
内质网应激相关蛋白在大鼠急性脊髓损伤中的表达及其意义
2014-09-13 03:17田小磊靳安民
中国老年学杂志 2014年20期
田小磊 王 坤 靳安民
(南方医科大学珠江医院骨科,广东 广州 510282)
脊髓损伤(SCI)可造成患者功能障碍并留下终生残疾,目前关于SCI的治疗和愈合护理尚存在争议,故对于如何最大限度降低患者残疾程度,恢复其脊髓功能在医学界成为研究热点〔1,2〕。研究发现,SCI不但能造成细胞坏死,同时能造成细胞发生凋亡现象,从而影响脊髓的相关功能〔3〕。过去认为细胞凋亡的主要途径为线粒体途径和死亡受体途径〔4〕,如今在一些研究中发现,内质网应激(ERS)也能造成细胞凋亡,其中转录因子(C/EBP)家族中的CHOP/GADD153是特异的ERS转录因子〔5〕。本研究通过观察大鼠急性SCI前后其脊髓组织中内质网应激相关蛋白(CHOP)的表达情况以及神经细胞的凋亡变化,从而进一步探讨ERS激对于急性SCI的意义。
1 资料与方法
1.1研究对象 选取56只6~8周龄健康 SD 大鼠 (湖南省人民医院动物实验室提供),雌雄各半,体重 320~400 g。随机分为对照组和实验组,每组28只,按损伤时间点将每组分为3、24 h、3、7 d四组(每组7只),每组均在脊髓损伤后计算时间,并于相应时间点进行神经功能Tarlov评分,而后处死动物进行心脏灌注和组织观测试验。
1.2建立动物模型 以Allen法作为参照制作脊髓损伤大鼠模型。实验组制作方法:(1)腹腔注射1%戊巴比妥钠(0.1 ml/kg)麻醉大鼠后,固定大鼠四肢及门牙于解剖板上;(2)剪净大鼠胸背部T10处及其周围4 cm2面积的毛,使其皮肤组织暴露;(3)用3%络合碘进行皮肤消毒,在T10水平背部正中线做2~3 cm纵行切口,依次切开皮肤、筋膜,并可见白色腱膜;(4)剥离棘突两侧的肌肉暴露T9~T11棘突和椎板;(5)切除T9、T10椎板,暴露硬膜囊范围约1.0×0.4 cm2大小;(6)用10 g质量的条形金属撞击器控制2.5 cm高度在导向器内做自由落体打击导向器底部,导向器嘴部撞击硬膜囊致脊髓中度损伤,若大鼠尾巴和后肢发生抽搐,则急性SCI大鼠模型制作成功;(7)逐层关闭切口,以敷料覆盖伤口并固定,单笼饲养;(8)每天每只大鼠以青霉素和庆大霉素联合抗感染,生存时间长于3 d者共给药3 d,并辅以人工排尿、排便及清理伤口等处理。 对照组的大鼠只做T9、T10椎板切除术,不进行脊髓撞击,其余处理与实验组相同。
1.3评定神经功能 分别于术后3、24 h、3、7 d观察实验组和对照组大鼠后肢运动情况,参照改良 Tarlov 标准进行评分:0分,后肢无主动活动;1分,后肢可小范围活动,无行走能力;2分,后肢活动频繁或有力、无行走能力;3分,可行走,步态严重障碍;4 分,可行走,步态轻度障碍;5分,可正常行走。
1.4心脏灌注和组织观测 (1)心脏灌注:①分别在相应时间点各取7只大鼠腹腔注射1%戊巴比妥钠(0.1 ml/kg) 麻醉大鼠后固定于解剖板,打开大3鼠胸腔暴露心脏;②用止血钳牵拉,将7号半输液器针头插入大鼠左心房直至主动脉内,剪开右心耳;③将生理盐水100 ml快速灌注大鼠心脏,控制流速确保血管不鼓破;④用4%多聚甲醛快速灌注约15 min后,控制滴速(20 滴/min)再灌注3 h。(2)组织提取:①以T8~T10为中心,取2 cm左右的损伤节段脊柱,仔细咬去椎板、椎体和周围瘢痕组织,剥离除完整的脊髓组织;②将磷酸缓冲液清洗脊髓组织,于4%多聚甲醛中进行固定保存。 (3)组织学观察:①用石蜡常规包埋,于损伤中心部位连续切片5张,5 μm厚;②随机抽取2张切片均进行免疫组织化学染色;③采用HPIAS-1000图像分析系统,于高倍镜下随机选择5个视野,测定累积光密度IOD,取其平均值。
1.5缺口原位末端标记法(TUNEL) 染色 随机抽取1张脊髓损伤组织切片进行TUNEL染色。染色方法:(1)将切片脱蜡入水,在室温下用3%H2O2处理10 min,0.01 mol/L磷酸缓冲液清洗1次,在温度37℃下用1∶200 蛋白酶 K消化10 min,于200 ml 0.1 mol 柠檬酸钠的湿盒中放置切片;(2)用微波修复5 min,0.01 mol/L磷酸缓冲液清洗2次;(3)在切片中滴加脱氧核糖核苷酸末端转移酶(TdT)及标记液(TUNEL 试剂盒来自Roche公司 );(4)在荧光显微镜激发强度为450~500 nm下观察其荧光表达(绿光,波长为515~565 nm);(5)在40倍镜下采集图像,若发出绿色荧光,说明为阳性细胞,且每张切片选取5个视野进行观察,可用平均光密度描述细胞凋亡的情况。
1.6统计学方法 应用SPSS17.0软件,7 d时点组进行两因素重复测量分析;实验组进行单因素方差分析;CHOP 表达和细胞凋亡资料进行两因素析因方差分析;重复测量分析中的精细多次比较,按Bonferroni校正法进行显著性水平α’的调整。
2 结 果
2.1神经功能改良Tarlov评分 术后实验组大鼠出现了不同程度的后肢运动功能障碍,而对照组各大鼠均未见明显运动功能损害。见表1。整体分析发现:两组4时点间,组别与时间的交互作用等均有显著性意义(P<0.05)。再进行精细比较,在各个时间点,两组间的差异均显著(P<0.05);经配对t检验,仅实验组的3 d和3 h两时点比较有统计意义(P<0.05),其他时点间比较均不显著。实验组大鼠后肢运动功能障碍在3 d时最明显,另外各时间点对照组大鼠改良Tarlov评分均高于实验组,且实验组大鼠与对照组改良Tarlov评分差别均较大。
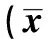
表1 7 d时点组大鼠急性SCI后不同时间点改良Tarlov评分±s,n=7)
考虑到Tarlov评分观测,都是在大鼠被处杀前,4个时间组分别进行了1~4次评分观测,这些多次评分尤其是实验组的多次评分,都携有不可忽略的试验信息,此数据的重复测量性质已被弱化,故按4个时间独立组进行分析(单因素方差分析)。3 h:3.82±0.51,1 :3.37±0.54,3 d:2.32±0.62,7 d:3.43±0.50。分析结果为:整体分析为显著性差异(P<0.05),两两比较也均有统计学意义(P<0.05)。
2.2组织学检查 对照组大鼠急性SCI组织切片中未见明显坏死细胞与凋亡细胞。 实验组当时间在3 h时大鼠脊髓组织主要以坏死细胞为主,并出现血肿、脂肪液化、细胞核碎裂、细胞溶解现象,当时间在1 d 时出现较明显凋亡细胞,并伴有神经细胞固缩以及凋亡小体,3 d时凋亡细胞更多,但7 d时凋亡细胞数则有所降低。
2.3CHOP 表达情况 整体比较:两个组间,4个时点间,分组及时间的交互作用均显著(P<0.05),提示CHOP水平两组不同,4个时点间不同,各组随时间的变化情况不同。见表2。
表2 各组大鼠急性SCI后不同时间点CHOP(累积光密度)表达±s,n=7)
结合数据来看:对照组大鼠急性SCI组织切片中免疫阳性细胞表达不明显。实验组在3 h后就有CHOP表达,SCI后1 d时CHOP表达虽有所升高,但与3 h相比差别不大;SCI 3 d时CHOP表达达到高峰,7 d时又有所下降,且SCI 1 d和7 d时CHOP的表达均与3 d时差别明显。另外,各时间点实验组CHOP表达与对照组均有较大差别。
2.4细胞凋亡情况 分析结果显示:整体比较,两个组间,4个时点间,分组及时间的交互作用差异显著(P<0.05),提示CHOP水平两组不同,4个时点间不同,各组随时间的变化情况不同。见表3。
结合数据来看:实验组3 h时细胞凋亡情况与对照组差别不明显,而1 d、3 d、7 d时两组细胞凋亡均具有较大差别;实验组3 h和1 d之间、1 d和3 d之间,3 d和7 d之间细胞凋亡均差别较大,其中3 d时细胞凋亡最为明显。
表3 各组大鼠急性SCI后不同时间点TUNEL(平均光密度)表达±s,n=7)
2.5CHOP 表达与细胞凋亡的相关性 CHOP表达与细胞凋亡情况,在时间上两者的趋势表现相似,均于SCI 3 d后达到高峰。两组相关分析得到 Pearson相关系数为0.729,且P=0.040,说明CHOP的表达与细胞凋亡具有直线相关。
3 讨 论
SCI因能够使患者发生功能障碍并造成终生残疾,严重影响了患者生活质量,降低了劳动生产率,增加了家庭和社会的负担〔6〕。一般情况下,SCI可以分为原发性SCI和继发性SCI。原发性SCI是由于机械性外力造成脊髓组织被破坏造成出血、坏死等损伤,而继发性SCI是在原发性SCI后,由于组织损伤部位及附近部位发生缺血再灌注损伤、脂质过氧化、兴奋性神经递质损伤、产生炎症介质、发生神经细胞凋亡等一系列生理病理变化而造成的SCI〔7〕。SCI的最终结局不仅与原发性损伤有关,同时还跟继发性损伤有直接联系。关于继发性损伤的机制目前尚不明确,一直是医学研究中的难题,通过物理或药物等人工干预方法进行治疗和护理,对于患者功能障碍的恢复及提高其生活自理能力具有重要意义〔8〕。
细胞凋亡是在病理条件下细胞内发生的一种程序性细胞死亡,目前有研究发现细胞凋亡能严重影响SCI后的神经功能,是造成脊髓发生继发性损伤的主要因素〔3〕。过去认为细胞凋亡的主要有两种途径即线粒体途径和死亡受体途径,如今在一些研究中发现,ERS也能造成细胞凋亡〔9〕,其中转录因子C/EBP家族中的CHOP/GADD153是特异的ERS转录因子〔10〕。
内质网是真核细胞中非常重要的一种细胞器,包括钙离子的调节、蛋白质的加工合成等。内质网中具有多种酶,且能对机体状态进行即时监控,当各种生理病理原因导致内质网功能发生变化时,内质网将进行对应应答反应即ERS反应。内质网发生应激反应的主要标志是在内质网内应激蛋白的诱导表达,包括内质网分子伴侣葡萄糖调节蛋白78(GRP78)、GRP94和协调因子153(GADD153)。当GRP78和GADD153表达升高说明细胞发生了ERTS〔11〕。若应激强度过强及时间过长使内质网处于病态状态导致其功能难以修复,从而能促使C反应蛋白(CRPs)、蛋白质折叠酶以及钙网蛋白等物质形成增加以增强细胞的耐受能力〔12〕。若存在持续严重的内质网应激时,机体能使一些促凋亡因子如caspase-12、CHOP/GADD153表达,激活内质网凋亡途径,从而造成细胞的凋亡和组织的损伤。
综上所述,急性SCI早期主要表现为细胞坏死,后期细胞凋亡数增多。急性SCI能促使内质网发生应激,同时能够增加促凋亡因子CHOP的表达。CHOP表达水平早期时比较低,3 d时CHOP表达水平明显升高,引起细胞凋亡数也增多,CHOP表达水平与细胞凋亡具有直线相关关系。
4 参考文献
1唐智生,毛容秋,陈 莹,等.脊髓损伤245例临床特点分析〔J〕.广西医学,2012;34(8):996-7.
2李志强,权 哲.脊髓损伤修复机制和治疗展望〔J〕.中国基层医药,2012;19(14):2206-9.
3李 剑.CHOP在内质网应激介导凋亡中的作用〔J〕.检验医学与临床,2011;8(2):208-11.
4陈 瑜,刘翠萍,茅晓东,等.糖基化终产物诱导小鼠足细胞内质网应激和细胞凋亡的研究〔J〕.医学研究生学报,2013;26(11):1129-33.
5Fassbender JM,Saraswat-Ohri S,Myers SA,etal.Deletion of endoplasmic reticulum stress-induced CHOP protects microvasculature post-spinal cord injury〔J〕.Curr Neurovasc Res,2012;4(9):274-81.
6王春生,孟尽海,吴 燕,等.丙泊酚对大鼠急性脊髓损伤早期的影响〔J〕.宁夏医科大学学报,2013;35(9):964-7.
7姜 杰,丁 寅,周 剑,等.大鼠颈椎骨折错位致脊髓损伤模型的继发性损伤研究〔J〕.中国临床解剖学杂志,2013;31(2):165-9.
8Ohri SS,Maddie MA,Zhang Y,etal.Deletion of the pro-apoptotic endoplasmic reticulum stress response effector CHOP does not result in improved locomotor function after severe contusive spinal cord injury〔J〕.J Neurotrauma,2012;29(3):579-88.
9刘 晗,秦秉玉,邵换璋,等.神经节苷脂对大鼠脊髓损伤后内质网应激介导凋亡的抑制作用〔J〕.中华实验外科杂志,2013;30(7):1409-12.
10王德桂,马泽民,吕国华,等.金纳多对大鼠急性脊髓损伤内质网应激蛋白GADD153/CHOP表达的影响〔J〕.海南医学,2013;24(14):2032-6.
11闫 颖,赵咏梅,赵志炜,等.内质网应激通路相关分子GRP78及CHOP在糖尿病脑病小鼠海马表达的变化〔J〕.首都医科大学学报,2011;32(1):90-4.
12徐菲菲.钙网蛋白介导的内质网应激相关凋亡参与心肌细胞缺氧/复氧损伤〔D〕.中国人民解放军军医进修学院博士论文,2009.
猜你喜欢
军事文摘(2022年8期)2022-11-03
现代临床医学(2021年1期)2021-01-26
山东医药(2021年28期)2021-01-11
中华养生保健(2020年5期)2020-11-16
科学与财富(2020年15期)2020-07-04
移动通信(2020年4期)2020-05-07
移动通信(2019年4期)2019-06-25
电子制作(2019年7期)2019-04-25
现代信息科技(2018年4期)2018-07-12
中华骨与关节外科杂志(2017年1期)2017-05-17
