
不同树龄茶园杂草群落物种组成及多样性差异
2014-08-13 08:41:28孙国俊朱叶芹张海艳李粉华
杂草学报 2014年1期
季 敏,孙国俊,2,朱叶芹,张海艳,李粉华,韩 敏,袁 方
(1.江苏省金坛市植保植检站,江苏金坛 213200;2.扬州大学园艺与植物保护学院,江苏扬州 225009;3.江苏省植物保护站,江苏南京 210036)
杂草特别是农田杂草是一类在人为与自然选择双重压力下产生的高度进化的植物类群。与作物相比,杂草的抗逆境能力强,并且经过长期的自然进化和人工选择,具有广泛的适应性和顽强的生命力。现有杂草在农田中的分布状况是杂草在人工和自然双重因素作用下产生的,不同农田环境中的杂草分布数据包含了它们对环境的适应和种内种间竞争的信息,应用数学方法对其进行解析并赋予生态学意义的解释,能够揭示不同杂草对生态要求的差异性,对单一种杂草的生态学特性进行全面描述比较困难,但是通过种间比较可以把握它们的生态学特性差异。
茶园杂草种类繁多,发生危害时较严重,尤其是幼龄茶园的地表空间较大,更易孳生杂草。杂草与茶树争水、争肥、争空间、争阳光,严重影响了茶园茶叶的产量和品质,许多杂草还是茶树病虫害如叶蝉类、蚜虫、螨类的中间寄主[1]。茶园杂草作为茶园生态系统中生物多样性的组成成分,能够改变茶园生态系统的结构,从而直接影响茶园昆虫、微生物等物种的多样性和数量配制。
茶树是多年生常绿灌丛,且茶园远比一般农田生态系统稳定,其杂草群落组成复杂、稳定性强。目前国内外已经开展了很多关于不同农艺管理措施对农田杂草群落组成及生物多样性影响[1-3]的研究,但是关于茶树种植时间的长短对茶园杂草群落影响的研究尚未见报道。李粉华等研究了不同树龄茶园春、秋季杂草的发生危害,但此类研究仅就调查数据做了定性分析,没有进行深入的多样性定量分析和研究[4-5]。群落物种多样性和分布格局一直是群落生态学研究中的重点和热点[6],植物群落多样性分布格局受环境因素(作物、水分、土壤、人类活动等)的影响较大[7]。为了研究幼龄、成年茶园杂草群落多样性差异,笔者于2011—2012年调查了江苏省金坛市茶园杂草的种类、数量、高度等,发现茶树种植时间的长短对茶园杂草的发生数量、群落组成、群落多样性等都有显著影响。本研究试图从生态学角度分析不同种植时间茶园杂草的种类、密度、优势杂草组成、杂草生物多样性,以期为茶园杂草的综合治理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区金坛市位于31°33′42″~31°53′22″ N,119°17′45″~119°44′59″ E,地处江苏省南部的长江下游。茶叶生产面积2 733 hm2,主要分布在茅山丘陵地带,海拔10~50 m,土质以黄棕壤土、基性岩土、石灰岩土为主。常年年均气温15.3 ℃,年均降雨量1 063.6 mm,雨水多集中在春、夏、秋季,常年年均日照时数为2 035.5 h[8]。
1.2 杂草生物多样性的调查
分别于2011、2012年的4、10月,在冬春、夏秋季多数杂草已开花结实时,根据茶叶生产布局选择有代表性的茶园,2年内在春、秋季分别调查60块、64块茶园。春季调查的60块茶园按不同种植时间进行分类,种植10年以上的成年茶园有36块,种植6年以下的幼龄茶园有24块;秋季调查的64块茶园中种植10年以上的成年茶园有44块,种植6年以下的幼龄茶园有20块。考虑到杂草生长与茶树的竞争性,且成年茶园茶树的生长时间长、覆盖度大、空行小,绝大多数杂草生长在茶间空行,因此调查的样方不宜过大。每块茶园采用“M”形9点取样法进行调查[9-10],每个样方0.25 m2(0.5 m×0.5 m),分别记载杂草种类、数量、高度,并详细记载调查地点、茶叶品种、种植时间、土质、土壤类型、坡度、坡向、农事管理措施等。
1.3 不同树龄茶园生境的差异性分析
幼龄茶园的主要生境特征:茶叶品种主要为无性系良种茶,田间茶树覆盖度小,空行大,管理较为精细,耕翻频繁,施肥频繁且多,除草剂使用频繁,多数田块土壤肥沃、疏松,但有少部分基质岩土,土壤较贫瘠。
成年茶园的主要生境特征:田间茶树覆盖度大,空行小,管理较为松散,只有部分茶园耕翻,施肥不多,除草剂使用次数较少,有部分茶园常年不使用除草剂,偏于荒废,土壤肥沃、疏松,部分茶园剪枝铺行。
比较可知,2种茶园生境的差异主要表现在茶树生长时间、田间覆盖度大小、人工管理措施等方面。
1.4 数据分析方法
按下列公式计算杂草相对多度(relative abundance,RA):
RA=RU+RD+RF
式中:RU、RD、RF分别为某杂草的相对均度(%)、相对密度(%)、相对频度(%)。
田间均度是指某杂草在田块中出现的样方数占调查田块总样方数的百分比,田间频度是指某杂草出现的田块数占总调查田块数的百分比,田间密度是指某杂草在各调查田块的平均密度和与田块数之比,相对均度是指某杂草均度与各种杂草均度和之比,相对密度是指某杂草平均密度与各种杂草密度和的比,相对频度是指某杂草频度与各种杂草频度和的比。由于相对多度综合了以上各调查数据及统计数据,因此相对多度较大的杂草将被认为是当地的主要优势杂草,可以体现杂草的丰富程度[11-12]。
物种丰富度S,即样方中包含的所有杂草种类数;物种多样性H′用Shannon-Wiener指数测度,公式为:
H′=∑PilnPi
式中:Pi=Ni/N,Ni为样方中第i物种的个体数,N为样方总个体数。
群落优势度D用Simpson指数测度,公式为:
群落均匀度J用Pielou均匀度指数测度,公式为:
J=H’/lnS[13-14]
上述公式中i的取值范围为1~S。
群落相似性[15]:用Sørenson指数对群落物种组成进行定性分析。
Cs=2j/(a+b)。
式中:Cs为不同类群杂草物种的相似性指数,j为2个群落共有的类群数,a、b分别为群落A和群落B的类群数。
用Morisita-Horn指数对群落的物种组成及其个体数进行定量分析
CMH=2∑(aibi)/(da+db)NaNb
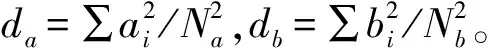
2 结果与分析
2.1 不同树龄茶园杂草的群落组成分析
2.1.1 不同树龄茶园春季杂草群落的物种组成分析 对2年春季的调查结果(表1)进行分析可知,成年茶园有22科38属38种杂草,幼龄茶园只有14科24属24种杂草,成年茶园比幼龄茶园分别多8科14属14种杂草。从杂草的科属种构成来看,幼龄茶园能发现的杂草在成年茶园几乎都有生长,共有的杂草有14科22属22种。成年茶园比幼龄茶园多的杂草科种类分别为蔷薇科、忍冬科、莎草科、报春花科、唇形科、凤尾蕨科、海金沙科、夹竹桃科、堇菜科,而幼龄茶园的十字花科杂草仅比成年茶园多1属1种。
由表1可以看出,成年茶园春季杂草优势科为菊科、禾本科,其杂草种类占总种类的比例分别达18.42%、10.53%;其次为蔷薇科、十字花科、石竹科、玄参科、百合科、豆科杂草,其杂草种类都分别占总种类的5%以上。幼龄茶园春季杂草优势科与成年茶园有较大区别,菊科杂草在幼龄茶园春季杂草的属、种数量中占绝对优势,其种类占杂草总种类的25.00%;其次是十字花科杂草,种类占杂草总种类的12.50%;再次是禾本科、石竹科、玄参科杂草,种类占杂草总种类的8.33%。
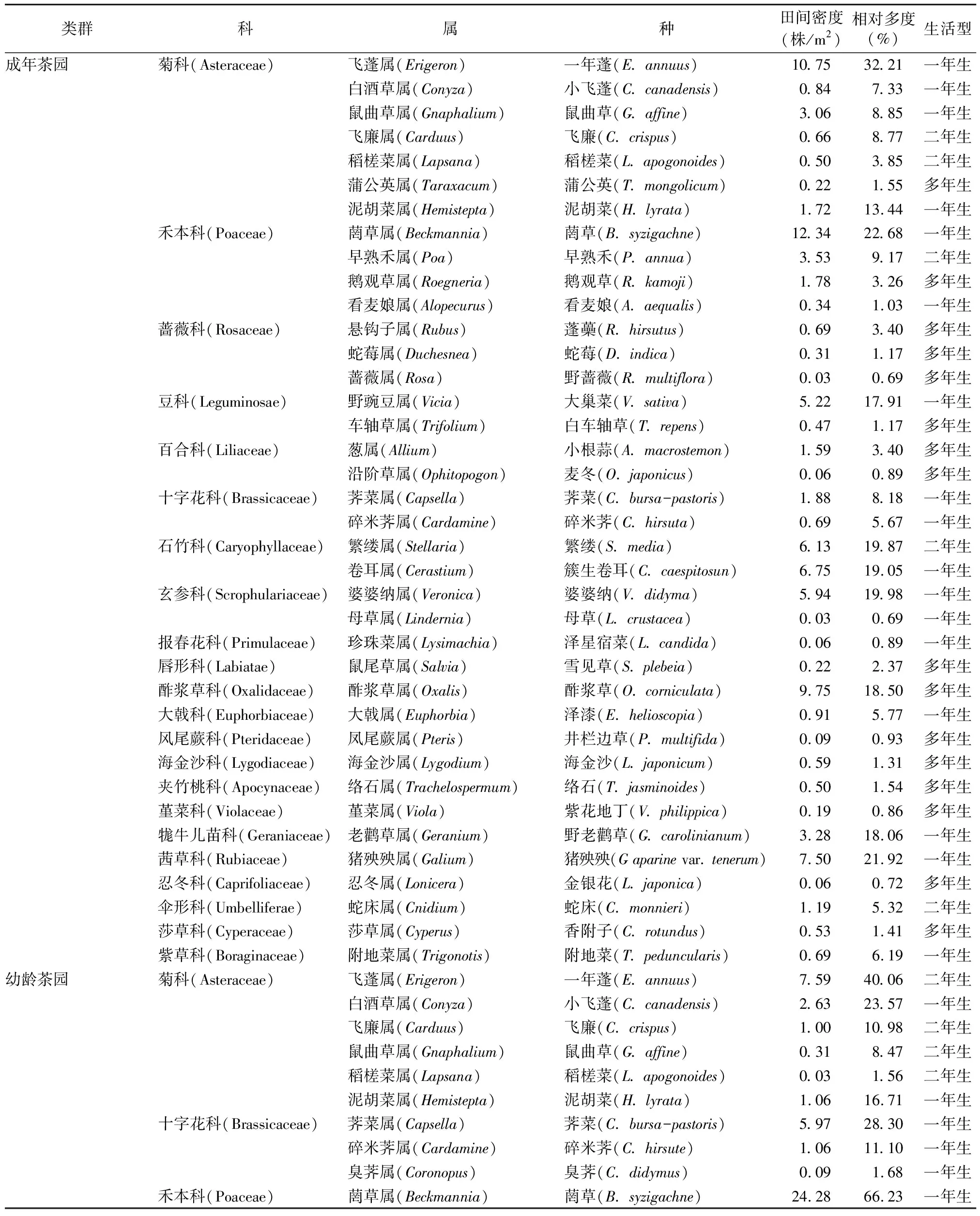
表1 不同树龄茶园春季杂草群落物种组成、生活型、数量特征

续表1(Continued)
2.1.2 不同树龄茶园秋季杂草群落物种组成分析 2年秋季茶园的调查结果(表2)表明,成年茶园有41科81属94种杂草,幼龄茶园只有24科47属53种杂草。从杂草的科属种来看,幼龄茶园能见的杂草,在成年茶园几乎都有生长,共有的杂草有22科41属47种。成年茶园比幼龄茶园多的杂草科分别为蔷薇科、百合科、车前科、凤尾蕨科、海金沙科、夹竹桃科、堇菜科、蕨科、藜科、里白科、柳叶菜科、茜草科、商陆科、天南星科、紫草科、桑科,而幼龄茶园杂草大部分都可在成年茶园中找到。
由表2可以看出,成年茶园秋季的优势杂草科为菊科、禾本科,其杂草种类占成年茶园秋季杂草总种类的10%以上,分别达15.96%、13.83%;其次为大戟科杂草,其杂草种类占成年茶园秋季杂草总种类的5%以上。菊科、禾本科杂草在幼龄茶园秋季杂草的属、种数量中仍占绝对优势,其种类分别占幼龄茶园秋季杂草总种类的18.87%、16.98%;其次是大戟科、莎草科、苋科杂草,其种类分别占幼龄茶园秋季杂草总种类的7.55%、5.66%、5.66%。秋季幼龄茶园与成年茶园在杂草的科、属方面的区别主要是由于次优势杂草科、属、种发生了变化。

表2 不同树龄茶园秋季杂草群落物种组成、生活型、数量特征
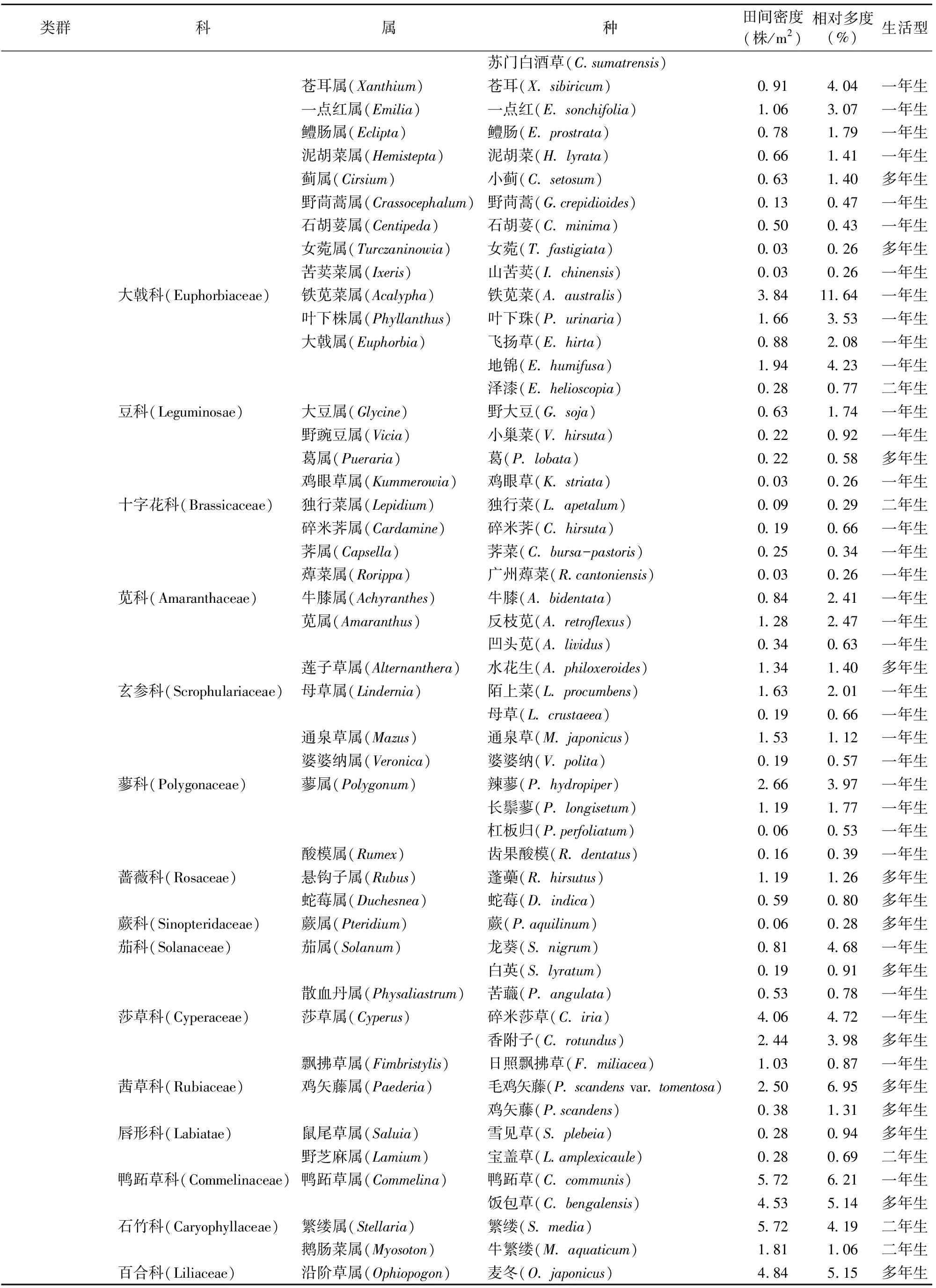
续表2(Continued)
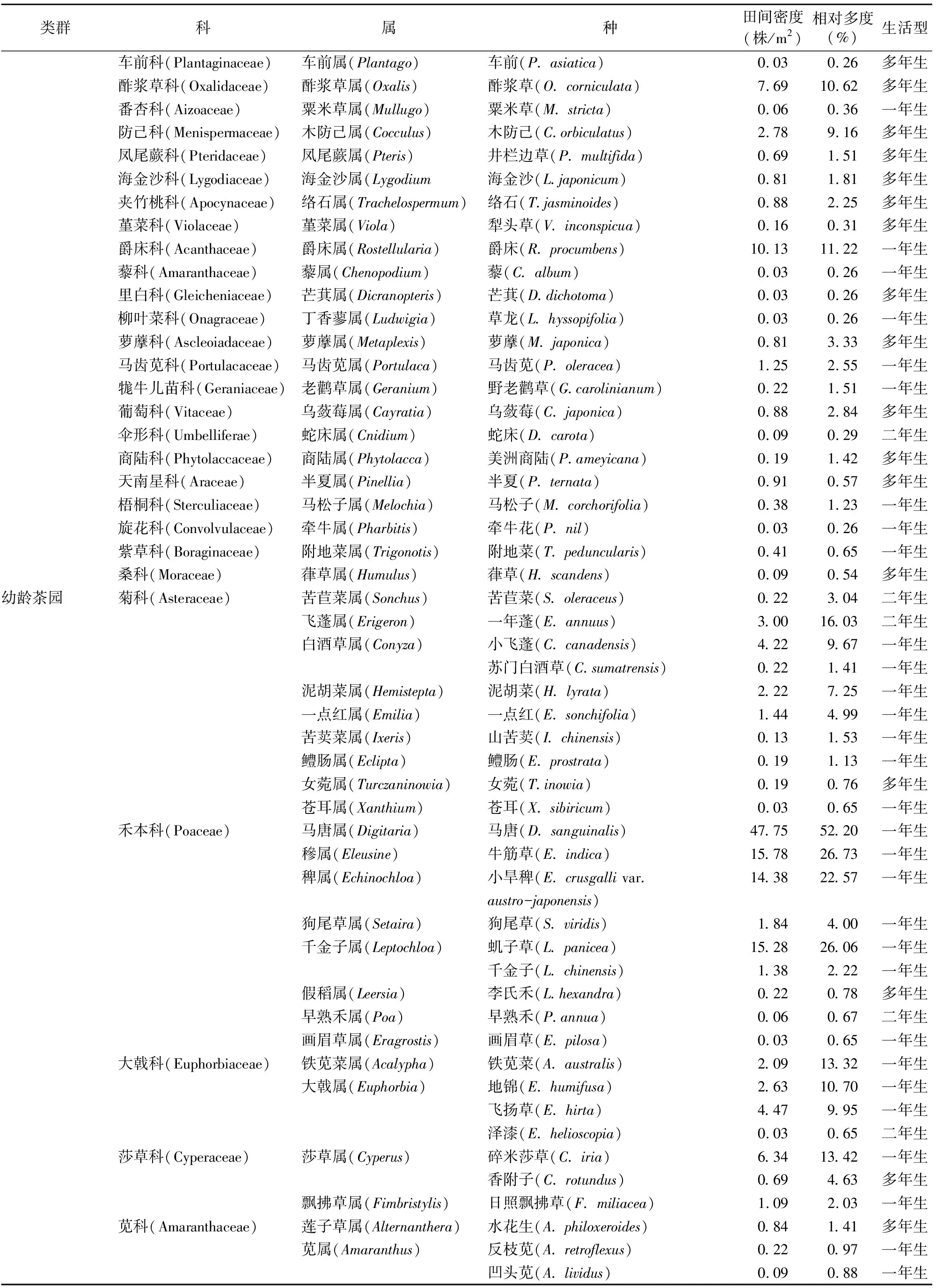
续表2(Continued)

续表2(Continued)
2.2 不同树龄茶园杂草群落生活型的组成分析
2.2.1 不同树龄茶园春季杂草生活型的组成分析 由图1的调查结果看出,成年茶园春季38种杂草中有多年生杂草17种,占成年茶园杂草总种类的44.74%;一年生和二年生短命杂草21种,占成年茶园杂草总种类的55.26%。幼龄茶园春季24种杂草中多年生杂草只有2种,占幼龄茶园杂草总种类的8.33%;一年生和二年生短命杂草22种,占幼龄茶园杂草总种类的91.67%。可以看出,成年茶园多年生杂草的比例明显高于幼龄茶园。
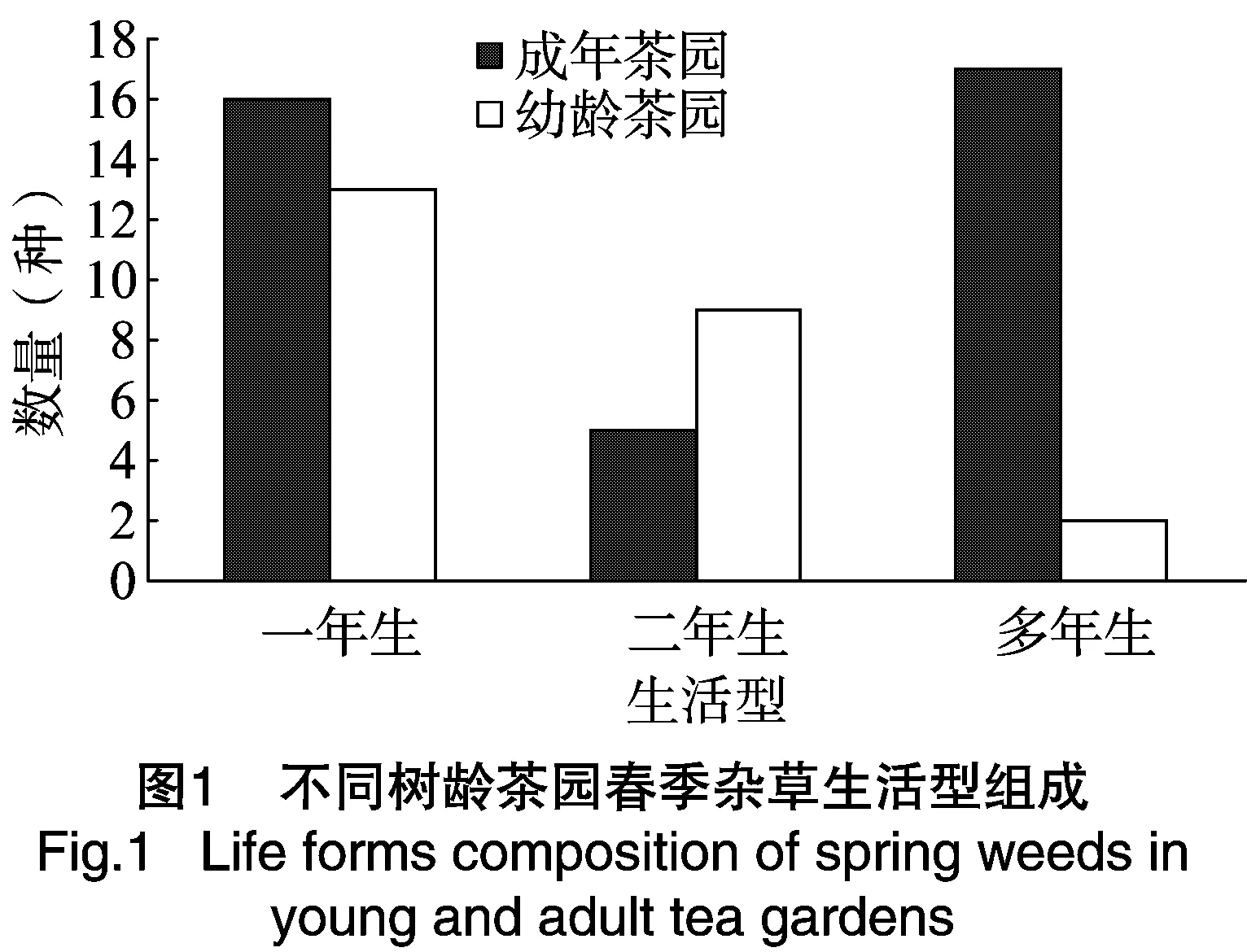
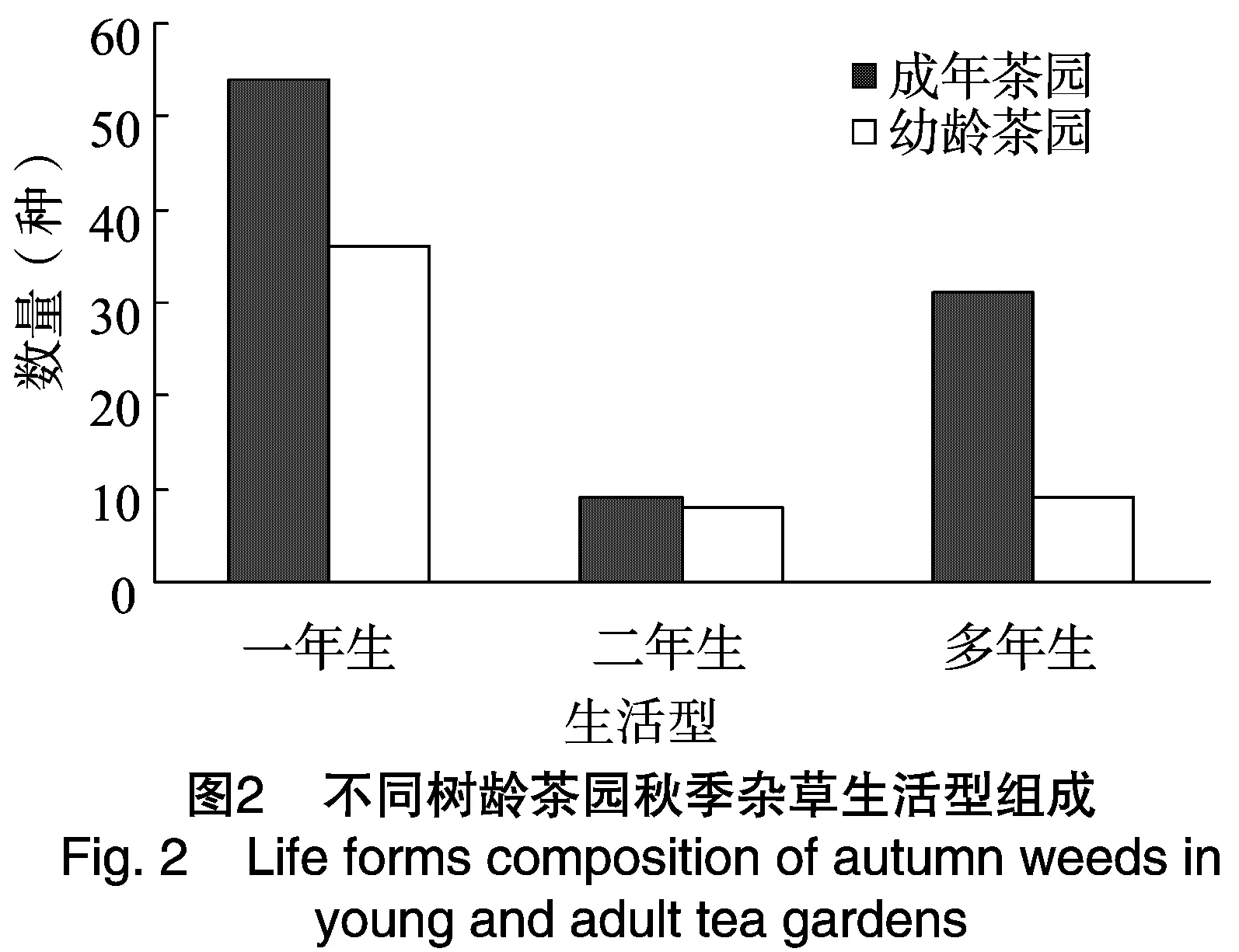
2.2.2 不同树龄茶园秋季杂草生活型组成分析 由图2的调查结果看出,成年茶园秋季94种杂草中有多年生杂草31种,占成年茶园杂草总种类的32.98%;一年生和二年生短命杂草63种,占成年茶园杂草总种类的67.02%。幼龄茶园秋季53种杂草中多年生杂草只有9种,占幼龄茶园杂草总种类的16.98%;一年生和二年生短命杂草44种,占幼龄茶园杂草总种类的83.02%。可以看出,成年茶园的秋季多年生杂草比例高于幼龄茶园,秋季茶园的多年生杂草种类明显多于春季。
2.3 不同树龄茶园杂草群落的数量特征分析
2.3.1 不同树龄茶园春季杂草群落的数量特征分析 调查结果显示,成年茶园中相对多度最大的杂草为一年蓬,其相对多度为32.21%,田间密度为10.75株/m2;相对多度≥20%且<30%的有2种杂草,分别为菵草、猪殃殃,其相对多度分别为22.68%、21.92%,田间密度分别为12.34、7.50株/m2;相对多度≥10%且<20%的有7种杂草,分别为婆婆纳、繁缕、簇生卷耳、酢浆草、野老鹳草、大巢菜、泥胡菜,其相对多度分别为19.98%、19.87%、19.05%、18.50%、18.06%、17.91%、13.44%,田间密度分别为5.94、6.13、6.75、9.75、3.28、5.22、1.72株/m2;相对多度≥5%且<10%的有9种杂草,分别为早熟禾、鼠曲草、飞廉、荠菜、小飞蓬、附地菜、泽漆、碎米荠、蛇床;其余19种杂草的相对多度<5%。
幼龄茶园相对多度最大的杂草为菵草,其相对多度为66.23%,田间密度为24.28株/m2;相对多度≥40%且<50%的杂草为一年蓬,其相对多度为40.06%,田间密度为7.59株/m2;相对多度≥20%且<30%的有2种杂草,为荠菜、小飞蓬,其相对多度分别为28.30%、23.57%,田间密度分别为5.97、2.63株/m2;相对多度≥10%且<20%的有4种杂草,分别为猪殃殃、泥胡菜、碎米荠、飞廉,其相对多度分别为19.16%、16.71%、11.10%、10.98%,田间密度分别为3.09、1.06、1.06、1.00株/m2;繁缕、酢浆草、野老鹳草、鼠曲草、附地菜、早熟禾、宝盖草、卷耳、婆婆纳、大巢菜等10种杂草的相对多度均≥5%且<10%,只有6种杂草的相对多度<5%。可以看出,不同树龄茶园杂草群落的优势杂草种类、数量区别较大。
2.3.2 不同树龄茶园秋季杂草群落数量的特征分析 成年茶园中相对多度最大的杂草为马唐,其相对多度为64.07%,田间密度为131.91株/m2;相对多度≥10%且<20%的有7种杂草,分别为牛筋草、一年蓬、小旱稗、铁苋菜、小飞蓬、爵床、酢浆草,其相对多度分别为15.03%、13.78%、12.38%、11.64%、11.59%、11.22%、10.62%,田间密度分别为19.56、7.09、12.94、3.84、5.38、10.13、7.69株/m2;相对多度≥5%且<10%的有6种杂草,分别为木防己、毛鸡矢藤、鸭跖草、虮子草、麦冬、饭包草,其余80种杂草的相对多度<5%。
幼龄茶园相对多度最大的杂草仍然为马唐,其相对多度为52.20%,田间密度为47.75株/m2;相对多度≥20%且<30%的有3种杂草,分别为牛筋草、虮子草、小旱稗,其相对多度分别为26.73%、26.06%、22.57%,田间密度分别为15.78、15.28、14.38株/m2;相对多度≥10%且<20%的有5种杂草,分别为一年蓬、碎米莎草、铁苋菜、马齿苋、地锦,其相对多度分别为16.03%、13.42%、13.32%、11.36%、10.70%,田间密度分别为3.00、6.34、2.09、2.41、2.63株/m2;相对多度≥5%且<10%的有6种杂草,分别为飞扬草、小飞蓬、泥胡菜、通泉草、粟米草、酢浆草;只有38种杂草的相对多度<5%。可以看出,除相对多度最大的杂草种类相同外,不同树龄茶园秋季杂草群落的优势杂草种类、数量区别较大。
2.4 不同树龄茶园杂草群落的生物多样性分析
2.4.1 不同树龄茶园春季杂草的生物多样性分析 表3的调查结果表明,成年茶园春季杂草物种丰富度为38,比幼龄茶园大;成年茶园春季杂草物种多样性Shannon-Wiener指数明显比幼龄茶园大;成年茶园春季杂草群落优势度Simpson指数明显比幼龄茶园小;成年茶园春季杂草群落Pielou均匀度指数略大于幼龄茶园。茶园杂草物种的丰富度、多样性、均匀度与优势度是从不同角度衡量物种群落稳定性的重要指标,茶园杂草物种丰富度、群落物种的多样性参数值与均匀度越大,优势度越小,则物种群落的结构越复杂,其反馈系统也就越强大,对于环境的变化或来自物种群落内部种群波动的缓冲作用越强,物种群落也就越稳定,这样的杂草群落优势种不太突出,杂草也就不容易严重发生[3]。
群落相似性系数(Sørenson指数为0.74,Morisita-Horn指数为0.34)表明,不同树龄茶园春季杂草群落组成的差异主要表现为杂草田间密度的差异。
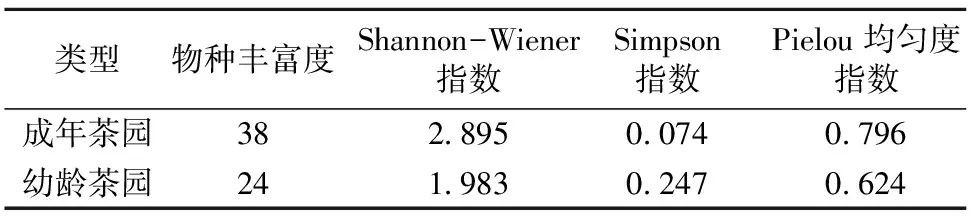
表3 不同树龄茶园春季杂草生物多样性分析结果
2.4.2 不同树龄茶园秋季杂草生物多样性分析 表4的调查结果表明,成年茶园秋季杂草物种丰富度为94,比幼龄茶园大;成年茶园秋季杂草物种多样性Shannon-Wiener指数大于幼龄茶园;成年茶园秋季杂草群落优势度Simpson指数比幼龄茶园大;成年茶园秋季杂草群落Pielou均匀度指数略小于幼龄茶园。
群落相似性系数(Sørenson指数为0.71,Morisita-Horn指数为0.06)表明,不同树龄茶园秋季杂草群落组成的差异主要表现为杂草田间密度的差异,且差异极大。
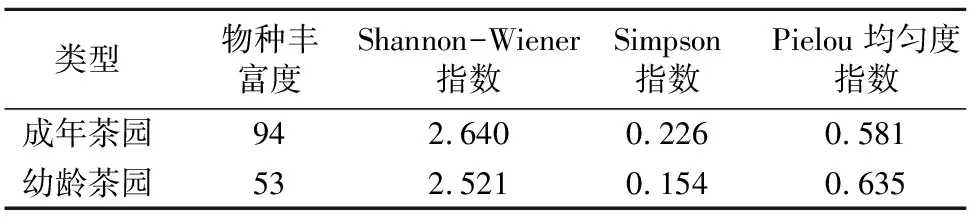
表4 不同树龄茶园秋季杂草生物多样性分析结果
3 结论与讨论
物种多样性反映了生物群落在组成、结构、功能和动态方面的异质性,体现了群落结构类型、组织水平、发展阶段、稳定程度和生境的差异[16],本研究表明,茶树林龄对茶园杂草多样性具有显著的影响。成年茶园春季杂草有38种,分属22科38属;幼龄茶园春季杂草有24种,分属14科24属。成年茶园秋季杂草有94种,分属41科81属;幼龄茶园秋季杂草有53种,分属24科47属。可见成年茶园杂草种类多,物种丰富,且成年茶园多年生杂草比例明显高于幼龄茶园。随着茶树栽种年限的延长,虽然茶树的覆盖率逐渐增高,但是茶园生态系统渐趋稳定,人为扰动偏少,有利于多种杂草的适生繁殖,也有利于多年生杂草的定植生存。人们通过对植物物种共存机制的研究认为,物种形成过程和物种迁徙特性在大尺度上决定了一个地区潜在共存的物种数量,但这些物种是否出现还取决于小尺度水平上群落内过程的作用[17]。研究地位于相近的地理位置,在大尺度上应存在相同数量的潜在共存物种。调查结果显示,不同树龄茶园杂草的物种种类及数量差异较大,这只能是小尺度水平上群落内过程作用导致的结果。对于小尺度的植物群落而言,群落生境的差异可能是形成多样性的主要原因[18]。
物种被认为是能最直接、最易观察和最适合研究生物多样性的生命层次类群[19],物种多样性的研究是生物多样性研究的基础环节,是保护生物学研究的基础,可为植物资源的开发、利用和科学研究提供最基础的数据和资料。周德明等对湖南地区4个林龄阶段的杉木人工林林下植被组成及多样性进行了研究,结果显示,林龄对杉木林下植被的丰富度指数影响不大,但优势种的种类有所差异[20]。Harms等研究表明,并不是所有的物种均与生境类型相关,也就是说生境类型并不能完全解释物种多样性的差异,其他方面的因素同样制约着林下植被的多样性[21]。不同树龄茶园杂草物种组成的研究结果表明,成年茶园春季主要优势科、属杂草为菊科、禾本科,幼龄茶园春季主要优势杂草为菊科。幼龄茶园春季优势科、属杂草与成年茶园春季优势科、属杂草区别较大的主要原因是冬、春季农田适宜生长的禾本科杂草总种类偏少;菊科、禾本科杂草同为幼龄、成年茶园秋季杂草的主要优势科、属杂草,2种生境优势科、属杂草的相同充分说明菊科、禾本科杂草为农田秋季的广布杂草种类。幼龄、成年茶园次优势科、属、种杂草的差异是杂草在茶园生态系统中适应性变化的结果。生境的变化导致群落物种的变化,使不同生活、生态特性的物种在不同群落环境中成为优势种、次优势种和群落生态功能的维持者[22],长期管理措施的差异会导致田间杂草群落物种多样性的巨大差异[2、23],本研究结果也再一次验证了这一结论。
杂草群落数量特征分析结果表明,由于幼龄、成年茶园杂草群落种类数量的差异,幼龄茶园优势杂草种类的比例高,其优势度明显;茶园春季杂草群落中阔叶杂草RA值大的种类多,茶园秋季杂草群落中禾本科杂草RA值大的种类多,禾本科杂草群落优势度高。幼龄茶园秋季杂草群落中禾本科杂草占绝对优势,成年茶园秋季杂草群落的主要优势杂草种类中有部分多年生藤本杂草。
植物多样性会受到各种环境因素的影响[24-25],随着树龄的增加,茶树个体对资源的需求也将会有所增加。研究结果表明,成年茶园春季杂草物种丰富度和多样性指数均高于幼龄茶园,成年茶园春季杂草均匀度指数高于幼龄茶园,成年茶园春季杂草优势度指数明显低于幼龄茶园,由此说明,幼龄茶园春季杂草物种虽然不多,但杂草优势种突出,容易严重发生危害;成年茶园秋季杂草物种丰富度和多样性指数均高于幼龄茶园,成年茶园秋季杂草均匀度指数略低于幼龄茶园,幼龄茶园秋季杂草优势度指数明显低于成年茶园,由此说明,成年茶园秋季杂草发生危害的均衡性差。
近年来,农业生态系统中杂草生物多样性的保护以及发挥其在维持生态平衡中的作用逐渐受到重视[26-31],因此在对农业生态系统中的杂草进行有效控制的同时,也应对其生物多样性给予适当的保护。运用科学方法控制杂草,不仅有利于作物的生长,保持农田生态系统中一定水平的杂草生物多样性,也有利于降低某些优势杂草在群落中的优势地位。本研究结果对指导茶园科学除草,控制优势杂草的危害和保护杂草生物多样性具有重要意义。
致谢:南京农业大学杂草研究室强胜教授帮助修改了论文,特此致谢!
参考文献:
[1]徐华勤,肖润林,向佐湘,等. 不同生态管理措施对丘陵茶园杂草生物多样性的影响[J]. 中国农学通报,2010,26(4):283-286.
[2]李儒海,强 胜,邱多生,等. 长期不同施肥方式对稻油轮作制水稻田杂草群落的影响[J]. 生态学报,2008,28(7):3236-3243.
[3]向佐湘,单武雄,何秋虹,等. 两种生态控草措施对丘陵茶园杂草群落及物种多样性的影响[J]. 中国生态农业学报,2009,17(5):857-861.
[4]李粉华,孙国俊,季 敏,等. 江苏金坛茶园春季主要杂草发生危害调查研究[J]. 江西农业学报,2013,25(4):30-33.
[5]李粉华,孙国俊,季 敏,等. 江苏金坛茶园秋季主要杂草发生危害调查研究[J]. 西南农业学报,2013,26(4):1518-1523.
[6]Tilman D. The ecological consequences of changes in biodiversity:A research for general principles[J]. Ecology,1999,80:1455-1474.
[7]Chesson P. Mechanisms of maintenance of species diversity[J]. Annual Review of Ecology and Systematics,2000,31:343-366.
[8]金坛年鉴编纂委员会. 金坛年鉴(1997—1999)[M]. 北京:中央文献出版社,2000:42-43.
[9]Thomas A C. Weed survey system used in Saskatchewan for cereal and oilseed crops[J]. Weed Science,1985,33(1):34-43.
[10]张朝贤,胡祥恩,钱益新,等. 江汉平原麦田杂草调查[J]. 植物保护,1998,24(3):14-16.
[11]强 胜. 杂草学[M]. 北京:中国农业出版社,2001:1-261.
[12]王英姿,纪明山,祁之秋,等. 辽宁省果园杂草发生情况调查[J]. 植物保护,2008,34(4):98-101.
[13]马克平,刘玉明. 生物群落多样性的测度方法 Ⅰα多样性的测度方法(下)[J]. 生物多样性,1994,2(4):231-239.
[14]方精云,王襄平,沈泽昊,等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性,2009,17(6):533-548.
[15]Wasilewska L. Soil invertebrates as bioindicators,with special reference to soil-inhabiting nematodes[J]. Russian Journal of Nematology,1997,5:117-129.
[16]杨立学,孙跃志. 不同林龄胡桃楸林下植物多样性的差异[J]. 生态学杂志,2013,32(4):807-812.
[17]侯继华,马克平. 植物群落物种共存机制的研究进展[J]. 植物生态学报,2002,26(z1):1-8.
[18]吴 娱,张相锋,董世魁,等. 阿尔金山自然保护区东部典型植物群落的物种组成、多样性及生物量[J]. 生态学杂志,2013,32(9):2250-2256.
[19]李 博. 生态学[M]. 北京:高等教育出版社,2000:1-340.
[20]周德明,王宗永. 南方不同林龄杉木人工林林下物种多样性比较[J]. 林业资源管理,2010(6):65-70.
[21]Harms K E,Condit R,Hubbell S P,et al. Habitat associations of trees and shrubs in a 50 hm2neotropical forest plot[J]. Journal of Ecology,2001,89(6):947-959.
[22]左小安,赵哈林,赵学勇,等. 科尔沁沙地不同恢复年限退化植被的物种多样性[J]. 草业学报,2009,18(4):9-16.
[23]张海艳,孙国俊,季 敏,等. 苏南丘陵茶园秋季杂草种间生态关系及群落分类[J]. 生态学杂志,2013,32(9):2289-2297.
[24]陶福禄,李树人,冯宗炜,等. 豫西山区日本落叶松林下植物物种多样性的研究[J]. 生态学杂志,1998,17(4):2-7,83.
[25]王应刚,李建梅,李淑兰,等. 人为干扰对城市地区植物多样性的影响[J]. 生态学杂志,2004,23(2):102-104.
[26]Tilman D,Knops J,Wedin D,et al. The influence of functional diversity and composition on ecosystem processes[J]. Science,1997,277(5330):1300-1302.
[27]Tilman D,Reich P B,Knops J M. Biodiversity and ecosystem stability in a decade-long grassland experiment[J]. Nature,2006,441(793):629-632.
[28]陈 欣,王兆骞,唐建军. 农业生态系统杂草多样性保持的生态学功能[J]. 生态学杂志,2000,19(4):50-52.
[29]Chen X,Tang J J,Fang Z G,et al. Effects of weed communities with various species numbers on soil features in a subtropical orchard ecosystem[J]. Agriculture,Ecosystems & Environment,2004,102(3):377-388.
[30]Chen X,Yang Y S,Tang J J. Species-diversified plant cover enhances orchard ecosystem resistance to climatic stress and soil erosion in subtropical hillside[J]. Journal of Zhejiang University:Science,2004,5(10):1191-1198.
[31]Yang Y S,Wang H,Tang J J,et al. Effects of weed management practices on orchard soil biological and fertility properties in southeastern China[J]. Soil and Tillage Research,2007,93(1):179-185.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:10:26
热带作物学报(2022年9期)2022-10-17 05:49:06
果树资源学报(2020年3期)2020-05-13 12:53:20
新疆农业科学(2019年11期)2019-12-31 07:02:30
收藏界(2018年1期)2018-10-10 05:23:08
创新作文(小学版)(2018年31期)2018-05-16 03:00:30
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
天然产物研究与开发(2016年11期)2016-06-15 20:29:17
小学阅读指南·低年级版(2016年6期)2016-05-14 21:39:20
中国果菜(2016年9期)2016-03-01 01:28:45
