
慢病毒介导的RASSF1A表达抑制小细胞肺癌H446细胞的生长*
2014-08-08 09:49孔丽君张立霞栾希英张丽沙王绪菡于恒运张月芝
中国病理生理杂志 2014年7期
孔丽君, 张立霞, 栾希英, 张丽沙, 王绪菡, 于恒运, 张月芝
(滨州医学院 1山东省肿瘤分子生物学重点实验室,生物化学与分子生物学教研室, 2烟台附属医院,山东 烟台 264003)
肺癌是我国常见的恶性肿瘤谱中的主要肿瘤之一,目前已发展成为我国癌症死亡率的首位[1],肺癌也是世界上发病率和致死率最高的肿瘤,与吸烟、大气污染、慢性肺病及内在因素都有密切关系,根据其病理特点,肺癌被分为非小细胞肺癌(non-small-cell lung cancer, NSCLC)和小细胞肺癌(small-cell lung cancer, SCLC)。随着环境污染的加重,SCLC的发病率也在逐年增加,约占肺癌的20%,其恶性程度高,倍增时间短,转移早,易复发,尽管对化疗、放疗敏感,但治疗水平在近30多年来仍处于平台期,5年生存率小于2%,因此探讨新的治疗方案具有重要意义。
Ras相关结构域家族1 (Ras-association domain family 1,RASSF1) 基因根据转录后剪切位点的不同有2个主要的转录本RASSF1A和RASSF1C,在正常的上皮细胞中两者均有表达。RASSF1基因位于3p21.3,其编码的蛋白质含有2个结构域,一是羧基末端Ras相关结构域,另一个SARAH结构域。Dittfeld等[2]研究显示RASSF1A的SARAH结构域缺失显著抑制肺癌细胞A549的生长。RASSF1A表达水平抑制主要由其基因启动子区域的高度甲基化引起,相关文献已在乳腺癌、肺癌、原发性肾癌、原发性鼻癌等中有过报道。本文主要探讨RASSFIA抑制小细胞肺癌细胞的生长及其机制。
材 料 和 方 法
1 细胞培养和试剂
本研究所用小细胞肺癌细胞株H446购自中科院上海细胞库,培养基为RPMI-1640加10%胎牛血清以及10%青、链霉素。新霉素(G418)和碘化丙啶(PI)购自Sigma,SYBR Green购自大连宝生物科技有限公司,ECL增强型化学发光液购自Millipore。细胞周期素D1(cyclin D1)、cyclin E、细胞周期素依赖性激酶2 (cyclin-dependent kinase 2, Cdk2)、Cdk4、Cdk6、p15、p16、p18、p19和GAPDH抗体购自Santa Cruz,p21、p27和E2F1抗体购自Cell Signal Technology。
2 方法
2.1慢病毒载体的构建以及包装 含有人RASSF1A的慢病毒表达载体pLV.EX3d.P/neo-EF1A>RASSF1A>IRES/eGFP由赛业广州生物科技有限公司构建,并包装成慢病毒颗粒,病毒滴度为1×1012TU/L。慢病毒载体经测序证实序列的正确性。不含RASSF1A基因的阴性对照病毒由赛业广州生物科技有限公司提供。
2.2稳定表达RASSF1A的小细胞肺癌H446细胞的构建 应用24孔板常规培养H446细胞,每孔接种5×104个细胞,细胞密度大约50%左右。24 h后,细胞贴壁良好,密度70%左右。感染前将慢病毒在冰上缓慢化开,应用完全培养基将病毒稀释10倍,病毒滴度大约在1×1011TU/L左右。应用100 μL稀释后的病毒颗粒取代原先的培养基,12 h后更换完全培养基,96 h后应用荧光倒置显微镜观察感染效率。1 g/L G418进行筛选,15 d后改用维持浓度0.5 g/L维持培养,30 d后挑取细胞生长克隆,继续培养,获取稳定表达RASSF1A的H446细胞命名为RASSF1A-H446。不含RASSF1A的阴性对照细胞命名为GFP-H446。
2.3MTT测细胞生长曲线 将1 000个细胞常规铺96孔板,细胞生长24 h后,每孔加入20 μL MTT(5 g/L),继续培养4 h,PBS洗3遍。每孔加150 μL二甲基亚砜,低速振荡10 min,490 nm波长处测吸光度(A)。
2.4流式细胞术测生长周期 应用6孔板常规培养H446、GFP-H446和RASSF1A-H446细胞,当细胞密度为50%左右时,撤血清使细胞饥饿24 h。然后加入完全培养基培养24 h后,应用0.01%EDTA-0.25%胰酶混合液消化细胞,离心,无菌的PBS洗涤1次。应用70%乙醇4 ℃旋转固定过夜,PI染色30 min,PBS洗涤3 次,流式细胞术检测细胞周期。
2.5Real-time PCR检测细胞周期相关蛋白的mRNA表达 6孔板常规培养H446、GFP-H446和RASSF1A-H446细胞,当细胞密度达到70%时,PBS洗3次。每孔加入1 mL Trizol,吹打15 s左右,让细胞完全破碎,收集裂解液于1.5 mL EP管中。按照说明书操作进行,提取并检测RNA浓度。取2 μg RNA进行反转录。反转录体系如下: RNA 2 μL,5×buffer 4 μL, dNTP 2 μL, 转录酶1 μL,随机引物2 μL,双蒸水5 μL。程序如下:65 ℃预变性5 min;37 ℃ 15 min; 98 ℃ 5 min。将以上反转录得到的cDNA进行real-time PCR检测细胞周期相关基因的表达,引物序列见表1。计算power值,表示不同基因的表达水平。

表1 引物序列
2.6Western blotting检测细胞周期相关蛋白的表达 6孔板常规培养H446、GFP-H446和RASSF1A-H446细胞,当细胞密度达到70%时,PBS洗3次,每孔加入70 μL RAPI裂解液,收集裂解液于1.5 mL EP管中,13 000 r/min、4 ℃离心15 min,分装上清。BCA法测蛋白浓度。每孔加入40 μg蛋白,进行SDS-PAGE,然后转膜、封闭、I抗4 ℃孵育过夜、II抗常温孵育40 min、ECL化学发光液显影、压片、定影。
3 统计学处理
数据用均数±标准差(mean±SD)表示,多组间比较采用单因素方差分析,以P<0.05为差异有统计学意义。
结 果
1 稳定表达RASSF1A的H446细胞的构建
将含有RASSF1A基因的慢病毒颗粒感染H446细胞,96 h后应用荧光显微镜观察细胞的感染效率,发现慢病毒颗粒转染细胞的效率高达80%,见图1A、B。应用1 g/L G418对细胞进行筛选,15 d后细胞出现明显的单克隆细胞群,改用0.5 g/L G418进行维持培养,获得稳定表达RASSF1A的H446细胞,见图1C、D。Western blotting结果表明,LV-RASSF1A组蛋白的表达显著高于空白对照(normal)和阴性对照(LV-GFP)组,见图1E。
2 RASSF1A抑制H446细胞的生长并阻滞细胞周期的运行
RASSF1A具有抑癌蛋白的特性,在80%~100% SCLC组织和细胞中表达缺失。MTT结果表明,LV-RASSF1A组细胞生长缓慢(图2A),而且细胞周期明显阻滞在G1期。图2B显示,LV-RASSF1A组与空白对照组及LV-GFP组间均有明显差异(均P<0.05)。
3 细胞周期相关蛋白的表达
与正常对照和阴性对照相比,在LV-RASSF1A组细胞中,cyclin D1、cyclin E、Cdk2、Cdk4和Cdk6的表达没有明显变化;p21和p27表达明显升高,而p15、p16、p18和p19的表达没有明显变化。由于小细胞肺癌中抑癌基因Rb缺失,在LV-RASSF1A细胞中没有检测到Rb蛋白的表达,但是转录因子E2F1的表达明显降低,见图3。
讨 论
在人类大多数肿瘤细胞中,RASSF1A 的表达缺失是一个常见的事件[3]。在本研究中,我们成功构建了含有RASSF1A基因的慢病毒表达载体,并包装成病毒颗粒。将该病毒颗粒感染小细胞肺癌H446细胞,G418筛选后,获得了稳定表达RASSF1A基因的H446细胞克隆(命名为:RASSF1A-H446)。MTT和流式细胞术检测发现细胞生长明显受到抑制,而且G1期阻滞较明显。以上结果表明RASSF1A能够明显抑制细胞增殖,而且使细胞周期阻滞在G1期。为探讨RASSF1A抑制细胞生长和G1期阻滞的机制,我们从mRNA和蛋白的水平分析了G1/S期细胞周期相关蛋白的表达,real-time PCR 和Western blotting结果显示cyclin D1、cyclin E、Cdk2、Cdk4、Cdk6、p15、p16、p18和p19的表达没有明显变化,而p21和p27的表达明显升高,E2F1的表达显著降低。以上结果表明RASSF1A主要通过升高p21、p27和降低E1F2的表达,抑制细胞的生长。
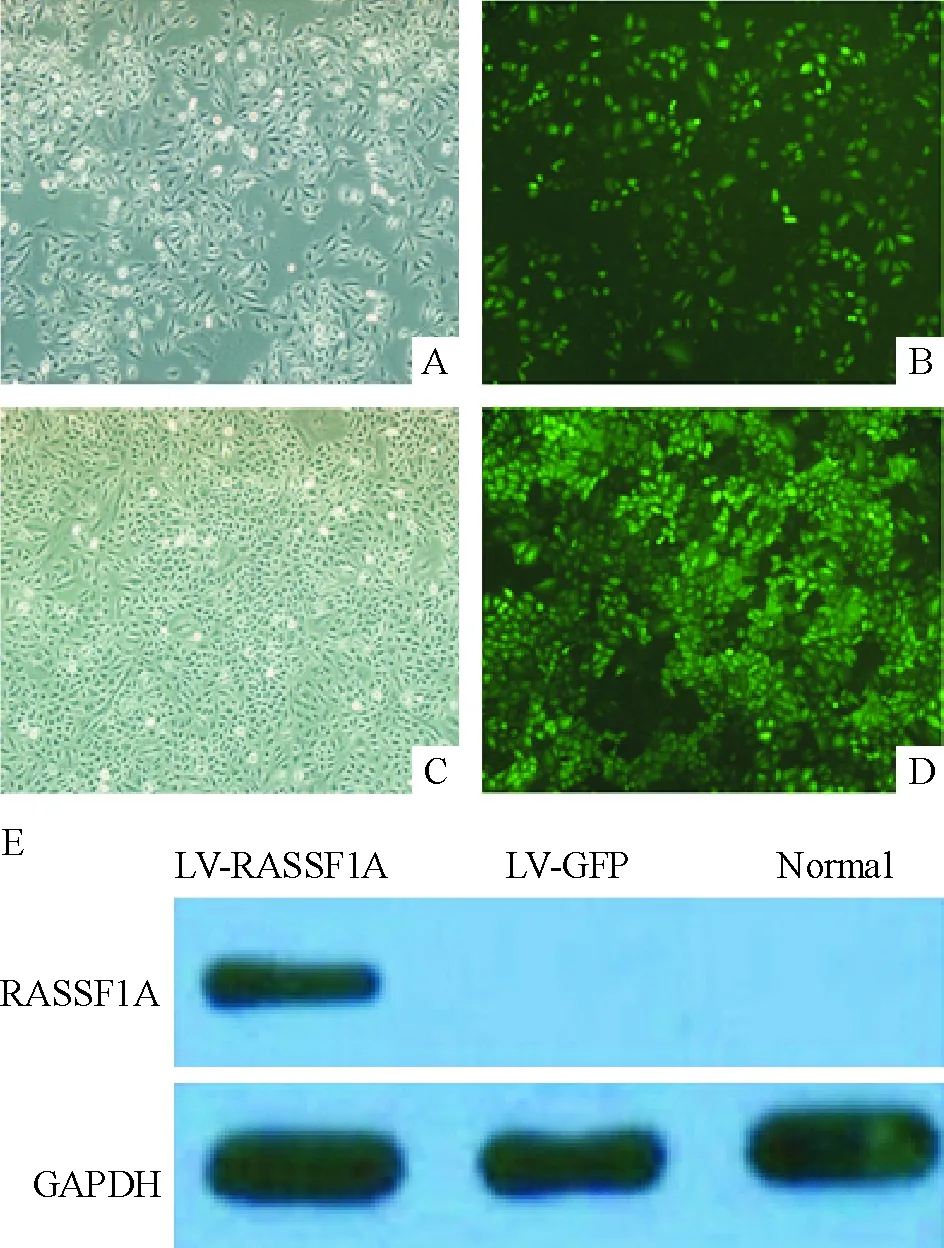
Figure 1. H446 cells transfected with RASSF1A gene were constructed and RASSF1A expression was detected (×100). A, B: the pictures of H446 cells after transfection for 96 h; C,D: the clones that stably expressed RASSF1A were obtained; E: RASSF1A was detected when H446 cells were infected by lentivirus containing RASSF1A gene.
95%以上的小细胞肺癌,由于Rb基因表达缺失,放松了对E2F1的管制。持续活化的E2F1促进细胞生长、侵袭和转移相关基因的持续表达,最终导致肿瘤的发生[4]。p21和p27作为细胞周期素依赖性激酶抑制剂,主要是抑制了Cdk2/cyclin E的活性,从而抑制转录因子FoxO1活性,抑制相关基因的转录[5]。RASSF1A通过促进p21和p27的表达,可能抑制了FoxO1的活性;同时还直接抑制了转录因子E2F1的表达,导致促进细胞生长、侵袭和转移相关基因转录水平的降低,抑制了细胞的生长,使细胞停滞在G1期。

Figure 2. The growth curve and cell cycle of RASSF1A cells. A: the growth curve was detected by MTT assay; B: the cell cycle was analyzed by flow cytometry.Mean±SD. n=3. *P<0.05 vs normal or LV-GFP.
RASSF1A作为一个抑癌蛋白,其抑制细胞生长的机制被广泛探讨。RASSF1A蛋白羧基末端的Ras蛋白结合的结构域能够与Ras蛋白相互结合,进而抑制细胞的生长[2]。还有文献报道,RASSF1A能够抑制cyclin D1在细胞内的聚集,进而影响细胞周期的顺利进行[6]。本研究中,我们发现RASSF1A在小细胞肺癌中也发挥抑癌作用,其抑制细胞生长的主要作用机制是促进p21和p27的表达,同时降低E2F1的表达,进而使细胞周期阻滞在G1期,抑制细胞的增殖,以上结果为小细胞肺癌基因治疗提供新的靶点。
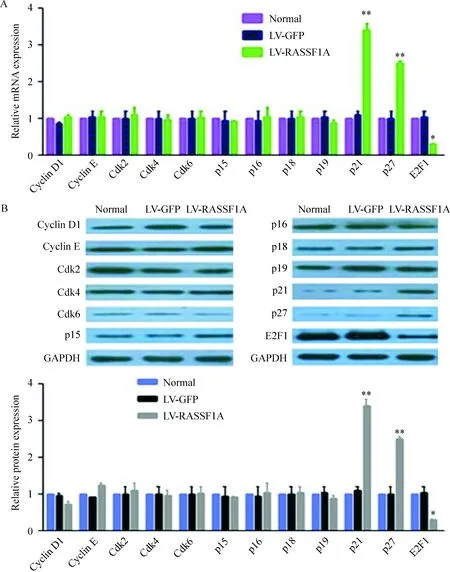
Figure 3. The mRNA (A) and protein (B) levels of cell cycle-associated proteins detected by real-time PCR and Western blotting. Mean±SD. n=3.*P<0.05,**P<0.01 vs normal or LV-GFP.
[参 考 文 献]
[1] 邹 敏,周韶璋,于起涛,等. 非小细胞肺癌患者肿瘤组织和血清表皮生长因子受体第19、21外显子的检测及其临床特征的分析[J].中国病理生理杂志,2013,29(5):839-844.
[2] Dittfeld C, Richter AM, Steinmann K, et al. The SARAH domain of RASSF1A and its tumor suppressor function[J]. Mol Biol Int, 2012,2012: 196715.
[3] Dammann R, Yang G, Pfeifer GP. Hypermethylation of the CpG island of Ras association domain family 1A (RASSF1A), a putative tumor suppressor gene from the 3p21.3 locus, occurs in a large percentage of human breast cancers[J]. Cancer Res, 2001, 61(7):3105-3109.
[4] Li J, Hu SX, Perng GS, et al. Expression of the retinoblastoma (RB) tumor suppressor gene inhibits tumor cell invasioninvitro[J]. Oncogene, 1996, 13(11): 2379-2386.
[5] Lin SY, Black AR, Kostic D, et al. Cell cycle-regulated association of E2F1 and Sp1 is related to their functional interaction[J]. Mol Cell Biol, 1996, 16(4):1668-1675.
[6] Shivakumar L, Minna J, Sakamaki T, et al. The RASSF1A tumor suppressor blocks cell cycle progression and inhibits cyclin D1 accumulation[J]. Mol Cell Biol,2002,22(12):4309-4318.
猜你喜欢
流体机械(2022年11期)2023-01-09
核科学与工程(2022年3期)2022-10-18
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
中成药(2017年9期)2017-12-19
中成药(2017年7期)2017-11-22
中华老年多器官疾病杂志(2016年7期)2016-04-28
船海工程(2015年3期)2015-10-21
中国老年学杂志(2015年9期)2015-01-31
中成药(2014年10期)2014-02-28
