
不同浓度七氟烷对大鼠认知功能及海马内磷酸化cAMP反应元件结合蛋白表达的影响
2014-09-13 03:17施小娇张晓暄宋雪松
中国老年学杂志 2014年20期
施小娇 金 梦 张晓暄 宋雪松
(吉林大学第一医院麻醉科,吉林 长春 130021)
异氟烷可诱发发育期动物脑神经毒性并影响成年后的学习记忆功能,七氟烷的研究结论不一,但已有其麻醉可增加婴幼儿术后认知功能障碍的发生率的报道,其机制还有待进一步阐明〔1~5〕。学习和记忆相关的海马神经元长时程增强(LTP)是一种突触可塑性的变化,其诱导和维持中均需要如N-甲基-D-天冬氨酸(NMDA)、γ-氨基丁酸(GABA)等受体的激活及相继的胞内激酶级联反应和核内相关基因转录和蛋白表达水平的改变,由核内第三信使介导的转录和表达变化是海马神经元早期LTP向稳定的晚期LTP转化及记忆巩固的关键环节。大量研究表明,核转录因子cAMP反应元件结合蛋白(CREB)参与介导上述效应,其磷酸化水平下降,引起与学习记忆功能密切相关的海马CA1区LTP被抑制〔6~8〕。本研究拟观察不同浓度七氟烷麻醉对大鼠早期认知功能的影响以及海马中磷酸化CREB表达的变化。
1 材料与方法
1.1动物分组与麻醉模型的建立 SPF级5周龄SD大鼠90只,体质量100~110 g,随机分为对照组(C组)、1%七氟烷麻醉组(S1组)、2%七氟烷麻醉组(S2组)、2.6%七氟烷麻醉组(S3组)、4.1%七氟烷麻醉组(S4组)。C组单纯吸氧6 h,氧流量2.0 L/min,其余各组在与C组相同的氧流量条件下分别给予1%、2%、2.6%和4%七氟烷吸入6 h,待翻正反射恢复后将大鼠放回饲养笼。
1.2主要仪器与试剂 麻醉气体监测仪(飞利浦S60,荷兰);Morris水迷宫实验装置(上海软隆科技发展公司);七氟烷挥发罐(艾伯维医疗贸易上海有限公司);兔抗pCREB抗体(Santa Cruz,美国)
1.3七氟烷吸入麻醉 吸入麻醉过程在一自制麻醉箱内进行。自制麻醉箱为有机玻璃材料制成,高、宽、长为25 cm×15 cm×15 cm,用带有3 mm小孔的筛状隔板将箱内空间分割为上(高15 cm)、下(高10 cm)2部,箱顶近中央分别有一大孔和小孔,大孔嵌装的气管导管标准衔接头,可与麻醉机螺纹管紧密连接;箱的两侧面和背面高于隔板2 cm处各有一小孔,每个小孔(包括顶部)分别嵌装一可与多功能气体分析仪自动取样管良好吻合的标准连接头。实验时,在麻醉箱底部放置一薄层钠石灰用以吸收CO2,调节麻醉机(Ohmeda Detax,美国)氧流量(2.0 L/min)和七氟烷蒸发罐,当通过麻醉箱各侧小孔测定箱内七氟烷浓度达到预定目标:S1组1%〔0.5最低肺泡有效浓度(MAC)〕、S2组2%(1 MAC)、S3组2.6%(1.3 MAC)和S2组4.1%(2 MAC),将大鼠放置于麻醉箱上部隔板上,6 h后关闭麻醉蒸发罐,待其麻醉自然消退和清醒;C组仅向箱内输送相同浓度的氧气6 h,但不开启七氟烷蒸发罐。
1.4Morris水迷宫实验 在七氟烷麻醉后1、2、3、4、5和6 d进行。定向航行实验5 d:每天4次,将大鼠分别从4个象限入水点面向池壁放入水中,记录60 s内寻找到平台的时间并让大鼠在平台上停留10 s,超过60 s未能找到平台者,用木棒轻柔人为地将其引上平台并停留在台上10 s,认定其寻找平台时间为60 s,取4次测定结果的平均值为当天的逃逸潜伏期。空间搜索实验:在七氟烷麻醉后第6天观察空间探索能力,从水槽内移走平台,自第三象限将大鼠面向槽壁放入水中,记录60 s内在原平台处穿台次数以及在平台象限(第三象限)内的活动时间。
1.5旷场实验 旷场行为观察箱为80 cm×80 cm× 60 cm的无顶木箱,箱底用墨线分为25个等份小方格。在七氟烷麻醉后1 d和5 d,自中央格放入大鼠,观察在2 min内穿越的格子数(3只脚进入其他小方格)和直立次数(两前腿同时离开底面)。每完成一只大鼠的测试后清理木箱内壁和底面,以免大鼠气味残留影响下次实验。
1.6免疫印记分析 于七氟烷麻醉后苏醒后,每组取6只大鼠在1%戊巴比妥钠60 mg/kg深麻醉下取左侧海马,放入匀浆液中低温匀浆,15 000 r/min, 4℃离心20 min,取上清。按Bradford法测定样本蛋白浓度后加入4倍样本缓冲液(250 mmol/L Tris/HCl,0.01% 考马斯亮蓝-G,8% SDS,200 mmol/L蔗糖,和300 mmol/L Dithiothreitol,pH6.8),95℃ 5 min变性。30 μg等量的样本加在10%的SDS聚丙烯酰胺凝胶上电泳(120 V,1.5 h),然后电转至硝酸纤维素膜上(100 mA,2 h),5%脱脂奶粉封闭30 min后与兔抗pCREB多克隆抗体(1∶400)和β-tubilin多克隆抗体 (1∶1 000) 4℃孵育过夜,TBST冲洗,3×10 min,加入碱性磷酸酶标记的羊抗兔二抗 (1∶1 000),室温振荡孵育1 h,TBST冲洗3×10 min,用NBT/BCIP检测反应条带,扫描图像用Image J 软件(NIH,USA)半定量分析。
1.7统计学处理 采用SPSS10.0软件进行配对t检验和单因素方差分析。
2 结 果
2.1各组学习记忆能力比较 与C组相比,麻醉后1、2 d,S1、S2、S3和S4组平均逃逸潜伏期均明显延长(P<0.05);S1组平均逃逸潜伏期与C组差别无统计学意义(P>0.05);麻醉后3 d,S3和S4组大鼠平均逃逸潜伏期均明显延长(P<0.05),麻醉后4 d, S4组平均逃逸潜伏期均明显延长(P<0.05),麻醉后5 d,各组平均逃逸潜伏期未见明显差异(P>0.05)。见表1。
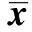
表1 各组麻醉后不同时间平均逃逸潜伏期的比较(s,±s,n=10)
2.2各组空间搜索能力比较 与C组相比, S1和S2组平均平台象限停留时间和穿台次数之间差别无统计学意义(P>0.05),S3和S4组明显减少(P<0.05)。见表2。
2.3各组旷场内自发行为比较 各组麻醉后1和5 d在旷场箱内的自发行为表现差异无统计学意义(P>0.05)。见表3。
2.4各组海马磷酸化CREB表达的影响 与C组(1.0)相比,S2、S3和S4组海马组织中磷酸化CREB表达(0.95±0.04、0.74±0.06、0.61±0.05)明显下降(P<0.05),而S1组(0.93±0.07)与C组差别无统计学意义(P>0.05)。见图1。
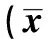
表2 各组平台象限停留时间和穿过平台次数的比较±s,n=10)
表3 各组旷场试验结果的比较±s,n=10)

图1 各组海马磷酸化CREB表达
3 讨 论
学习记忆是人类和动物共同的赖以生存的最基本的高级神经功能之一,已知人类中枢神经系统的快速发育期主要在2~3岁,此时期大脑的发育及改变必将会影响成年后大脑的功能变化。含氟吸入性麻醉药(如异氟烷和七氟烷)是目前临床全身麻醉中广泛使用的一类药物,特别是七氟烷因起效迅速、无刺激性气味已广泛用于婴幼儿麻醉的诱导和维持。已知单独应用七氟烷麻醉,其MAC为2.05%(ED50),而吸入浓度达到1.3MAC(2.6%)时才认为达到麻醉标准(ED95)。
Morris水迷宫实验是一种能够客观评价实验动物认知功能的方法,其中定向航行的逃逸潜伏期可反映获取空间信息的能力,而空间探索实验的行为反映了实验动物对在定向航行实验训练中所获取信息的记忆储存及再现能力。本研究说明较高浓度七氟烷麻醉后大鼠早期学习空间信息的能力及记忆功能可能受到了影响,这种学习记忆影响仅仅是短暂和早期的。朱霞等〔9〕观察七氟烷对幼鼠麻醉前的学习记忆影响的研究提示在临床麻醉中尽量减低吸入麻醉药的浓度可能更安全。
七氟烷,包括其他含氟挥发性吸入麻醉药影响生长期的神经细胞的结构和功能发育仍未完全阐明,但有研究〔3~5〕指出GABA受体和NMDA受体活性均参与调节生长期和成年期神经细胞的增殖、分化,除了引起发育中的动物脑神经元结构异常外,还可能造成其成年后脑神经元密度下降以及长期的认知功能障碍,此两种受体也在许多全身麻醉药作用机制中发挥重要作用。磷酸化是细胞信号通路中转录因子活性调控的主要机制。CREB是最早证实由磷酸化调控其活性的转录因子之一,在脑内所有细胞中均有表达,定位于核内并在多种信号分子诱导下调控大量下游靶基因的表达。CREB作为一种核内转录因子,磷酸化后能够激活或促进一系列与学习记忆相关的基因表达,表达长时间记忆相关蛋白,在短时间记忆转变为长时间记忆过程中起到关键的分子开关作用〔10〕。本研究说明CREB磷酸化水平的降低可能参与介导引起幼鼠七氟烷麻醉后学习记忆能力下降。CREB可在不同信号通路的激酶作用下实现Ser133磷酸化而活化发挥生物学效应,全身麻醉药通过兴奋GABA受体或抑制NMDA受体引起细胞内激酶级联信号传导通路如cAMP/cAMP-依赖性蛋白激酶A (cAMP-PKA)、钙/钙调蛋白依赖性蛋白激酶Ⅳ(CaMKⅣ)、CaMⅡ、Ras-胞外信号调节激酶-核糖体S6激酶(Ras-ERK-RSK)等的变化,进一步引起CREB活性的变化,导致短期内学习记忆能力的损害。
4 参考文献
1Lei X,Guo Q,Zhang J.Mechanistic insights into neurotoxicity induced by anesthetics in the developing brain〔J〕.Int J Mol Sci,2012;13(6):6772-99.
2Loepke AW,Soriano SG.An assessment of the effects of general anesthetics on developing brain structure and neurocognitive function〔J〕.Anesth Analg,2008;106(6):1681-707.
3Liang G,Ward C,Peng J,etal.Isoflurance causes greater neurodegeneration than an equivalent exposure of sevoflurance in the developing brain of neonatal mice〔J〕.Anesthesiology,2010;112(6):1325-34.
4Millar K,Asbury AJ,Bowman AW,etal.The effects of brief sevoflurance-nitrous oxide anesthesia upon children’s postoperative cognition and behavior〔J〕.Anesthesia,2006;61:541-7.
5Sun L.Early childhood general anaesthesia exposure and neurocognitive development〔J〕.Br J Anaesth,2010;105(s1):i61-8.
6Meyer CJ,Alenghat FJ,Rim P,etal.Mechanical control of cyclic AMP signalling and gene transcription through integrins〔J〕.Nature Cell Biol,2000;2(9):666-8.
7Schinelli S,Zanassi P,Paolillo M,etal.Stimulation of endothelin B receptors in astrocytes induces cAMP response element-binding protein phosphorylation and c-fos expression via multiple mitogen-activated protein kinase signalin pathways〔J〕.J Neurosci,2001;21(22):8842-53.
8Bing G,Ottavio V,Fabrizio T,etal.Persistent improvement in synaptic and cognitive functions in an Alzheimer mouse model after rolipram treatment〔J〕.J Clin Invest,2004;14(11):1624-34.
9朱 霞,朱贵芹,张励才.不同浓度七氟醚对幼鼠学习记忆及海马NR2B Tyr472磷酸化表达的影响〔J〕.中国药理学通报,2013;29(12):1717-20.
10Paramanik V,Thakur MK.Role of CREB signaling in aging brain〔J〕.Arch Ital Biol,2013;151(1):33-42.
猜你喜欢
现代食品科技(2022年8期)2022-09-02
中学生数理化·高一版(2022年3期)2022-04-05
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2021年12期)2021-12-30
中国药学药品知识仓库(2021年18期)2021-02-28
云南医药(2020年5期)2020-10-27
初中生学习指导·提升版(2020年10期)2020-09-10
山东医药(2020年14期)2020-06-05
天津医科大学学报(2019年6期)2019-08-13
分析化学(2017年12期)2017-12-25
