
贵州高原水库冬季浮游植物中汞及甲基汞分布特征*
2014-08-28 03:11李秋华高廷进闫海鱼
湖泊科学 2014年1期
李秋华,高廷进,孟 博,闫海鱼,,,, ,
(1:贵州师范大学贵州省山地环境信息系统和生态环境保护重点实验室,贵阳 550001) (2:中国科学院地球化学研究所环境地球化学国家重点实验室,贵阳 550002) (3:贵州大学喀斯特环境与地质灾害防治教育部重点实验室,贵阳 550003)
汞是一种有毒元素,普遍存在于环境污染物中[1],受汞污染的水生环境中,生物体中的高浓度汞通常要在汞污染源停止或受污染的沉积物被移除很多年后才能恢复到原来的水平[2].自水俣病以来,水生生态系统中汞污染问题特别是甲基汞污染问题受到了人们的极大关注和重视[3].甲基汞(MeHg)是一种普遍存在、具有潜在毒性的神经毒素,很容易被水生生物富集,对人类健康造成严重威胁[4-6].MeHg不同于非活性元素Hg,它可以沿着食物链生物放大[7].MeHg进入水生食物链前,必须先通过单细胞生物细胞膜,被转运到细胞内[8],主要是微藻.随之在主要以微藻为食的浮游动物中富集[9-10].浮游植物在水生食物链中是第一级,也是最关键的一级,因此污染物尤其是可富集的有毒污染物(汞、甲基汞)进入水体环境,在水生食物链中传递,先经过浮游植物的富集.MeHg在水生生物体中有很强的富集放大效应,其富集因子能达到104~107,所以即使在MeHg含量很低的水环境中,也可能在生物体中观察到高浓度的MeHg,而且水生生物体中的MeHg在总汞中所占的比例可达到85%~90%[11-13],研究浮游生物中Hg和MeHg的富集情况,对于更好地理解Hg和MeHg在食物网中的生物富集和生物放大过程有着重要意义.国内对甲基汞在水生食物链中的传递研究较少,对淡水浮游植物的富集研究更少,仅局限于鱼类等高等水生动物中甲基汞的分布研究[14-16]及以大型浮游动物为模式生物研究环境因素对其富集汞和甲基汞的影响[17-19],而缺少对食物链的第一级浮游植物的研究.本文通过对贵州省5座高原水库浮游植物中汞及甲基汞的研究,掌握汞及甲基汞在浮游植物中的富集特征和富集系数,为了解甲基汞在食物链中的传递和环境因子对汞的甲基化速率影响奠定基础,对解决汞污染水体中汞的动态变化和传递及水质管理和综合治理有重要意义.
表1 贵州高原5座水库采样点
Tab.1 Sampling sites of five reservoirs in Guizhou Plateau
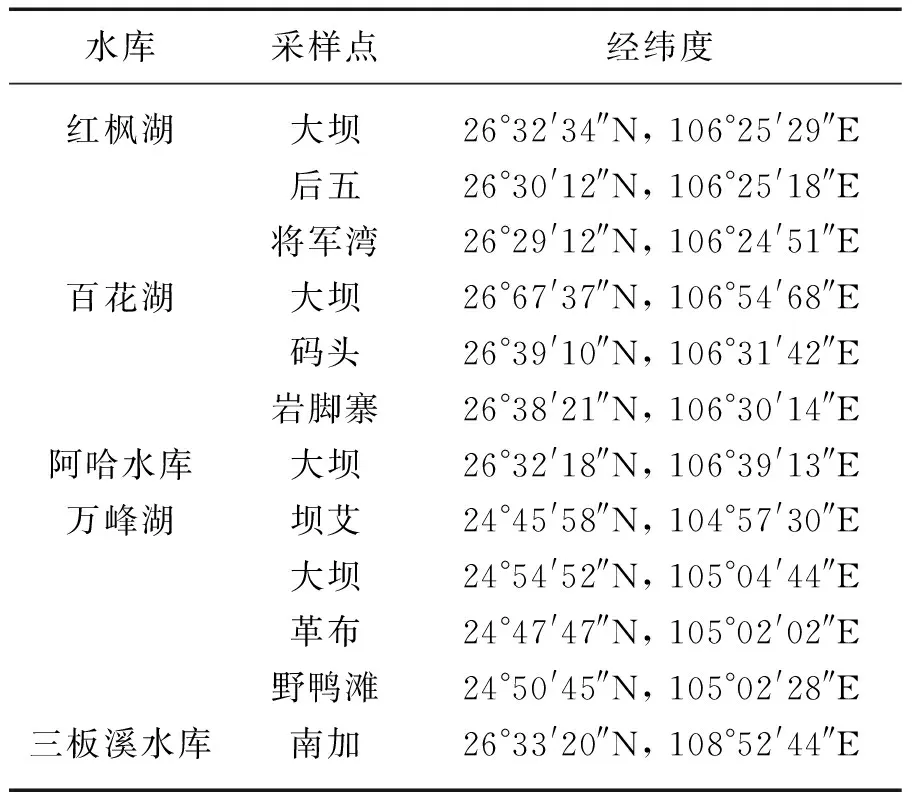
水库采样点经纬度红枫湖大坝26°32'34″N, 106°25'29″E后五26°30'12″N, 106°25'18″E将军湾26°29'12″N, 106°24'51″E百花湖大坝26°67'37″N, 106°54'68″E码头26°39'10″N, 106°31'42″E岩脚寨26°38'21″N, 106°30'14″E阿哈水库大坝26°32'18″N, 106°39'13″E万峰湖坝艾24°45'58″N, 104°57'30″E大坝24°54'52″N, 105°04'44″E革布24°47'47″N, 105°02'02″E野鸭滩24°50'45″N, 105°02'28″E三板溪水库南加26°33'20″N, 108°52'44″E
1 材料与方法
1.1 采样时间、地点及方法
于2012年12月,在贵州省境内的红枫湖(水库)、百花湖(水库)、阿哈水库、万峰湖(水库)、三板溪水库采集表层浮游植物和水样.其中红枫湖是长江水系和珠江水系分水岭处的一个人工湖,百花湖为红枫湖的下游水库.红枫湖和百花湖为大型水库,阿哈水库为中型水库.红枫湖(水库)、百花湖(水库)、阿哈水库均属于长江水系.万峰湖位于黔、滇、桂三省结合处,是国家“西电东送”的重要能源基地,珠江三角洲水质调济的重要源泉,“珠三角”经济区的重要水源供给地.三板溪水库位于沅水干流上游.采样点的设置根据水库的地形地貌设在上游、中游和下游等处,共设1~4个采样点[20-22].由于阿哈水库较小,仅在大坝处设置1个采样点.三板溪水库水质相对较好,浮游植物样品很难采集到,仅南加采样点检测到浮游植物(表1).
1.2 样品的采集
1) 用于水体中汞测定的样品:采用5L Niskin采样器在不同采样点采集表层水样,每个采样点采集3瓶(100ml)水样.采样时采样器位于采样船上游,硼硅玻璃瓶在采样前用样品水润洗3次[23],采用0.45μm(Millipore)的微孔滤膜过滤水样后装入100ml的硼硅玻璃瓶内,同时采集相应未经过滤的水样装入1000ml的硼硅玻璃瓶中,现场采集的过滤和未过滤水样均按0.5%的比例立即加入超纯盐酸溶液、放入木箱存放.样品带回实验室在4℃以下避光低温保存,并在28d内测定完毕.
2) 用于浮游植物中汞测定的样品:采用自制的25#浮游生物网,用机动船拖网采样,主要采集水体表层0~20cm的样品,带回实验室过滤并多次洗脱植物表层吸附的汞,通过显微镜镜检,样品主要是浮游植物,含有少量的小型浮游动物.过滤后的浮游植物经冷冻干燥机(LGJ-12型)冷冻干燥处理,并使用玻璃棒磨成粉末状后进行保存、测定.
1.3 样品的测定
1) 水体汞的测定:总汞和溶解态总汞采用两次金汞齐富集结合冷原子荧光法进行测定[23].在100ml水样中加入0.5ml BrCl氧化至少24h,再加入浓度为25%的NH2OH·HCl 0.2ml,反应约30min后,倒进气泡瓶中,加入200μl浓度为20%的SnCl2,用氮气将生成的Hg0富集在金汞齐上,用冷原子荧光法进行测定.
2) 水体甲基汞的测定:总甲基汞和溶解态甲基汞采用蒸馏-水相乙基化-GC-CVAFS法测定.取45ml水样在Teflon蒸馏瓶中蒸馏2.5~3.0h,收集蒸馏液,加入醋酸-醋酸钠缓冲液,将pH调节到4.9~5.3之间,加入乙基化试剂反应15min后,通纯氮载气将汞富集到Tenex管上,上机测定.测定时具体仪器参数设置参照文献[24].
3) 浮游植物总汞的测定:总汞采用硝酸水浴消解-冷原子荧光光谱法测定[25].称取0.1~0.2g(精确至0.0001g)样品于25ml比色管中;加入5ml超纯HNO3后用保鲜膜盖住比色管,在通风橱内水浴加热(95℃)3h;向比色管中加入少许超纯水和0.5ml BrCl摇匀;用超纯水定容至25ml,加盖放置24h以上,使BrCl充分将各形态Hg氧化为Hg2+;测定时加入200~400μl NH2OH·HCl去除游离态卤素,超纯水定容至25ml,摇匀,澄清后取上清液0.2~1.0ml,放入气泡瓶中进行预富集后上机测定.
4) 浮游植物甲基汞的测定:甲基汞采用溶剂萃取-水相乙基化衍生GC-CVAFS联用法测定[26].称取0.1~0.2g(精确至0.0001g)样品置于50ml离心管中;加入5ml 25% KOH溶液;将离心管置于水浴锅或烘箱内充分消解3h以上(消解温度约75~80℃);消解完成后,取出离心管冷却至室温,缓慢滴加浓HCl约3~5ml,调节溶液至酸性(pH=1~2);加入10ml二氯甲烷并称重,密封后充分振荡30min,离心(3000转/min)25min;抽掉上层废液,转移萃取液到50ml离心管内,称重,加入适量超纯水(45ml);此时溶液可放置24h以上;置于水浴锅内,水浴升温50℃,气化二氯甲烷,用竹签去除气泡,待气泡消失时,升温至80℃(不能超过),吹入N2约8min,以确保萃取液内完全没有二氯甲烷;定容至50ml,摇匀待测.

6) 浮游植物定量分析:在表层0.5m处采集1~2L水样,用福尔马林固定后的浓度为3%~5%,在实验室浓缩沉淀.浮游植物定量样品在显微镜下进行鉴定和计数,计数方法采用目镜行格法,一般随机计数2个垂直行格,对于生物量较大而不合适采用行格法的采用视野计数法,一般观察30~50个视野,使得细胞数在300个以上.
1.4 数据处理与分析
数据分析和作图采用SPSS 18软件和Excel 2003软件;浮游植物的生物富集系数(BAF)=[浮游植物的汞(甲基汞)含量(ng/g)]/[水体中溶解态汞(甲基汞)含量(ng/ml][27].
2 结果与讨论
2.1 水体中不同形态汞的分布特征
2.1.1 总汞及溶解态总汞的分布特征 贵州高原5座水库水体中总汞含量在2.45~22.56ng/L之间,以百花湖岩脚寨水体总汞含量最高,而万峰湖野鸭滩含量最低.百花湖水体总汞含量明显高于其它4座水库,而且表现出了上游岩脚寨含量最高,大坝处含量最低的特征;其它4座水库总汞含量相差不大.水体中溶解态总汞含量在1.32~3.16ng/L之间,变化幅度相对较小,但是仍以百花湖岩脚寨含量最高,而万峰湖野鸭滩含量最低.这主要是因为百花湖是一个汞污染的水库,其沉积物、水体各形态汞都较周边其它水体高[28],尽管最近5年“两湖一库”专项治理工程启动,污染源的排放得到很好的控制[29],但位于百花湖上游的岩脚寨作为离排污口最近的1个采样点,其无机汞和甲基汞含量偏高是十分正常的(图1a).
2.1.2 总甲基汞及溶解态甲基汞的分布特征 贵州高原5座水库水体中甲基汞含量在0.028~0.302ng/L之间,以阿哈水库大坝处最高,而三板溪水库南加采样点最低.除阿哈水库和百花湖水体甲基汞含量超过0.1ng/L之外,其它水库都相对较低.水体中溶解态甲基汞含量在0.021~0.090ng/L之间,表现出与水体中甲基汞一致的趋势(图1b).阿哈水库位于城市近郊,且受煤矿废水污染,接纳了大量含有汞、铁、锰及硫化物污染物,属于复合型污染水体,因此其水体中甲基汞含量也较高[30].
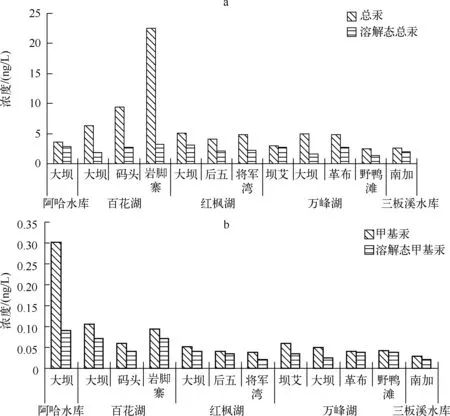
图1 5座水库水体中总汞、溶解态总汞(a)和总甲基汞、溶解态甲基汞(b)的分布特征Fig.1 Distribution characteristics of THg and DHg(a), MeHg and DMeHg(b) in water of five reservoirs
2.2 浮游植物的组成及体内汞的分布特征
2.2.1 浮游植物优势种类 冬季(12月份)贵州高原5座水库浮游植物主要是由硅藻组成,但是各水库浮游植物优势种的组成有所不同.在冬季阿哈水库和万峰湖都是以小环藻(Cyclotellasp.)为优势藻类;百花湖以颗粒直链藻极狭变种螺旋变型(Melosiragranulatavar.angustissimaf.spiralisHustedt)为优势藻;三板溪水库以颗粒直链藻(Melosiragranulata)为优势藻;而红枫湖各点差异较大,大坝和后五采样点以单角盘星藻具孔变种(Pediastrumsimplexvar.duodenarium)为优势藻,将军湾采样点却以湖泊假鱼腥藻(Pseudanabaenalimnetica)为优势藻.在冬季次优势藻以绿藻和硅藻为主,仅在红枫湖以湖泊假鱼腥藻为次优势藻(表2).
2.2.2 浮游植物丰度及组成 5座水库浮游植物丰度有明显差异,以百花湖大坝处浮游植物丰度最高,为29.05×104cells/L,而以三板溪水库南加采样点浮游植物丰度最低,仅为0.49×104cells/L.百花湖大坝、红枫湖后五和万峰湖坝艾、革布采样点浮游植物丰度相对较高,处于中营养状态;而阿哈水库大坝、百花湖码头和岩脚寨及万峰湖的大坝和三板溪水库的南加采样点浮游植物丰度都较低,处于贫营养状态(图2a).

表2 各采样点浮游植物优势种

图2 各采样点浮游植物丰度分布特征(a)和组成特征(b)Fig.2 Distribution(a) and composition(b) characteristics of phytoplankton abundance at different sampling sites
贵州高原5座水库浮游植物丰度组成百分数有显著差异,阿哈水库主要由硅藻(58.45%)和蓝藻(33.33%)组成,另外还有少量的绿藻(3.38%)和甲藻(3.38%).百花湖主要由硅藻组成,占总丰度的86.72%~94.20%,绿藻占5.80%~13.28%.红枫湖各样点间也存在显著差异,大坝、后五采样点主要由绿藻(61.94%、48.08%)和硅藻(26.22%、42.97%)及少量的蓝藻(11.84%、10.03%)组成,而在将军湾采样点主要是由蓝藻(65.82%)和绿藻(34.18%)组成.万峰湖主要由硅藻组成,占总丰度的51.73%~90.77%,以野鸭滩采样点的硅藻百分数最高;其次是绿藻(7.41%~29.21%),以大坝采样点绿藻百分数最高,在坝艾采样点还含有一定的蓝藻(30.96%).三板溪水库南加采样点主要是由硅藻(96.63%)和少量的绿藻(2.88%)组成(图2b).
2.2.3 浮游植物中总汞及甲基汞的分布特征 浮游植物中总汞和甲基汞的浓度分别在29.29~277.07ng/g和1.12~10.93ng/g之间,浮游植物中总汞浓度以百花湖岩脚寨采样点最高,三板溪水库南加采样点最低;而浮游植物中甲基汞浓度以百花湖大坝采样点最高,万峰湖大坝采样点最低.浮游植物中总汞浓度均表现出了百花湖最高,其次是阿哈水库,其它水库相差不大;而甲基汞以百花湖最高,其次是红枫湖,其它水库浮游植物中甲基汞浓度相差不大(图3a).
水体中甲基汞占总汞的百分比为0.42%~8.46%,以阿哈水库大坝采样点处最高,而可溶性的甲基汞/可溶性总汞的百分比明显要比甲基汞/总汞要低.浮游植物中甲基汞占总汞的百分比在1.27%~14.98%之间,以红枫湖最高.在甲基汞占总汞的百分比中,浮游植物的百分比明显高于水体,这也说明汞转换为甲基汞之后,更稳定,也更容易被富集(图3b).
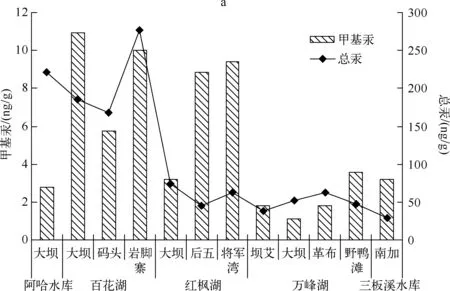

图3 浮游植物中总汞及甲基汞的分布特征(a)和水体中甲基汞占总汞的百分比(b)Fig.3 Distribution characteristics of THg and MeHg in phytoplankton(a) and the percentage of MeHg/THg in water(b)
不同水库藻类对无机汞和甲基汞吸收和吸附程度的差异说明,在不同水域由于优势藻类的种类和丰度的差异影响了其分布特征.国外研究也显示:硅藻、绿藻、蓝藻和隐藻对汞和甲基汞的富集规律是不同的[31];Moye对浮游植物富集甲基汞的机制研究中也发现,光合作用强度、辐射、光等都可以影响浮游植物对甲基汞的富集[32].Miles等在研究水体和藻类单一甲基汞分离中发现,受磷限制的藻类影响着甲基汞的水平[33].这充分说明浮游植物种类的不同对汞及甲基汞的分布影响也不同,影响浮游植物群落结构动态变化的因素都有可能影响汞及甲基汞的富集.
2.2.4 浮游植物对汞的富集系数 总汞和甲基汞在浮游植物中的生物富集系数分别在1.42×104~9.78×104和3.10×104~4.43×105之间(图4).百花湖大坝样点处总汞生物富集系数最高,其次是阿哈水库;红枫湖将军湾样点处甲基汞生物富集系数明显高于其它采样点,而以阿哈水库甲基汞的生物富集系数最低,其次是万峰湖.总汞和甲基汞在浮游植物中的生物富集系数在104~105之间,而且甲基汞的生物富集系数要明显高于总汞的生物富集系数.本研究浮游植物中总汞和甲基汞的生物富集系数与已报道的范围相近[34].

图4 浮游植物中汞及甲基汞的生物富集系数Fig.4 Bioaccumulation factors of THg and MeHg in phytoplankton
2.3 水体和浮游植物中的汞与环境因子之间的相关分析
在5座高原水库中,浮游植物中的总汞与水体中的总汞、甲基汞和溶解态甲基汞(r总汞=0.75,r甲基汞=0.64,r溶解态甲基汞=0.84,n=12,P<0.01)都呈极显著正相关,说明水中较高的无机汞和甲基汞会吸附在浮游植物体上,其原理可能是通过浮游植物胞外分泌物吸附[35].浮游植物中的甲基汞与水体中的总汞及甲基汞均不存在显著相关性(P>0.05),这说明甲基汞可能是通过食物链进入水生食物链,而不是主要以吸附作用进入浮游植物.浮游植物中的甲基汞与水体富营养化的环境因子——透明度、总氮、硝态氮(r透明度=-0.68,r总氮=-0.59,r硝态氮=-0.65,n=12,P<0.05)都呈显著负相关;浮游植物中的总汞与水体中的电导率、叶绿素a(r电导率=0.60,r叶绿素a=0.66,n=12,P<0.05)都呈显著正相关,而与透明度(r=-0.65,n=12,P<0.05)呈显著负相关(表3),这也表明水体富营养化程度的不同对浮游植物中的甲基汞有影响,富营养化程度的增加能显著降低浮游植物中甲基汞的浓度,浮游植物的快速生长减弱了浮游植物对汞的吸收、吸附和汞的甲基化.
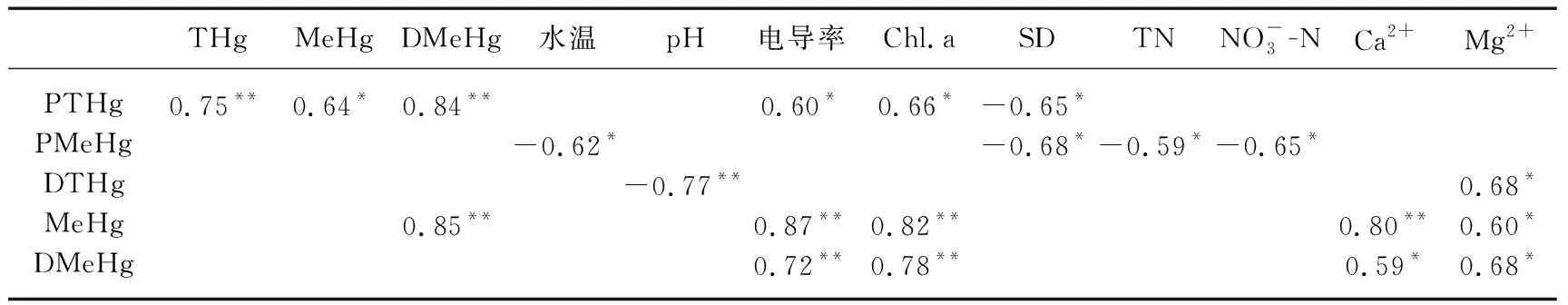
表3 水体和浮游植物中总汞、甲基汞与环境因子之间的相关性(n=12)1)
1) PTHg:浮游植物中总汞;PMeHg:浮游植物中甲基汞;DTHg:水体溶解态总汞;MeHg:水体甲基汞;DMeHg:水体溶解态甲基汞;** 表示在0.01水平(双侧)上呈极显著相关;* 表示在0.05水平(双侧)上呈显著相关.
3 结论
1) 5座高原水库中,百花湖浮游植物中的总汞和甲基汞含量最高,分别为277.07和10.93ng/g,远远高于其它水库;总汞和甲基汞在浮游植物中的生物富集系数分别在1.42×104~9.78×104和3.10×104~4.43×105之间.
2) 浮游植物中的总汞与水体中的总汞和甲基汞均呈极显著正相关,而浮游植物中的甲基汞与水体中的总汞和甲基汞相关性不显著.
[1] Back RC, Watras CJ. Mercury in zooplankton of northern Wisconsin lakes: taxonomic and site-specific trends.Water,Air,andSoilPollution, 1995,80:931-938.
[2] Kim JP, Burggraaf SC. Mercury bioaccumulation in rainbow trout (Oncorhynchusmykiss) and the trout food web in lake Okareka, Okaro, Tarawera, Rotomahana and Rotorua, New Zealand.Water,Air,andSoilPollution, 1999,115: 535-546.
[3] 冯新斌,仇广乐,付学吾等.环境汞污染.化学进展,2009,21(2/3):436-456.
[4] Bernard S, Enayati A, Redwood Letal. Autism: a novel form of mercury poisoning.MedicalHypotheses, 2001,56: 462-471.
[5] Castoldi AF, Coccini T, Ceccatelli Setal. Neurotoxicity and molecular effects of methylmercury.BrainResearchBulletin, 2001,55: 197-203.
[6] Yoshizawa K, Rimm EB, Morris JSetal. Mercury and the risk of coronary heart disease in men.NewEnglandJournalofMedicine, 2002,347: 1755-1760.
[7] Wiener JG, Krabbenhoft DP, Heinz GHetal. Ecotoxicology of mercury. In: Hoffman DJ, Rattner BA, Burton GAJetaleds. Handbook of ecotoxicology. Boca Raton, FL: CRC Press, 2003: 409-463.
[8] Morel FMM, Kraepiel AMLetal. The chemical cycle and bioaccumulation of mercury.AnnualReviewofEcologyandSystematics, 1998,29: 543-566.
[9] Kainz M, Mazumder A. Effect of algal and bacterial diet on methyl mercury concentrations in zooplankton.EnvironSciandTechnol, 2005,39: 1666-1672.
[10] Tremblay A. Bioaccumulation of methylmercury in invertebrates from boreal hydroelectric reservoirs. In: Lucotte M, Schetagne R, Théríen Netaleds. Mercury in the biogeochemical cycle: Natural environment and hydroelectric reservoirs of Northern Québec. Berlin: Springer, 1999: 193-214.
[11] Watras CJ, Bloom NS. Mercury and methylmercury in individual zooplankton-implications for bioaccumulation.LimnolOceanogr, 1992,37:1313-1318.
[12] Watras CJ, Back RC, Halvorsen Setal. Bioaccumulation of mercury in pelagic freshwater food webs.ScioftheTotalEnviron, 1998,219: 183-208.
[13] Chen CY, Stemberger RS, Klaue Betal. Accumulation of heavy metals in food web components across a gradient of lakes.LimnolOceanogr,2000,45:1525-1536.
[14] 闫海鱼,冯新斌,刘 霆等.贵州百花湖鱼体汞污染现状.生态学杂志,2008,27(8):1357-1361.
[15] 何天容,冯新斌.红枫湖出入库河流汞浓度分布特征及影响因素分析.环境科学与技术,2010,33(7):138-141.
[16] 姚 珩,冯新斌,闫海鱼等.乌江洪家渡水库鱼体汞含量.生态学杂志,2010,29(6):1155-1160.
[17] Tsui MTK, Wang WX. Temperature influences on the accumulation and elimination of mercury in a freshwater cladoceran,Daphniamagna.AquaticToxicology,2004,70: 245-256.
[18] Tsui MTK, Wang WX. Influences of maternal exposure on the tolerance and physiological performance ofDaphniamagnaunder mercury stress.EnvironmentalToxicologyandChemistry, 2005,24(5):1228-1234.
[19] Tsui MTK, Wang WX. Acute toxicity of mercury toDaphniamagnaunder different conditions.EnvironSciandTechnol, 2006,40:4025-4030.
[20] 李秋华,商立海,李广辉等.万峰湖浮游植物群落的时空分布.生态学杂志,2011,30(5):1031-1038.
[21] 李秋华,陈丽丽,夏品华等.贵州百花湖麦西河河口浮游植物群落结构及其与环境因子之间关系.湖泊科学,2011,23(4):523-529.
[22] 商立海,李秋华,邱华北等.贵州红枫湖水体叶绿素a的分布与磷循环.生态学杂志,2011,30(5):1023-1030.
[23] 闫海鱼,冯新斌,汤顺林等.天然水体中痕量不同形态汞的分析方法的建立.分析测试学报,2003,22(5):10-13.
[24] 蒋红梅,冯新斌,梁 琏等.蒸馏-乙基化GC-CVAFS法测定天然水体中的甲基汞.中国环境科学,2004,24(5):568-571.
[25] 郑 伟,冯新斌,李仲根等.硝酸水浴消解-冷原子荧光光谱法测定植物中的总汞.矿物岩石地球化学通报,2006,25(3):285-287.
[26] 仇广乐,何天容,王少峰等.溶剂萃取-水相乙基化衍生GC-CVAFS联用测定苔藓样品中的甲基汞.分析测试学报,2005,24(1):29-32.
[27] Pichet P, Morrison K, Rheault Ietal. Analysis of total mercury and methylmercury in environmental samples. In: Lucotte M, Schetagne R, Théríen Netaleds. Mercury in the biogeochemical cycle: Natural environment and hydroelectric reservoirs of Northern Québec. Berlin: Springer Verlag, 1999: 41-54.
[28] 王翠萍.污染源控制对历史性汞污染湖泊中汞形态分布的影响——以百花水库为例[学位论文].贵阳:贵州大学,2011.
[29] U.S. EPA. Mercury Study Report to the Congress, EPA 452yR-97-0003, EPA, USA, December 1997.
[30] Yan HY, Feng XB, Shang LHetal. The variations of mercury in sediment profiles from a historically mercury-contaminated reservoir, Guizhou province,China.ScioftheTotalEnviron, 2008,407(1): 497-506.
[31] Liu B, Yan HY, Wang CPetal. Insights into low fish mercury bioaccumulation in a mercury-contaminated reservoir, Guizhou, China.EnvironmentalPollution, 2012,160: 109-117.
[32] Feng XB, Bai WY, Shang LHetal. Mercury speciation and distribution in Aha Reservoir which was contaminated by coal mining activities in Guiyang, Guizhou, China.AppliedGeochemistry, 2011,26(2): 213-221.
[33] Pickhardt PC, Fisher NS. Accumulation of inorganic and methylmercury by freshwater phytoplankton in two contrasting water bodies.EnvironSciandTechnol, 2007,41:125-131.
[34] Moye HA, Miles CJ, Phlips EJetal. Kinetics and uptake mechanisms for monomethylmercury between freshwater algae and water.EnvironSciandTechnol, 2002,36: 3550-3555.
[35] Miles CJ, Moye HA, Phlips EJetal. Partitioning of monomethylmercury between freshwater algae and water.EnvironSciandTechnol, 2001,35: 4277-4282.
猜你喜欢
福建农业学报(2021年9期)2022-01-05
快乐作文(3.4年级)(2020年4期)2020-05-25
人物画报(2020年6期)2020-01-06
北方音乐(2018年16期)2018-05-14
百科知识(2018年6期)2018-04-03
海洋开发与管理(2017年9期)2017-12-05
分析化学(2014年10期)2014-10-24
中国环境科学(2014年2期)2014-04-26
中国三峡(2013年11期)2013-11-21
少年文艺·我爱写作文(2009年4期)2009-05-11
