
PI3K/Akt信号通路在脓毒症肾脏损伤诱导的自噬中的调节作用*
2014-08-08 09:20:48向镜芬龚剑锋雷伟健邓艳琼钟国权孟启勇
中国病理生理杂志 2014年6期
向镜芬, 杨 祥, 龚剑锋, 雷伟健, 邓艳琼, 牟 丹, 钟国权, 孟启勇
(暨南大学第五附属医院,清远市人民医院重症医学科, 广东 清远 511500)
脓毒症是由感染引发的全身性炎症反应综合征,是ICU患者发生急性肾损伤(acute kidney injury, AKI)的常见原因[1]。AKI是预测脓毒症患者死亡的独立危险因素[2]。寻找介导脓毒症AKI肾小管上皮细胞损伤的关键分子并阐明其调控机制,对于早期干预脓毒症AKI具有重要的科学意义。近年来研究发现细菌内毒素及其炎症因子是诱导肾脏损伤的直接且重要的原因。脂多糖(lipopolysaccharide,LPS)刺激机体引发严重炎症反应,常常产生大量致炎症因子如肿瘤坏死因子α(tumor necrosis factor α,TNF-α)和活性氧簇(reactive oxygen species,ROS),从而导致氧化应激、线粒体损伤和能量耗竭,最终导致肾小管上皮细胞凋亡[3]。在此过程中,细胞自噬对于肾小管上皮损伤和修复可能发挥了重要的作用。
自噬是真核细胞处于缺氧、饥饿或感染时,在自噬相关基因的调控下,利用溶酶体降解自身受损的细胞器和大分子物质,为细胞修复、更新提供原料与营养、保持能量稳态的过程,是有别于凋亡的另一种程序性死亡机制[4],磷脂酰肌醇3-激酶/蛋白激酶B(phosphoinositide 3-kinase/protein kinase B,PI3K/Akt)通路是影响细胞自噬重要的信号通路[5]。近年来有研究表明,自噬在脓毒症的致病过程中发挥重要作用[6-8]。那么自噬是否参与了脓毒症时肾小管上皮细胞的损伤机制?PI3K/beclin-1复合物的形成对脓毒血症时肾小管上皮细胞的自噬是否至关重要?值得深入研究。本研究拟建立盲肠结扎穿孔(cecal ligation and puncture,CLP)模拟腹膜炎大鼠脓毒症模型,观察肾小管上皮细胞是否存在自噬现象。并且进一步进行体外实验证实是否一定浓度的LPS可刺激肾小管上皮细胞发生自噬,探讨PI3K/Akt信号通路是否通过形成复合物促进自噬对脓毒症时肾小管上皮细胞的保护作用,为临床干预脓毒症AKI提供新的策略。
材 料 和 方 法
1 材料
雄性SD大鼠,250~300 g,9只购自中山大学动物中心。HK-2细胞株购自ATCC。BCA蛋白浓度检测试剂盒购自Pierce。微管相关蛋白轻链3(microtubule-associated protein light chain 3,LC3)抗体购自Sigma。p-Akt(308)抗体、p-Akt(472)抗体、Akt抗体、beclin-1抗体、GAPDH抗体和活化caspase-3 抗体均购自CST。多聚赖氨酸购自Sigma。DMEM/F12培养基和胎牛血清购于Gibco。二甲基亚砜购自Sigma。山羊抗兔Ⅱ抗为Jackson产品。其它所用生化试剂均为国产分析纯。
2 方法
2.1脓毒症诱发急性肾损伤模型的构建 SD大鼠麻醉用 10%水合氯醛(生理盐水配制)腹腔注射,剂量为4 mL/kg。在腹部中线打开腹腔(切口3~4 cm),找到盲肠,仔细将盲肠间的肠系膜剪开(注意不要剪断膜上的血管,以避免大出血),在离盲肠4 cm处用5-0丝线结扎,18G针头(10 mL注射器针头)穿刺4次,在穿孔处轻轻挤出1 mm排泄物。6-0丝线两层缝合伤口。腹腔注射(50 mL/kg皮下注射)预热生理盐水,笼中苏醒。CLP 6 h后皮下注射广谱抗生素(亚胺培南/西司他丁)14 mg/kg(10 mL预热生理盐水)。麻醉动物后6 h和24 h,采血、处死并摘取肾脏。
2.2肾损伤指标检测
2.2.1组织切片糖原染色 各组处死后取右侧肾脏一部分,将剩余部分的肾皮质置于液氮中冻存后转至-70 ℃ 存放, 用于Western blotting检测蛋白的表达。将肾脏置于甲醛溶液固定48 h,放入自动脱水机中进行固定,共设定 12 h,固定后再进行石蜡包埋,包埋后在切片机上进行切片,后将切片放入防脱载玻片上,置于4 ℃冰箱保存。PAS染色步骤如下:常规切片 2~3 μm,脱蜡至水,0.5%高碘酸氧化 6~8 min,蒸馏水洗,无色品红加盖避光7 min,不经水洗,擦净组织旁多余染液,0.5%偏重亚硫酸钠作用 1 min,水洗 2~5 min,Harris 苏木素复染 1 min 左右,水洗,1%盐酸乙醇分化。片刻,自来水反复冲洗,温水反蓝,梯度乙醇脱水,二甲苯透明,中性树胶封片。置于普通光学显微镜下观察肾间质及肾小管的变化并拍照。
2.2.2血尿素氮(blood urea nitrogen,BUN)及血清肌酐(serum creatinine, SCr)检测 实验结束小鼠腹腔麻醉后心脏取血, 一部分离心分离血清, 酶法测定BUN和Scr浓度[9]。所需试剂盒均购自南京建成科技有限公司。
2.3Western blotting检测信号蛋白的表达 CLP大鼠肾脏组织置于匀浆器中,冰上匀浆后,加入蛋白裂解液及PMSF,冰上放置40 min,4 ℃、12 000 r/min 离心 40 min,取上清。以BSA为标准,用Bradford法对上清进行蛋白定量,取20~40 μg蛋白样品,10%SDS-PAGE,100 V转移1 h至PVDF膜,放入封闭液中37 ℃ 1 h,Ⅰ抗4 ℃过夜;次日TBST洗膜5 min 3次,将膜与碱性磷酸酶标记的抗IgG抗体孵育,室温摇床上轻摇1 h,TBST洗膜5 min 3次,曝光,用图像分析测定各吸光度值作定量分析。
2.4HK-2细胞培养 HK-2细胞于10%胎牛血清+DMEM细胞培养液37 ℃、5%CO2条件下培养,0.25%胰蛋白酶消化、传代。将HK-2细胞以2×108/L接种于6孔板,待细胞生长至70%~80%融合时,用无血清无添加因子的Keratinocyte-SFM培养液培养24 h,使细胞生长同步化。然后将细胞置于无血清无添加因子的培养基中,分别用加入不同浓度的LPS(0、0.1、10和20 mg/L)培养12 h;而在另外一组加入10 mg/L LPS,然后培养不同时间(0~12 h),收集细胞总蛋白,进行Western blotting检测。
2.5Hoechst 33342和Annexin V/PI双染定量和定性凋亡 将HK-2细胞以2×108/L接种于6孔板并分为4组:LPS刺激组、LPS+3-甲基腺嘌呤(3-me-thyladenine,3-MA)组、LPS+Akt inhibitor组和对照组(加PBS),抑制剂均购自Sigma。分组处理后,去除培养液,消化,离心,收集细胞,PBS洗2遍,加入5 μL Hoechst 33342染色液和5 μL PI染色液。混匀后,4 ℃孵育20~30 min,PBS洗1次,在荧光显微镜下观察细胞凋亡情况。同上培养,收集细胞后,用4 ℃预冷的PBS洗涤2次,1 000 r/min离心5 min;用250 μL结合缓冲液重悬细胞,取100 μL细胞悬液于5 mL流式管中加入5 μL Annexin V和10 μL PI溶液,室温避光染色10 min,加入300 μL PBS后用流式细胞仪检测凋亡细胞、正常细胞、坏死细胞,对结果进行统计分析。同时,离心收集细胞进行Western blotting检测LC3的表达。
3 统计学处理
用SPSS 13.0统计软件分析,数据以均数±标准差(mean±SD)表示,进行方差分析和LSD-t检验,以P<0.05为差异有统计学意义。
结 果
1 CLP大鼠肾脏发生明显损伤
与假手术组比较,盲肠结扎穿刺动物普遍出现肾损伤,且随着时间的推移,损伤程度加剧,出现肾小管上皮细胞膨胀,刷状缘缺失,空泡变性,坏死,管型形成,细胞脱落等病理改变,见图1A。与假手术组比较,CLP组动物在术后,随着时间的推移,BUN和SCr均有上升,见图1B、C。

Figure 1. The changes of kidney histopathology, blood urea nitrogen (BUN) and serum creatinine (SCr) in CLP and control (sham) rats. A: HE staining (×40); B: BUN; C: SCr. Mean±SD. n=3.*P<0.05 vs sham group.
2 CLP大鼠肾脏自噬相关蛋白LC3、beclin-1含量及Akt磷酸化水平的变化
Western blotting结果表明,对照组(0 h)LC3表达较低,CLP发生6 h后开始上调,12 h达到最高峰,18 h有所下降;Beclin-1表达趋势与LC3相同,见图2A,证明CLP肾脏损伤时有显著的自噬发生。Wes-tern blotting显示细胞活化caspase-3蛋白表达水平升高,并呈时间依赖性,说明CLP肾脏损伤诱导了caspase-3介导的细胞凋亡,呈时间依赖性,见图2A。Western blotting结果表明,对照组p-Akt表达较低,CLP发生2 h后开始上调,6 h达到最高峰,12 h有所下降;Akt在CLP肾脏损伤后随时间的推移无明显变化,见图2B。

Figure 2. The protein expression of LC3, beclin-1,activated caspase-3 and p-Akt in renal tissues at different time points after CLP detected by Western blotting. A: expression of LC3, beclin-1 and activeated caspase-3; B: expression of p-Akt(308) and p-Akt(472). Mean±SD. n=3.*P<0.05 vs 0 h.
3 体外细胞实验检测自噬相关蛋白LC3含量及Akt磷酸化水平
3.1不同浓度LPS刺激HK-2细胞后自噬相关蛋白的表达 Western blotting结果表明,体外不同浓度LPS刺激HK-2细胞后,对照组(未刺激组)p-Akt(308)表达水平较低,随着刺激浓度的增加,p-Akt(308)表达水平逐渐提高;对照组(未刺激组)p-Akt(472)表达水平较低,0.1 mg/L LPS刺激后p-Akt(472)表达水平逐渐提高,10 mg/L LPS刺激组p-Akt(472)表达水平最高,20 mg/L LPS刺激组p-Akt(472)表达水平有所下降。LC3表达水平与p-Akt(472)表达变化趋势一致,见图3A。
3.2LPS刺激HK-2细胞后不同时点自噬相关蛋白的表达 Western blotting结果表明,对照组(未刺激组)p-Akt(308)表达水平较低,随着刺激时间的延长,p-Akt(308)表达水平逐渐提高;对照组(未刺激组)p-Akt(472)表达水平较低,LPS 刺激4 h后p-Akt(472)表达水平逐渐提高,8 h p-Akt(472)表达水平最高,12 h p-Akt(472)表达水平有所下降。LC3表达水平与p-Akt(472)表达变化趋势一致,见图3B。
4 PI3K 信号通路抑制剂对LPS刺激HK-2细胞后自噬相关蛋白表达的影响
Western blotting分析表明,与LPS刺激组相比,抑制剂组(LPS+3-MA和LPS+Akt inhibitor组)LC3表达量显著下调,见图4A。Hoechst 33342和Annexin V/PI双染进行细胞凋亡实验,结果表明与LPS刺激组相比,抑制剂组(LPS+3-MA和LPS+Akt inhibitor组)HK-2细胞凋亡明显增加(P<0.05),见图4B。

Figure 3. The protein expression of LC3 and p-Akt in HK-2 cells after LPS treatment at different time points and different concentrations. A: LPS treatment at 0, 0.1, 10 and 20 mg/L for 12 h; B: 10 mg/L LPS treatment at 0, 4, 8 and 12 h. Mean±SD. n=3. *P<0.05 vs 0 mg/L or 0 h.
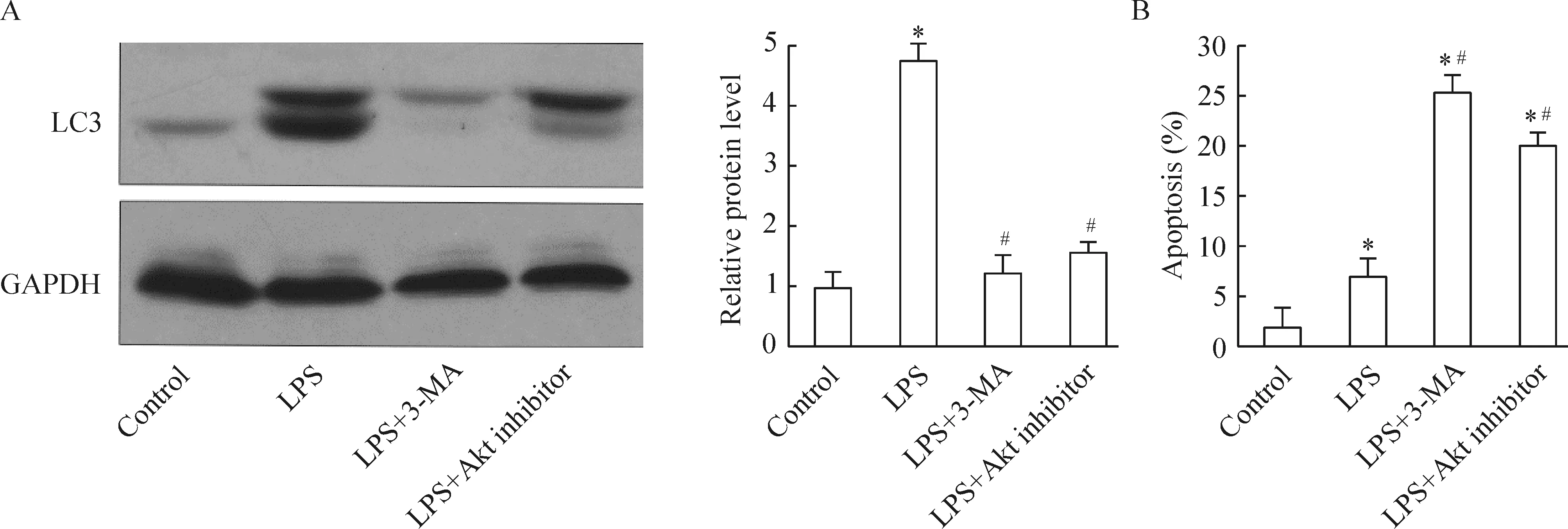
Figure 4. The levels of autophagy-related protein LC3 in HK-2 cells (A) and apoptosis of HK-2 cells (B) treated with LPS plus PI3K inhibitor or Akt inhibitor.Mean±SD.n=3.*P<0.05 vs control; #P<0.01 vs LPS alone.
讨 论
脓毒症合并AKI患者死亡率明显高于不伴AKI者,病死率超过70%[1],成为危重病急救医学面临的急需解决的难题之一。研究发现,脓毒症所致AKI的发生机制不同于经典的缺血再灌注引起的AKI[10]。过去认为脓毒症合并AKI时的主要病理生理机制主要是肾小管管周微循环障碍导致肾脏低灌注,及肾脏缺血后引起的急性肾小管坏死(acute tubular necrosis,ATN)。然而,脓毒症AKI死亡患者肾脏病理显示,70%患者并无ATN发生,而肾小管上皮细胞凋亡更为常见[11]。另一方面,脓毒症动物模型肾脏血流动力学结果显示,近2/3提示肾脏缺血,而1/3提示肾脏血流量保持不变或增加,后者更接近人体发生脓毒症的高动力型脓毒休克状态[11]。在肾脏血流量不变甚至增加的情况下,肾小管上皮细胞仍然发生凋亡,提示肾脏血流动力学改变仅仅是脓毒症引起肾脏损伤的一部分原因。而细菌内毒素及其炎症因子是诱导肾脏损伤的直接且重要的原因。以LPS为代表的内毒素,刺激机体引发严重炎症反应,常常产生大量致炎症因子如TNF-α和ROS,从而导致氧化应激、线粒体损伤和能量耗竭,最终导致肾小管上皮细胞凋亡。在此过程中,细胞自噬对于肾小管上皮损伤和修复可能发挥了重要的作用。
自噬是真核细胞处于缺氧、饥饿或感染时,在自噬相关基因的调控下,利用溶酶体降解自身受损的细胞器和大分子物质,为细胞修复、更新提供原料与营养、保持能量稳态的过程,是有别于凋亡的另一种程序性死亡机制。凋亡的最终结局是细胞死亡,而自噬是细胞存活与死亡的一把“双刃剑”[12]。同时两者在一定程度上又密不可分:自噬为凋亡所需,自噬通常先于凋亡,进而启动凋亡;适度的自噬抑制凋亡,保护细胞免于发生凋亡和坏死;自噬过度可以向凋亡转化,共同促进细胞死亡[13]。目前关于自噬在肾小管上皮细胞中的研究多集中在缺血再灌注、药物肾损伤和梗阻性肾病方面[14-16]。近年来有研究表明,自噬在脓毒症的致病过程中发挥重要作用。脓毒症患者肝脏组织及脓毒症大鼠模型的心脏、肝脏中可观察到自噬小体产生,同时伴有线体粒氧化损伤,证明脓毒症时组织细胞有自噬现象[6-8]。此外,在CLP模拟腹膜炎及LPS腹腔注射这2种脓毒症小鼠模型中都看到,敲除关键性自噬基因LC3及beclin-1后,炎症因子IL-1β、IL-18等分泌明显增加[17],减少自噬将促进或加重炎症反应并导致细胞死亡[18],提示自噬在脓毒症发病及炎症反应中起到重要作用。PI3K/Akt是一条经典的存活信号通路,在细胞凋亡和肿瘤发生中发挥着重要作用。PI3K可以使其下游分子Akt磷酸化。近些年来,在许多研究中证明了PI3K/Akt信号通路可以通过下游分子mTOR来调节自噬。
我们建立了CLP模拟腹膜炎大鼠脓毒症模型,观察肾小管上皮细胞是否自噬现象。观察到随着时间的推移,肾脏损伤程度加剧,出现肾小管上皮细胞膨胀、刷状缘缺失、空泡变性、坏死、管型形成、细胞脱落等病理改变,同时BUN和SCr均有上升,提示CLP模拟腹膜炎大鼠脓毒血症造成肾脏损伤的模型构建成功。为了检测CLP肾脏损伤时肾小管上皮细胞是否发生自噬,我们采用Western blotting检测了自噬相关分子的表达,结果表明肾脏损伤自噬相关蛋白LC3、beclin-1含量及Akt磷酸化水平均随着CLP肾脏损伤发生时间的推移表达上升。我们发现CLP后beclin-1 表达于6 h开始上调,12 h达最高峰,有趣的是beclin-1 表达量在18 h有所下调,可能由于长时间内毒素刺激诱导肾小管上皮细胞大量凋亡,caspase-3过度活化,而活化的 caspase-3具有剪切 beclin-1 的作用[18],从而导致beclin-1 表达下降。我们进一步在体外实验证实了一定浓度的LPS可刺激肾小管上皮细胞发生自噬。不同浓度的LPS以及不同刺激时间均可促使HK-2细胞的自噬相关蛋白LC3及p-Akt表达量上升。同时,使用PI3K及Akt抑制剂后自噬相关蛋白的表达量有所下降,同时HK-2细胞凋亡明显增加,提示PI3K/Akt信号通路可能对CLP肾脏损伤诱导的自噬有调节作用。
本研究通过建立经典的脓毒症AKI大鼠模型(CLP模型)和LPS细胞模型,明确脓毒症时肾小管上皮细胞是否发生自噬;采用公认的自噬抑制剂3-MA和PI3K抑制剂观察它们对脓毒症肾小管上皮细胞损伤的影响,探讨beclin-1及Akt是否通过通过促进自噬对脓毒症肾小管上皮细胞的保护作用。本研究有助于揭示脓毒症时肾小管上皮细胞损伤的新机制,为临床干预脓毒症AKI提供新的策略。
[参 考 文 献]
[1] Schrier RW, Wang W. Acute renal failure and sepsis[J]. N Engl J Med, 2004,351(2):159-169.
[2] Bagshaw SM, Uchino S, Bellomo R, et al. Septic acute kidney injury in critically ill patients: clinical characteristics and outcomes[J]. Clin J Am Soc Nephrol, 2007,2(3):431-439.
[3] 克拉拉·阿巴斯, 吴立玲. 脓毒血症时大鼠肾脏Gq/11蛋白的变化[J]. 中国病理生理杂志, 2000,16(10):1097.
[4] Klionsky DJ, Emr SD. Autophagy as a regulated pathway of cellular degradation[J]. Science, 2000,290(5497):1717-1721.
[5] Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism[J]. Cell, 2006,124(3):471-484.
[6] Chien WS, Chen YH, Chiang PC, et al. Suppression of autophagy in rat liver at late stage of polymicrobial sepsis[J]. Shock, 2011,35(5):506-511.
[7] Watanabe E, Muenzer JT, Hawkins WG, et al. Sepsis induces extensive autophagic vacuolization in hepatocytes: a clinical and laboratory-based study[J]. Lab Invest, 2009,89(5):549-561.
[8] Hsieh CH, Pai PY, Hsueh HW, et al. Complete induction of autophagy is essential for cardioprotection in sepsis[J]. Ann Surg, 2011,253(6):1190-1200.
[9] 卢晓梅, 金玉楠, 于艳秋,等. 奥美沙坦酯对慢性心力衰竭小鼠肾脏损伤的保护作用[J]. 中国病理生理杂志, 2011,27(1):145-148.
[10] Wan L, Bagshaw SM, Langenberg C, et al. Pathophysio-logy of septic acute kidney injury: what do we really know?[J]. Crit Care Med, 2008,36(4 Suppl):S198-S203.
[11] Langenberg C, Wan L, Egi M, et al. Renal blood flow in experimental septic acute renal failure[J]. Kidney Int, 2006,69(11):1996-2002.
[12] Vellai T, Toth ML, Kovacs AL. Janus-faced autophagy: a dual role of cellular self-eating in neurodegeneration?[J]. Autophagy, 2007,3(5):461-463.
[13] Hsieh YC, Athar M, Chaudry IH. When apoptosis meets autophagy: deciding cell fate after trauma and sepsis[J]. Trends Mol Med, 2009,15(3):129-138.
[14] Isaka Y, Kimura T, Takabatake Y. The protective role of autophagy against aging and acute ischemic injury in kidney proximal tubular cells[J]. Autophagy, 2011,7(9):1085-1087.
[15] Bolisetty S, Traylor AM, Kim J, et al. Heme oxyge-nase-1 inhibits renal tubular macroautophagy in acute kidney injury[J]. J Am Soc Nephrol, 2010,21(10):1702-1712.
[16] Li L, Zepeda-Orozco D, Black R, et al. Autophagy is a component of epithelial cell fate in obstructive uropathy[J]. Am J Pathol, 2010,176(4):1767-1778.
[17] Nakahira K, Haspel JA, Rathinam VA, et al. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome[J]. Nat Immunol, 2011,12(3):222-230.
[18] Carchman EH, Rao J, Loughran PA, et al. Heme oxyge-nase-1-mediated autophagy protects against hepatocyte cell death and hepatic injury from infection/sepsis in mice[J]. Hepatology, 2011,53(6):2053-2062.
猜你喜欢
中老年保健(2022年3期)2022-08-24 02:58:10
现代临床医学(2021年6期)2021-11-20 06:34:42
中华养生保健(2020年4期)2020-11-16 01:31:40
中国中医急症(2019年10期)2019-05-21 07:20:46
中国生殖健康(2019年10期)2019-01-07 01:21:22
特别健康(2018年9期)2018-09-26 05:45:46
中华老年多器官疾病杂志(2016年9期)2016-04-28 08:52:44
中外医疗(2015年11期)2016-01-04 03:58:45
医学研究杂志(2015年8期)2015-06-22 14:00:57
医学研究杂志(2015年12期)2015-06-10 06:57:46
