨-桦次生林空间结构特征[J].应用生态学报,2011,22(11):2792-2798.
[5]金辉,李永宁,霍艳玲,等.冀北山地杨桦次生林空间点格局分析[J].河北农业大学学报,2012,35(3):63-68.
[6]徐学华,张慧,李永宁,等.冀北山地杨桦次生林优势种群结构及时间序列预测分析[J].河北农业大学学报,2012,35(2):72-78,87.
[7]惠刚盈,Gadow K V,Albert M.一个新的林分空间结构参数:大小比数[J].林业科学研究,1999,12(1):1-6.
[8]惠刚盈,Gadow K V,Albert M.角尺度:一个描述林木个体分布格局的结构参数[J].林业科学,1999,35(1):37-42.
[9]惠刚盈,Gadow K V,胡艳波.林分空间结构参数角尺度的标准角选择[J].林业科学研究,2004,17(6):687-692.
[10]胡艳波.基于结构化森林经营的天然异龄林空间优化经营模型研究[D].北京:中国林业科学研究院,2010.
[11]夏富才,姚大地,赵秀海,等.长白山北坡阔叶红松林空间结构[J].东北林业大学学报,2009,37(10):5-7.
[12]岳永杰,余新晓,李钢铁,等.北京松山自然保护区蒙古栎林的空间结构特征[J].应用生态学报,2009,20(8):1811-1816.
猜你喜欢
文苑(2019年20期)2019-11-20
文苑·经典美文(2019年10期)2019-10-16
现代农业研究(2017年11期)2018-01-12
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
现代园艺(2017年20期)2017-02-03
华人时刊(2016年16期)2016-04-05
学生天地·小学中高年级(2009年1期)2009-02-13
