
镉胁迫对小麦种子萌发的影响*
2014-09-17 07:46:10曲凯丽张艺馨
哈尔滨师范大学自然科学学报 2014年6期
曲凯丽,张艺馨,朱 宏
(哈尔滨师范大学)
0 引言
近年来,随着现代工农业生产的发展和人类活动的急剧增加,使大量镉进入土壤环境,严重危害农业生态环境,有关镉污染的毒害效应越来越引起人们的高度重视.重金属危害首先发生在植物种子萌发和幼苗生长阶段上种子萌发时期的生长状况直接影响作物以后的生长[1],而且进一步随着生物链危及到人类的健康和生存,研究镉污染对植物的影响具有重要的理论和实践意义.
小麦是我国主要的粮食作物之一,其萌发时期的生长状况直接影响以后的生长发育和产量,而且小麦种子萌发和幼苗时期的生长状况是评价其重金属耐性的重要指标.该实验以小麦种子为材料,研究了镉对小麦种子萌发各项指标的影响,为生产实践中早期预测重金属污染提供了一定的依据.
1 材料与方法
1.1 试验材料
选取子粒饱满、大小均匀的小麦种子若干于烧杯中,用质量分数10%的次氯酸钠消毒.消毒10 min后,用去离子水冲洗,然后置于滤纸上晾干.
1.2 试验设计
[2,7~9]中对种子发芽产生抑制效果的镉质量浓度,镉处理时采用5个质量浓度梯度,即 100、200、300、400、500μmol·L-1,记作Cd100,Cd200,Cd300,Cd400,Cd500,并以去离子水处理为对照.溶液采用分析纯 Cd(NO3)2·4H2O配置,分别称量后,以去离子水配制成各处理质量浓度.选取直径为9 cm的硬质玻璃培养皿,皿内以2层滤纸为发芽床.种子晾干后置于皿中,每皿放100粒小麦种子,然后移取等量各处理质量浓度的镉离子溶液分别加入培养皿中(以浸湿滤纸为宜),用去离子水做空白(对照).每个处理重复3次.置于25℃的人工气候箱内培养.每日滴加各溶液等量保持滤纸湿润,并记录每天发芽数(胚根与种子等长、胚芽长度达到种子50%为种子正常发芽[11]).
1.2.1 种子萌发率的测定[11-12]
以胚根突破种皮0.5 mm作为萌发的标准,7 d后计算种子萌发数.
萌发率(Gr)=(萌发种子数/供试种子数)×100%.
1.2.2 种子发芽势的测定[11-12]
发芽势(Gp)=4 d内萌发种子数/供试种子数×100%.
1.2.3 种子发芽指数的测定[11-12]
发芽指数(Gi)=ΣGt/Dt;
1.2.4 种子活力指数的测定[11-12]
活力指数(Vi)=S×Σ(Gt/Dt).
式中,Gt为t天的萌发数;Dt为发芽日数;S为幼苗根长(mm).
1.2.5 胚根鲜重的测定
萌发7 d后,取不同处理的萌发后种子的胚根用蒸馏水洗净、滤纸吸干水分,称其鲜重.
1.2.6 胚芽长度、胚根长度的测定[11-12]
发芽7 d后,每皿随机取15粒萌发种子测其芽长、根长,以其均值做为胚芽、胚根长度的指标.
1.2.7 胚芽的膜相对透性的测定[3]
称取不同镉处理后的各0.2 g,剪碎后置20 mL蒸馏水中,浸泡24 h后测电导率E1,100%水浴20 min后测相应电导率E2.膜相对透性=(E1/E2)×100%.
1.2.8 数据分析
所有数据均采用 Microsoft Excel 2003和SPSS17.0统计分析软件进行数据分析及差异显著性检验.
2 结果与分析
2.1 镉对种子萌发率的影响
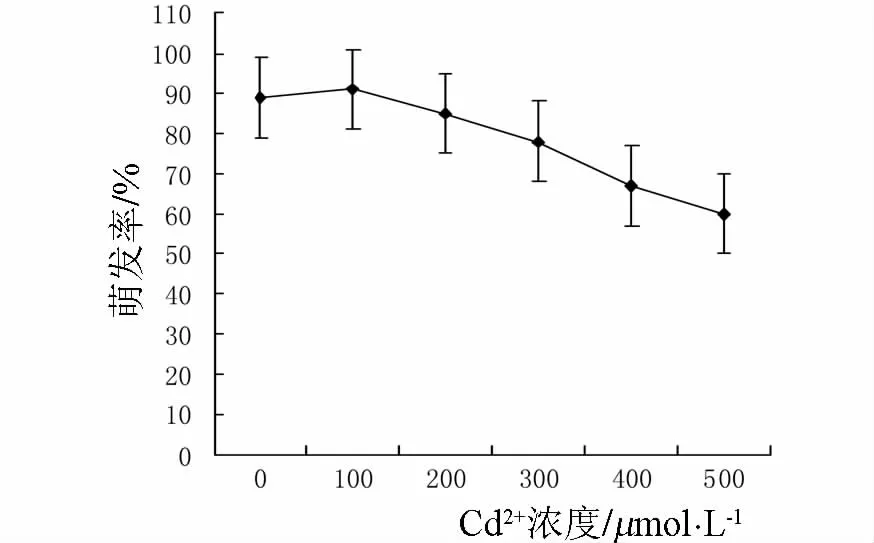
图1 镉胁迫对种子萌发率的影响
由图1可以看出,在100μmol·L-1镉浓度条件下,小麦种子的萌发率略有上升,随后不同浓度的镉离子处理对小麦种子的发芽率均有抑制作用,随着处理液浓度的升高抑制效应逐渐显著.
2.2 镉对胚根鲜重的影响
由图2可以看出,镉胁迫后,小麦胚根鲜重的下降趋势明显.浓度为100 μmol·L-1时的胚根鲜重与 CK 相差 0.21 g,浓度为 200、300、400和500 μmol·L-1时的胚根鲜重分别与CK相差1.35 g、2.03 g,2.11 g 和 2.70 g,可见当浓度高于100 μmol·L-1时胚根鲜重下降趋势较明显.说明镉对胚根生长有很强的抑制作用,且随着镉浓度的提高,镉对植物生长抑制作用增强.
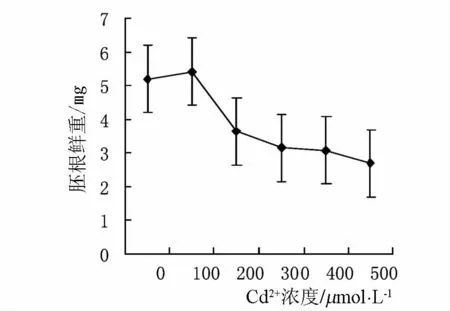
图2 镉胁迫对胚根鲜重的影响
2.3 镉对胚芽长度、胚根长度的影响
由图3可以看出,随着镉离子浓度的上升,小麦胚芽长和胚根长均呈下降的趋势.这说明随着镉浓度的增大,对胚芽和胚根生长的抑制作用增强.在浓度为 0、100、200、300、400、500 μmol·L-1时,胚芽长在各浓度之间的差分别为0.2、4、2、1.6 和 0.8 cm,在浓度大于 100 μmol·L-1时,下降趋势开始增强;胚根长在各浓度之间的差为0.2、0.8、4、5.9 和 1.3 cm.即镉处理后根长下降趋势明显,受抑显著.由此可以得出结论,镉胁迫对小麦胚根长的抑制作用比胚芽长明显.
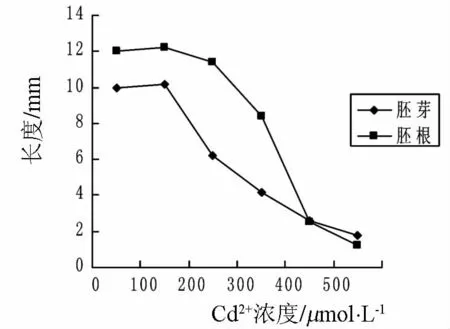
图3 镉胁迫对胚芽长度、胚根长度的影响
2.4 镉对幼芽细胞膜相对透性的影响
由图4可知,随镉离子浓度的增加,细胞膜透性总体呈增加趋势.在镉浓度为200 μmol·L-1与 300 μmol·L-1之间细胞膜透性增加趋势较明显,相差 23.9%.当浓度为 100、200 μmol·L-1时,细胞膜透性分别比对照增加了10.5%和23.9%,可见低浓度镉对小麦芽细胞膜透性影响较不明显,而高浓度镉则对细胞膜透性影响显著.
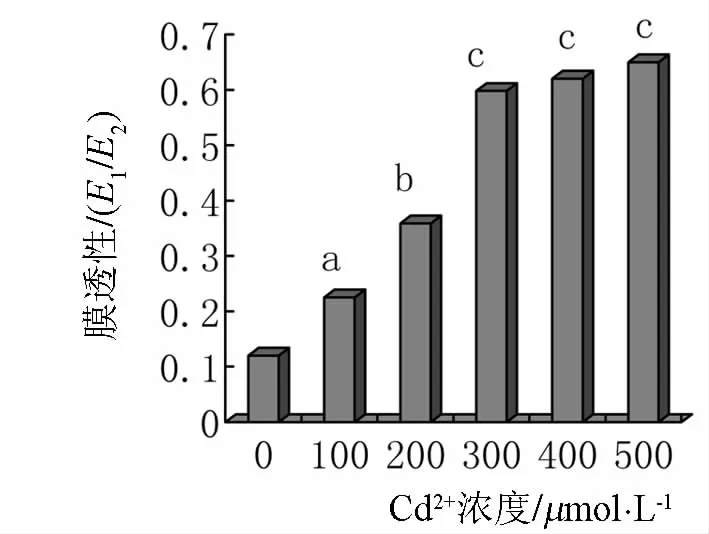
图4 镉胁迫对膜透性的影响
2.5 Cd2+对小麦种子活力指数的影响
活力指数是表征种子活力的综合指标重要指标,可反映Cd2+对小麦种子萌发的胁迫情况.由图5可见,对照小麦种子的活力指数为83,Cd2+浓度为100 μmol·L-1时种子的活力指数是85,随着镉浓度的增加活力指数显著的降低,Cd2+浓度是 200、300、400 和 500 μmol·L-1时种子的活力指数分别是 68.6、37、20 和 13.8,分别是对照的 82.7%、44.6%、24.1% 和 16.7%,镉胁迫抑制了小麦种子的活力指数.
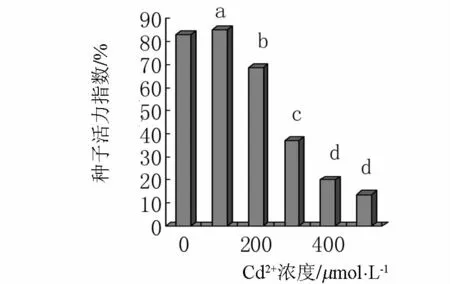
图5 镉胁迫对种子活力指数的影响
2.6 Cd2+对小麦种子发芽指数的影响
发芽指数能反映种子萌发的快慢和整齐程度.随着Cd2+处理浓度的升高对小麦种子的发芽指数的抑制作用逐渐增强.对照小麦种子的发芽指数是22,在低浓度Cd2+处理下发芽指数为25,从 200 μmol·L-1镉处理开始随着镉浓度的升高种子的发芽指数逐渐降低,当镉浓度是200、300、400、500 μmol·L-1时小麦种子的发芽指数分别是 17.7、15、10、8,分别是对照的80.5%、68.2%、45.5%、36.4%(如图 6 所示),从200 μmol·L-1镉处理开始,发芽指数急剧降低.
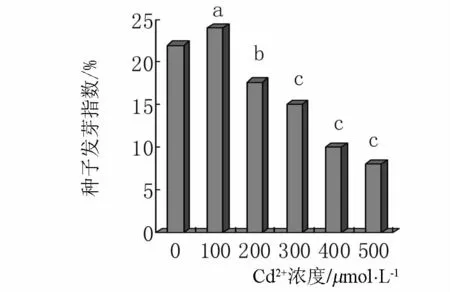
图6 镉胁迫对种子发芽指数的影响
2.7 Cd2+对小麦种子发芽势的影响
发芽势是反映种子质量优劣的主要指标之一,也是鉴别种子发芽整齐度的重要指标.从图7可见,随着Cd2+浓度的增加小麦种子的发芽势不断降低,高浓度时降低显著.对照小麦种子的发芽势是91%,100 μmol·L-1镉处理下发芽势是 93%,当 Cd2+浓度是 200、300、400 μmol·L-1时,种子的发芽势分别是75%、68%、52%,与对照相比显著的降低.Cd2+浓度为500 μmol·L-1时,种子发芽势是42%,是对照的46.2%,与对照相差较大.
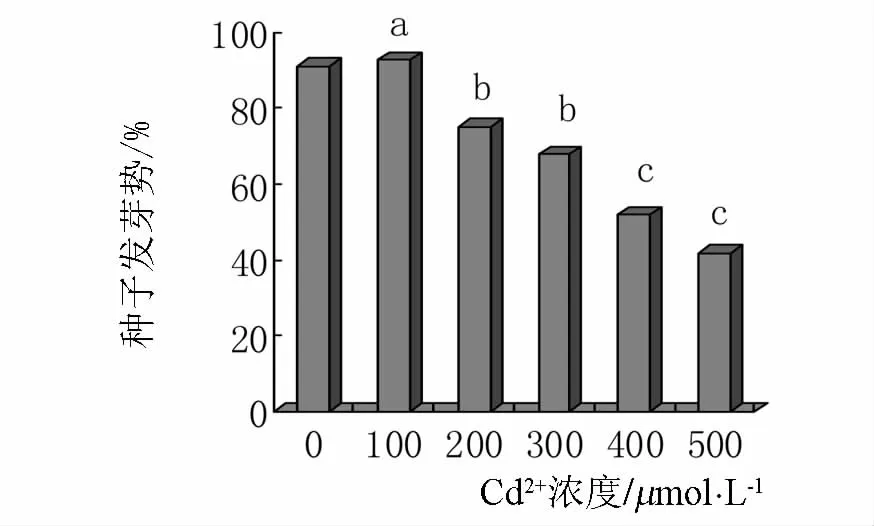
图7 镉胁迫对种子发芽势的影响
3 结论
种子萌发是植物生命起始的重要事件,也是植物最早接受重金属胁迫的阶段.了解种子萌发对镉的胁迫反应,是系统认识重金属伤害机理的较好途径.
不同镉质量浓度对小麦种子萌发影响试验的研究结果表明:随着镉离子浓度的增加,小麦种子的发芽率、胚芽长、胚根长、胚根鲜重、种子发芽势、发芽指数和活力指数迅速下降,镉离子浓度越大,作用时间越长,抑制效应越强,镉影响了芽细胞膜的相对透性,可见高浓度重金属镉对细胞膜的破坏作用较明显.低质量浓度(100 μmol·L-1)下,小麦种子的发芽率、发芽势和发芽指数等稍有上升,表明低质量镉浓度对小麦种子的萌发有促进作用,这与许多研究得到的结论一致[1,13,18-19].造成这种情况的原因可能是低质量浓度可提高胚的生理活性[19-20].高质量浓度对这种植物种子的萌发有较强的抑制作用.这可能与镉质量浓度过高会抑制种子内贮藏淀粉和蛋白质的分解,从而影响种子萌发所需要的物质和能力有关[19-20].
细胞膜透性的变化主要是由于膜脂过氧化,膜蛋白变性及膜脂流动性改变,造成膜相变和膜结构破坏所致,其中丙二醛(MDA)含量的变化是反应膜脂过氧化强弱的一个重要指标.Ewa等的研究表明随着Cd和Cu浓度的增加拟南芥的膜脂过氧化和MDA的含量不断增加[21].芥菜型油菜MDA含量随镉处理浓度增大而呈上升趋势[22].在植物中,膜脂过氧化水平跟 Lipoxygenase的活性有关,而细胞内MDA的含量的增加随着自由基(O2·和 OH·)的水平的升高而增加[23-24].
本试验结果还表明,镉胁迫对小麦胚根长的影响大于对胚芽长的影响,其可能是由于重金属与植物作用时,总是最先接触到根部,根细胞壁中存在大量交换位点,能将重金属离子固定在这些位点上,从而阻止重金属离子进一步向芽转移[25].
参考文献
[1] 任朋娟,孟昭福,马云飞,等.BS-12和Cd2+复合污染对空心菜种子萌发及生长的影响[J].西北农业学报,2011,20(4):129-133.
[2] 宗宪春,张永乐,司徒琳莉等.镉胁迫对三种花卉种子萌发的影响[J].湖北农业科学,2012,51(4):744-746.
[3] 金美芳,林明凤,施翔,等.镉胁迫对油菜种子萌发和根系生长的影响[J].种子,2011,30(10):70-73.
[4] 刘垚,张薇,刘鸣达.镉-苯并(α)芘单一及复合污染对小麦种子萌发的影响[J].农业环境科学学报,2012,31(2):265-269.
[5] 王素平.H2O2对镉胁迫下黄瓜种子萌发及生长的影响[J].种子,2012,31(3):98-100.
[6] 陶玲,任琚,祝广华,等.重金属对植物种子萌发的影响研究进展[J].农业环境科学学,2007,26(增刊):52-57.
[7] 魏树和,杨传杰,周启星.三叶鬼针草等7种常见菊科杂草植物对重金属的超富集特征[J].环境科学,2008,29(10):2912-2918.
[8] 赵勇,李红娟,孙志强.土壤、蔬菜的Cd污染相关性分析及土壤污染阈限值研究[J].农业工程学报,2006,22(7):149-152.
[9] 高洁,周永刚,张琪等.污泥和盐渍土混配介质的性质及对5种野生植物生长的影响[J].生态与农村环境学报,2012,28(3):294-299.
[10]曾翔,张玉烛,王凯荣等.镉对水稻种子萌发的影响[J].应用生态学报,2007,18(7):1665-1668.
[11]郑光华.种子活力的原理及其应用[M].北京:科学出版社,1986.154.
[12] 胡晋.种子生物学[M].北京:高等教育出版社,2006.191.
[13]鲁先文,丁时和.Cd,Pb污染对油菜种子萌发及幼苗生物量的影响[J].中国农学通报,2011,27(6):124-127.
[14]白瑞琴,晁公平,孙华等.重金属镉胁迫对蜀葵、二月蓝种子萌发和幼苗生长的毒害效应研究[J].华北农学报,2009,24(2):134-138.
[15]周云龙.植物生物学[M].北京:高等教育出版社,2004.25-39.
[16]宋玉芳,周启星,许华夏等.重金属对土壤中小麦种子发芽与根伸长抑制的生态毒性[J].应用生态学报,2002,13(4):459-462.
[17]欧丽,刘足根,方红亚等.Cd对野茼蒿种子发芽的影响[J].生态毒理学报,2011,6(4):441-444.
[18]慈恩,高明,王子芳等.镉对紫花苜蓿种子萌发与幼苗生长的影响研究[J].中国生态农业学报,2007,15(1):96-98.
[19]周青,黄晓华,张一等.镉对种子萌发的影响[J].农业环境保护,2000,19(3):156-158.
[20]葛成军,陈秋波,俞花美,等.Cd胁迫对2种热带牧草种子发芽与根伸长的抑制效应[J].热带作物学报,2008,29(5):567-571.
[21] Ewa S P,Maria D,Zbigniew K.Lipid peroxidation and antioxidative response in Arabidopsis thaliana exposed to cadmium and copper[J].Acta Physiol Plant,2010,32:169-175.
[22]刘丽莉,冯涛,向言词.外源钙对镉胁迫下芥菜型油菜幼苗生长和生理特性的影响[J].农业环境科学学报,2009,28(5):978-983.
[23] Skorzynska-Polit E,Drazkiewicz M,Krupa Z.The activity of the antioxidative system in cadmium-treated Arabidopsis thalianan[J].Biol Plant,2003,47:71-78.
[24]Skorzynska-Polit E,Krupa Z.Activity of lipoxygenase in Arabidopsis thaliana-a preliminary study[J].Cell Mol Biol Lett,2003(8):79-84.
[25]闫华晓,赵辉.镉离子对玉米种子萌发和生长的影响[J].作物杂志,2007(5):25-28.
猜你喜欢
广西林业科学(2021年6期)2022-01-21 07:39:28
山西农业科学(2021年8期)2021-08-17 06:20:10
青年文学家(2021年4期)2021-03-18 07:00:23
农民致富之友(2020年19期)2020-07-18 15:32:39
山西农业科学(2020年1期)2020-02-27 17:00:54
延边大学学报(自然科学版)(2019年2期)2019-10-08 03:03:22
家庭百事通·健康一点通(2019年11期)2019-09-26 09:27:52
广西林业科学(2016年2期)2016-03-20 05:53:25
生物加工过程(2015年2期)2015-11-11 06:02:32
饮食科学(2014年8期)2014-08-22 17:42:07
