
川西亚高山不同海拔岷江冷杉树轮碳稳定同位素对气候的响应
2014-08-04 02:26:02刘世荣姜春前
生态学报 2014年7期
靳 翔,徐 庆,*,刘世荣,姜春前
(1.中国林业科学研究院森林生态环境与保护研究所国家林业局森林生态环境重点实验室,北京100091;2.中国林业科学研究院林业研究所,北京100091)
树木年轮稳定同位素技术因其信息分辨率高、生理解释性强、反映环境参数多和精确度高等优点近年来日益受到生态学和气象学家的重视,已成为古气候、古环境及大气CO2变化等方面研究的重要途径之一[1-6]。树木年轮(简称树轮)稳定同位素的研究在国外已有较多的报道[1,7-9]。在我国,树轮稳定同位素研究较晚[10-13],发展较快,碳稳定同位素技术在树木年轮气候学研究中的优势在于树轮中δ13C变化可以反映树木整个生命过程中光合作用和水分循环之间的平衡状态,并记录了对其所在自然环境的生理响应[4-5],特别是对于中国这样典型的季风气候国家,开展运用碳稳定同位素技术研究树轮对气候要素的响应将具有重要的理论意义[13]。
树木生长主要受两方面因子制约:一是树木本身的遗传因子影响,二是受外界环境条件的支配,因此树轮成为外部气候要素变化的记录器[14]。海拔梯度作为树木生长的一个重要的生态因子,往往造成热量和水分的重新分配,使得树木生长的环境更为复杂。早期关于树木年轮的海拔梯度研究主要是以调查树木生长为主,利用树轮碳稳定同位素研究气候变化的响应是一个复杂的过程,且不同地区不同树种年轮δ13C对气候因子的响应也不同[14-17]。随着树轮气候学研究的开展,不同海拔高度上影响树木生长的环境要素研究越来越受到重视。
前人对川西亚高山森林植物对气候变化响应的研究,主要侧重采用传统的研究方法(如树轮宽度)分析重建过去气候条件[6,18-20];在区域范围内不同海拔的树木年轮指标对气候要素响应的研究相对较少[13,21-23],运用稳定碳同位素技术研究川西卧龙亚高山不同海拔高度主要代表树种岷江冷杉(Abies faxoniana)对气候要素的响应的研究未见报道。
1 研究区自然概况
卧龙自然保护区系成都平原向青藏高原过渡的高山深谷地带,属青藏高原气候区的东缘,其西风急流南支和东南季风控制了该地区的主要天气,冬半年(11月—翌年4月)在干冷的西风急流南支影响下,天气多晴朗干燥,在冷气流的进退过程中,也常形成降雪或雨;夏半年(5—10月)湿润的东南季风顺河而上,遇到高山冷气流而形成丰富的迎坡降水,因而温暖湿润。该地区年均相对湿度80.3%,冬半年为75.5%,夏半年为84.8%,全年无霜期180—200 d,年均气温(8.5±0.5)℃,7月平均气温(17.1±0.8)℃,1月平均气温-0.9℃,年均日照950 h,年降水量(890±100)mm[24]。
岷江冷杉是川西卧龙巴郎山暗针叶林的优势树种,也是暗针叶林的建群种,主要分布于海拔2600—3200 m处,常见的树种有铁杉(Tsuga spp.)和麦吊云杉(Picea brachytyla)等。
2 研究方法
2.1 样品采集与预处理
2010年7 月在四川卧龙巴郎山(邛崃山)阴坡亚高山暗针叶林(102°52'—103°25'E,30°45'—31°25'N)中采集岷江冷杉树木年轮样本。分别在海拔2750 m,2950 m和3150 m处设置3个研究样地(200 m×30 m,即低海拔区、中海拔区和高海拔区),基于国际树木年轮库(ITRDB)的要求,分别在3个海拔高度(研究样地)处采集岷江冷杉树轮样本各20株,尽量选取人类干扰较少、径级较大且年龄较老(>50a以上)的优势树木个体,每个样木取树芯2个,采集到的样芯装入内径(5.5 mm)稍大于样芯直径的采样管中,并在样管上用油性笔标注代码密封保存。
本研究选取年轮较宽无明显差异、生长正常、缺轮较少、年轮界限较为明显、敏感度较高、没有异常的树轮样芯,利用木板夹对样芯进行固定、打磨,使样本达到光、滑、亮,轮界清晰分明,细加工后至少保持原来直径的1/2以上部分。打磨后的样芯截面平坦不能出现斜面,且能够在显微镜下清晰地分辨细胞大小,对在显微镜下仍看不清楚的,需要进一步用更细的砂纸抛光。然后对比宽度年表交叉定年,确定每根粗芯准确日历年并利用不锈钢针做标记。自外向内依次在显微镜下用手术刀对树轮样芯逐轮剥离,为防止碳源污染,样品在玻璃垫板上利用不锈钢手术刀在双目显微镜下进行样品的逐轮剥离(每年1个样),利用硫酸纸把剥离的样品转移到经过净化处理的样品瓶中,将不同样芯相同年份的样品混合为一个样品,并在样品瓶上进行编号。把装入样品瓶的待测样品放置在75℃下烘干24 h。将烘干的样品利用高速离心球磨仪粉碎至60目(300 μm),以待提取纤维素。
在Flash EA1112 HT经过高温燃烧后生成CO2,经纯化后进入DELTA V Advantage同位素质谱仪分析稳定碳同位素组成。所有样品测定和分析在中国林业科学研究院国家重点开放实验室完成,碳同位素(δ13C)测量精度为±<0.1‰。

式中,Rsample样品中元素的重轻同位素丰度之比,Rstandard是国际通用标准物的重轻同位素丰度之比[25]。
2.2 气候数据来源
气象数据源自离采样点较近的都江堰和小金气象站(国家标准气象台站,经纬度分别为103°34'E,30°59'N和102°21'E,31°N),由国家气象局(http://cdc.cma.gov.cn/)提供。所用气候资料包括月值资料和年值资料,其中有1955—2009年完整年的月平均温度、月降水量和月平均相对湿度,经检验,2站点的气象数据可靠,无明显突变,可被认为代表了当地自然界的气候变化趋势(图1)。采用Kendall方法检查了序列是否存在突变点,用Double-mass方法检查序列的非随机变化。检验的结果表明,该站气候数据可用来代表自然气候的变化。
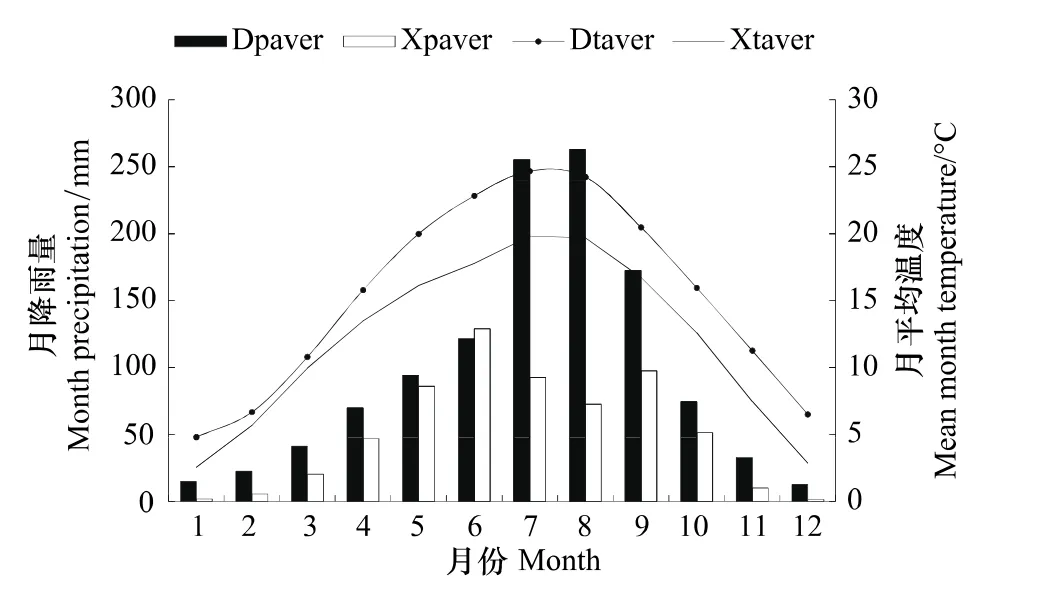
图1 川西地区都江堰和小金气象站月平均温度和月平均降雨量(1955—2009年)Fig.1 Variations of mean month temperature and mean month precipitation of Dujiangyan and Xiaojin meteorological station in western Sichuan from 1955 to 2009
3 结果与分析
3.1 树轮稳定碳同位素序列统计特征
川西卧龙亚高山岷江冷杉树轮碳稳定同位素序列(1904—2009)变化范围为-23.33‰—-26.31‰,平均值为-24.91‰,变异系数均小于零,表明岷江冷杉树轮碳稳定同位素序列在年际变化中较为稳定;并有较强的一阶自相关性(表1)。从表1可以看出,岷江冷杉树轮碳稳定同位素序列在不同海拔高度之间存在着差异,没有表现出明显的海拔趋势,而在同一海拔高度的岷江冷杉树轮碳稳定同位素序列的变化趋势基本一致。3个不同海拔高度的岷江冷杉树轮碳稳定同位素一阶自相关系数分别为0.651、0.936、0.333,中海拔(海拔2950 m)的岷江冷杉树轮碳稳定同位素一阶自相关系数最大,表明其具有更为显著的“滞后效应”,是因为中海拔区气候相对适宜,更有利于研究岷江冷杉碳稳定同位素与气候因子之间的关系。

表1 川西亚高山不同海拔岷江冷杉稳定碳同位素序列的基本统计特征Table1 Statistical results of stable carbon isotope of detrend series of tree-rings of Abies faxoniana in different elevations
3.2 大气CO2浓度校正
树木生长过程中通过光合作用不断吸收大气中的二氧化碳,外界大气中的二氧化碳含量的变化必然会影响到树轮中碳稳定同位素值的变化,利用树轮碳稳定同位素的研究是需要剔除大气二氧化碳的影响[8-9,12,26-28]。3个不同海拔高度岷江冷杉树轮δ13C下降趋势比较明显(见图2)。
树轮稳定同位素含有丰富的古气候信息,同时也包含了一部分非气候趋势和变化。在对原始数据进行处理,提取非气候信息的同时,保留真实的气候信号,最终得到用于气候分析的稳定碳同位素年表。经统计分析检验:岷江冷杉树轮δ13C序列与树轮年代呈非线性关系,本文采用二次多项式拟合与趋势比率法[29]消除大气δ13Ca对树轮δ13C测定结果的影响,二次多项式关系优于其它曲线关系[11]。拟合方程分别为:

式中,δ13Cx代表各海拔高度岷江冷杉树轮δ13C的拟合序列值,Y代表年。
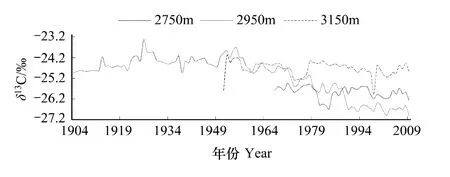
图2 不同海拔岷江冷杉树轮δ13C序列Fig.2 Tree-ring δ13CofAbiesfaxonianachronologyin different elevations

图3 用多项式与趋势比率法消除大气CO2影响后的不同海拔岷江冷杉树轮δ13C序列Fig.3 The Tree-ring δ13C of Abies faxoniana chronology after obviating the influence of atmospheric CO2by using polynomial and the Faller's method in different elevations
3.3 树轮δ13C序列与气候因子的关系
从表1可以看出,不同海拔岷江冷杉树轮δ13C序列的自相关系数均大于0.3,表明树木径向生长不仅与当年的气候因子(温度等)有关,而且受上年气候因子(温度、湿度、降水)的影响,即“滞后效应”,气候因子通过光合作用效率而影响δ13C,δ13C可以反映温度、湿度、降水等变化。选取上年1—12月及当年1—12月的气候因子与年表进行相关分析。气候因子包括温度、降水量及反映温度和降水量对树木生长综合影响的相对湿度,研究时段为器测资料以来的年代(1955—2009年)。岷江冷杉树轮δ13C比值序列(DS)与当年及前一年的各月的平均气温、月降水量及月平均相对湿度分别求相关。

表2 不同海拔岷江冷杉树轮δ13C比值序列(DS)与季节气候因子的关系Table2 Correlation function analysis between season-climate factors and tree-ring δ13C(DS)chronology of Abies faxoniana in different elevations
从图4可以看出,中海拔(海拔2950 m)的岷江冷杉树轮δ13C比值序列(DS)与上一年1月降水量呈显著正相关(P<0.05);而各海拔岷江冷杉树轮δ13C比值序列(DS)与降水量无显著相关性。中海拔区的岷江冷杉树轮δ13C比值序列(DS)与前一年6、12月和当年6月的月平均温度呈显著正相关(P<0.05),与前一年1、11月和当年1、2、11月的平均温度呈极显著正相关(P<0.01);低海拔(海拔2750 m)岷江冷杉树轮δ13C比值序列(DS)与当年12月月平均温度呈显著负相关(P<0.05);高海拔(海拔3150 m)的岷江冷杉树轮δ13C比值序列(DS)与当年4月月平均温度呈显著正相关(P<0.05)。低海拔的树轮δ13C比值序列(DS)与当年8月月平均相对湿度呈显著正相关(P<0.05);高海拔的树轮δ13C比值序列(DS)与上一年7、8月和当年4月月平均相对湿度呈显著负相关(P<0.05),尤其是前一年8月月平均相对湿度与树轮δ13C呈极显著负相关(P<0.01)。这是因为低海拔的岷江冷杉位于皮条河附近(30 m),更适生于冷湿及排水较好的环境,使得相对湿度促进了树轮碳稳定同位素的分馏作用,而随着海拔的升高温度降低,降水量也逐渐降低,使高海拔相对湿度也发生相应的变化,从而抑制了树轮碳稳定同位素的分馏作用。
3个不同海拔岷江冷杉树轮δ13C比值序列(DS)与季节气候因子进行相关分析见表2。从表2可看出,低海拔区的岷江冷杉树轮δ13C比值序列(DS)与夏季平均温度呈显著负相关(P<0.05),与夏季平均相对湿度呈显著正相关(P<0.05),而与降水量无显著相关性;中海拔区的岷江冷杉树轮δ13C比值序列(DS)与秋、冬季平均温度呈极显著正相关(P<0.01),与其它季节气候因子(降水量和相对湿度)无显著相关性;高海拔区的岷江冷杉树轮δ13C比值序列(DS)与季节气候因子无显著相关性。
为了更深入地了解气候因子对各海拔岷江冷杉树轮稳定碳同位素的影响程度,提取主要的控制因子,通过多元回归的方法来分析温度、降水和相对湿度对树轮稳定碳同位素的交互作用。得到多元回归方程如下:

图4 岷江冷杉树轮δ13C比值序列(DS)与气候因子的关系Fig.4 Correlation function analysis between climate factors and tree-ring δ13C(DS)chronology of Abies faxoniana
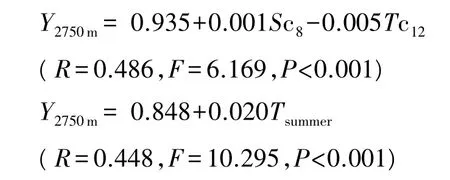
式中,Y2750m为低海拔区岷江冷杉树轮δ13C;Sc8为当年8月平均相对湿度;Tc12为当年12月月平均温度;Tsummer为夏季平均温度。
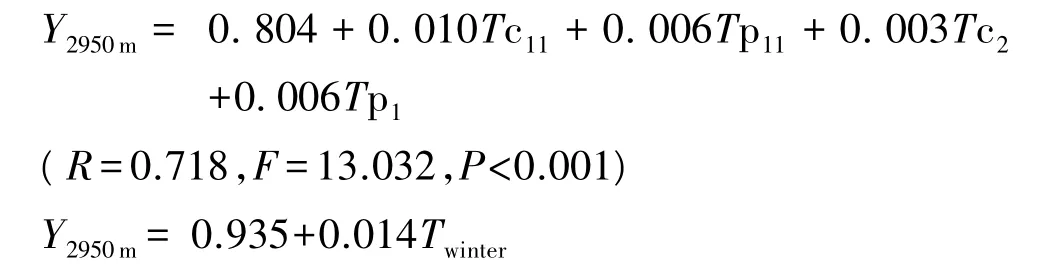

式中,Y2950m为中海拔区岷江冷杉树轮δ13C;Tp1、Tp11、Tc2、Tc11为分别上一年1、11月及当年2、11月月平均温度;Twinter为冬季平均温度。

式中,Y3150m为高海拔区岷江冷杉树轮δ13C;Sp8为上一年8月平均相对湿度;Tc4为当年4月月平均温度。
研究结果表明,低海拔区的岷江冷杉的树轮碳稳定同位素序列的主控因子是当年8月月平均相对湿度和当年12月月平均温度,夏季温度是该海拔岷江冷杉树木生长的限制因子;中海拔区的岷江冷杉的树轮碳稳定同位素序列的主控因子是上一年1、11月月平均温度和当年2、11月月平均温度,冬季温度是该海拔岷江冷杉树木生长的限制因子;高海拔区的岷江冷杉树木生长主要受上一年8月月平均相对湿度和当年4月月平均温度的影响。
5 结论与讨论
川西亚高山岷江冷杉树轮δ13C组成的变化范围为-23.33‰—-26.31‰,平均值为-24.91‰,变异系数为-0.011—-0.038,并表现出较强的一阶自相关,表明岷江冷杉树轮δ13C在年际变化中较为稳定,比天目山地区的柳杉树轮碳稳定同位素特征量要高[11]。岷江冷杉树轮δ13C序列在不同海拔高度之间存在着差异,没有表现出明显的海拔趋势,这与前人研究报道有一定的差异性[30]。自Korner等率先开展植物叶片δ13C值与海拔的关系以来[31],研究报道大多认为:随着海拔的增加,树轮δ13C值表现为逐渐变重的趋势,且与海拔高度的相关性弱[30-31]。而同一海拔高度的岷江冷杉树轮δ13C序列的变化趋势基本一致,对环境变化有较好的指示作用。某一年的树轮生长状况不仅与当年的气候因子有关,而且可能受上年气象因子的影响[6,8,12,32-35],这表明树轮δ13C序列与DS序列均存在较强的“滞后效应”,这种现象可能与当年的光合作用合成有机物的过程中,部分利用了储存在植物体内前一年光合作用合成的有机物有关[12]。
卧龙地区低海拔的岷江冷杉的树轮稳定碳同位素序列的主控因子是当年12月月平均温度,夏季温度是影响低海拔岷江冷杉树轮碳同位素分馏的主要因子,温度升高导致树木蒸腾作用加强,限制了树木的生长,故与树轮宽度呈负相关[36-38],这是因为12月温度的降低抑制岷江冷杉的径向生长,此时温度降低,使得光合作用的酶活性降低,抑制了光合作用的进行,形成窄年轮;中海拔的岷江冷杉的树轮稳定碳同位素序列的主控因子是上一年1、11月月平均温度和当年2、11月月平均温度,冬季温度是影响低海拔岷江冷杉树轮碳同位素分馏的主要因子,这是因为前一年1、6、11、12月和当年1、2、6、11月温度有助于岷江冷杉的径向生长,此时温度逐渐升高,达到了岷江冷杉最适温度,能更有效地吸收大气中的二氧化碳,有效的促进岷江冷杉的生命活动;高海拔的岷江冷杉树木生长主要受当年4月月平均温度的影响,这是因为当年4月温度有助于岷江冷杉的径向生长,此时温度逐渐升高,达到了岷江冷杉最适温度,能更有效地吸收大气中的二氧化碳,促进光合作用,从而促进岷江冷杉的生长。进入秋、冬季后,温度降低,光合作用速率减弱或停止,树木的生命活动即将停止或已停止,树木的呼吸和代谢需要消耗养分,对下一年树木径向生长有潜在影响;冬季温度的变化,对树木生命活动影响较大;当冬季温度偏暖时,树木的呼吸作用等生理活动加强,细胞内部CO2分压增大,树木年轮碳稳定同位素降低[11]。当冬季温度偏冷时,情况刚好相反。
低海拔区的岷江冷杉的树轮碳稳定同位素序列的主控因子是当年8月月平均相对湿度。低海拔区的岷江冷杉靠近皮条河,土壤相对湿度较大,岷江冷杉更适生于冷湿及排水较好的环境,使得相对湿度促进了树轮碳稳定同位素的分馏作用;中海拔的岷江冷杉树轮δ13C比值序列(DS)只与上一年1月降水量呈显著正相关(P≤0.05),降水的增加,使得光合作用效率增强,促进了岷江冷杉树木的生长;而其它各海拔岷江冷杉树轮δ13C比值序列(DS)与降水量无显著相关性。高海拔区的岷江冷杉树木生长主要受上一年8月月平均相对湿度的影响。这是由于上一年7、8月温度较高,降水量较大,使空气中相对湿度较大,对岷江冷杉树木光合作用有着抑制作用,同时呼吸作用增强,大量消耗树体营养物质,且岷江冷杉主要吸收利用浅层地下水,对雨水依赖性较小[39],因此降水不是该树木生长主要限制因子[6]。
卧龙自然保护区属于湿润、半湿润地区,亚高山暗针叶林中海拔区的降水充足,降水不是川西亚高山暗针叶林中海拔地区岷江冷杉生长的限制因子,岷江冷杉生理活动的正常进行须要有良好的温度条件。这与张同文等对阿勒泰西部地区年轮年表分析表明,树木生长与生长季温度呈正相关,与生长季降水呈负相关的结论基本一致[40],主要是由于阴雨天气持续不断,并会伴有大雾的出现,浓厚的云层会减少太阳光照的强度,植物接受的太阳辐射减少,光合作用速率降低,导致树轮δ13C较低;8月雨量较多,但光照强,温度高,蒸发大,干季水分胁迫可导致气孔关闭,导通度降低,从而引起岷江冷杉植物光合作用所固定的δ13C增大[11]。
研究表明,川西亚高山暗针叶林不同海拔高度的岷江冷杉的树木径向生长主要受到气温的制约,从生物学基础上阐明了树木生长与环境的关系;证明了在一些年轮宽度逐年差异不明显地区,利用树轮样本碳稳定同位素可以清楚地机械地了解气候对同化作用及木材形成过程中C传递过程和途径的影响;也进一步证明了建群种岷江冷杉对雨水的依赖很小,这有利于植物生存,且维持了川西亚高山暗针叶林群落的稳定性,同时,进一步说明了冬季温度的升高,有利于植物生长期的提前造成植物生长旺盛,抗旱能力减弱,影响着生态系统的稳定性。
本研究结果为重建岷江冷杉树木生长的历史环境单因子变化趋势及树轮碳稳定同位素序列变化趋势与环境关系的研究提供了理论分析依据,其研究结果有效地弥补了中国暖湿地区树轮δ13C稳定同位素研究的不足,深入揭示了在气候变化背景下,川西亚高山暗针叶林树木生长对气候因子变化(气候变暖、降水量减少等)的积极响应,为预测未来气候变化对岷江冷杉生长的影响具有参考价值。
[1] Loader N J,Robertson I,McCarroll D.Comparison of stable carbon isotope ratios in the whole wood,cellulose and lignin of oak tree-rings.Palaeogeography,Palaeoclimatology,Palaeoecology,2003,196(3/4):395-407.
[2] Liu Y,Wu X D,Leavitt S W,Hughes M K.Tree ring stable C Isotope and climate change in Huangling.Science in China(D),1996,26(2):125-130.
[3] Loader N J,McCarroll D,Gagen M,Robertson I,Jalkanen R.Extracting climatic information from stable isotopes in tree rings//Dawson T D,Siegwolf R.Stable Isotopes as Indicators of Ecological Change.London:Academic Press,2007:27-48.
[4] Ji C L,Xu Q,Jin X,Liu S R.Application of the tree-ring stable isotopes in study on the global climate change.Scientia Silvae Sinicae,2010,7(7):129-135.
[5] Liu X H,An W L,Liang E Y,Wang W Z,Shao X M,Huang L,Qin D H.Spatio-temporal variability and climatic significance of tree ring's δ13C of Picea crassifolia on the Qilian Mountains.Journal of Glaciology and Geocryology,2010,32(4):666-676.
[6] Jin X,Xu Q,Liu S R,Jiang C Q.Responses of the tree-ring Abies faxoniana and Tsuga chinensis to climate factors in subalpine in Western Sichuan.Scientia Silvae Sinicae,2013,49(1):21-26.
[7] McCarroll D,Loader N J.Stable isotopes in tree rings.Quaternary Science Reviews,2004,23(7/8):771-801.
[8] Leavitt S W,Treydte K,Liu Y.Environment in time and space:opportunities from tree-ring isotope networks//West J B,Bowen G J,Dawson T E,Tu K P,eds.Isoscapes:Understanding Movement,Pattern,andProcess onEarththrough Isotope Mapping.Springer,2010:113-115.
[9] Cernusak L A,Hutley L B.Stable isotopes reveal the contribution of corticular photosynthesis to growth in branches of eucalyptus miniata.Plant Physiology,2011,155(1):515-523.
[10] Li Z H,Liu R M,An Z S,Wu X D,Liu Y,Leavitt S W,Hughes M K.Tree-Ring δ13C seasonal changes and climatic significance.ChineseScienceBulletin,1995,40(22):2064-2064.
[11] Zhao X Y,Wang J,Qian J L,Jiang X Y.The climate change in autumn recorded in the δ13C of tree rings in the past 300 years at Tianmu Mountain.Journal of Mountain Science,2005,23(5):540-549.
[12] Zhang R B,Yuan Y J,Wei W T,Shang H M,Yu S L,Zhang T W,Chen F,Fan Z A,Qin L.Response of stable carbon isotope of larix sibirica ledeb.tree-rings to climate change.Arid Zone Research,2012,29(2):328-334.
[13] Jin X,Xu Q,Liu S R,Jiang C Q.Relationships of stable carbon isotope of Abies faxoniana tree-rings to climate in sub-alpine forest in Western Sichuan.Scientia Silvae Sinicae,2013,49(7):10-15.
[14] Peng J F,Gou X H,Chen F H,Liu P X,Zhang Y,Fang K Y.Characteristics of ring-width chronologies of Picea crassifolia and their responses to climate at different elevations in the Anyemaqen Mountains.Acta Ecologica Sinica,2007,27(8):3268-3276.
[15] Farquhar G D,O'Leary M H,Berry J A.On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves.Australian Journal of Plant Physiology,1982,9(2):121-137.
[16] Briffa K R,Schweingruber F H,Jones P D,Osborn T J,Shiyatov S G,Vaganov E A.Reduced sensitivity of recent tree-growth to temperature at high northern latitudes.Nature,1998,391(5):678-682.
[17] Deslauriers A,Rossi S,Anfodillo T,Saracino A.Cambial phenology,wood formation and temperature thresholds in two contrasting yearsathighaltitudeinsouthernItaly.Tree Physiology,2008,28(6):863-871.
[18] Shao X M,Fan J M.Indicates the tree-ring width data in the past climate change.Quaternary Research,1999,1(1):81-89.
[19] Qin N S,Shi X H,Shao X M,Wang Q C.Average Maximum temperature change recorded by tree rings in West Sichuan Plateau.Plateau and Mountain Meteorology Research,2008,28(4):18-24.
[20] Li Z S,Liu G H,Zhang Q B,Hu C J,Luo S Z,Liu X L,He F.Tree ring reconstruction of summer temperature variations over the past 159 years in Wolong National Natural Reserve,western Sichuan,China.Chinese Journal of Plant Ecology,2010,34(6):628-641.
[21] Buckley B U,Cook E R,Peterson M J,Barbetti M.A changing temperature responses with elevation for Largarostrobos franklinii in Tasmania Australia.Climatic Change,1997,36(3/4):477-498.
[22] Splechtna B E,Dobry J,Klinka K.Tree-ring characteristics of subalpine fir(Abies lasiocarpa(hook)Nutt.)in relation to elevation and climatic fluctuations.Annals of Forest Science,2000,57:89-100.
[23] Zhang Q B,Hebda R J.Variation in radial growth patterns of Pseudotsuga menziesii on the central coast of British Columbia,Canada.Canadian Journal of Forest Research,2004,34(9):1946-1954.
[24] Zheng S W,Li Y Q,He F,Chen H,Su Y M,Liu X L.Comparison analysis and research on principal meteorology factors inside&outside Wolong forest ecology station.Journal of Chengdu University:Natural Science Edition,2006,25(3):210-213.
[25] Xu Q,Jiang Y X,Liu S R,An S Q,Duan Z F.Study on the relationship between precipitation and river water at Balang Mountain Watershed in Wolong NatureReserveofSichuan Province.Forest Research,2007,20(3):297-301.
[26] Liu Y,Ma L M,Can Q F,An Z S,Liu W G,Gao L Y.Since the reconstruction of Helan mountain in 1890 using stable carbon isotopic tree-ring summer(6—8 months)temperature.Science in China(D),2002,32(8):667-674.
[27] Fan W Y,Wang X A.Advances in studies on relationships between tree-ring stable carbonisotope and climate change.Journal of Shaanxi Normal University:Natural Science Edition,2004,32(9):148-151.
[28] Shang H M,Wen W T,Yuan Y J,Yu S L,Zhang R B,Chen X J.Environmental response of tree-ring width and stable carbon isotope at North Slope of Eastern Tianshan Mountains.Desert and Oasis Meteorology,2010,4(5):6-10.
[29] Diao M X,Zhang X,Rao L C.Theoretical Statistics[M] .Beijing:China Science and Technology Press,1998.165-170.
[30] Chen T,Chen F H,An L Z,Liu X H.Variations of tree-ring and foliar δ13C values of Sabina przewalskii with altitude.Journal of glaciology and geocryology,2004,26(6):767-771.
[31] Korner C,Farquhar G D,Roksandic Z.A global survey of carbon isotope discrimination in plants from high altitude.Oecologia,1988,74(4):623-632.
[32] Liu X H,Qin D H,Shao X M,Ren J W,Wang Y.Stable Carbon isotope ofAbiesspectabibisfromNyingchiCountyofTibet Autonomous Region and its responds to climate change.Journal of Glaciology and Geocryology,2002,24(5):574-578.
[33] Deslauriers A,Giovannelli A,Rossi S,Castro G,Fragnelli G,Traversi L.Intra-annual cambial activity and carbon availability in stem of poplar.Tree Physiology,2009,29(10):1223-1235.
[34] Lü J,Tu Q P,Qian J L.The application of carbon isotopes in tree rings to meteorology.Meteorological Monthly,2001,27(1):9-13.
[35] Shang Z Y,Wang J,Zhang W,Li Y Y,Cui M X,Chen Z J,Zhao X Y.Vertical variability of Pinus sylvestris varmongolica tree ring δ13C and its relationship with tree ring width in northern Daxing'an Mountains of Northeast China.Chinese Journal of Applied Ecology,2013,24(1):1-9.
[36] Szeicz J M,MacDonald G M.Age-dependent tree-ring growth responses of subarctic white spruce to climate.Canada Journal of Forest Research,1994,24(1):120-132.
[37] Yadav R R,Park W k,Singh J,Dubey B.Do the Western Himalayas defy global warming?Geophysical Research Letters,2004,31(17):L17201-1-L17201-5,doi:10.1029/2004 GL020201.
[38] Gou X H,Chen F H,Yang M X,Jacoby G,Peng J F,Zhang Y X.A comparison of tree-ring records and glacier variations over the past700years,northeasternTibetanPlateau.Annalsof Glaciology,2006,43(1):86-90.
[39] Xu Q,Li H B,Chen J Q,Cheng X L,Liu S R,An S Q.Water use patterns of three species in subalpine forest,Southwest China:the deuterium isotope approach.Ecohydrology,2011,4(2):236-244.
[40] Zhang T W,Yuan Y J,Yu S L,Wei W T,Yang Q,Shang H M.June to September precipitation series of 1481—2004 reconstructed from tree-ring in the Western Region of Altay Prefecture,Xinjiang.Journal of Glaciology and Geocryology,2008,30(4):659-667.
参考文献:
[2] 刘禹,吴祥定,Leavitt S W,Hughes M K.黄陵树木年轮稳定C同位素与气候变化.中国科学(D辑),1996,26(2):125-130.
[4] 冀春雷,徐庆,靳翔,刘世荣.树木年轮碳氢氧稳定同位素在全球气候变化研究中的应用.林业科学,2010,7(7):129-135.
[5] 刘晓宏,安文玲,梁尔源,王文志,邵雪梅,黄磊,秦大河.祁连山青海云杉树轮δ13C的时空变化及其气候意义.冰川冻土,2010,32(4):666-676.
[6] 靳翔,徐庆,刘世荣,姜春前.川西亚高山岷江冷杉和铁杉年轮对气候因子的响应.林业科学,2013,49(1):21-26.
[10] 李正华,刘荣谟,安芷生,吴祥定,刘禹,Leavitt S W,Hughes M K.树木年轮δ13C季节性变化及其气候意义.科学通报,1995,40(22):2064-2064.
[11] 赵兴云,王建,钱君龙,姜修洋.天目山地区树轮δ13C记录的300多年的秋季气候变化.山地学报,2005,23(5):540-549.
[12] 张瑞波,袁玉江,魏文涛,尚华明,喻树龙,张同文,陈锋,范子昂,秦莉.西伯利亚落叶松树轮稳定碳同位素对气候的响应.干旱区研究,2012,29(2):328-334.
[13] 靳翔,徐庆,刘世荣,姜春前.川西亚高山森林岷江冷杉树轮稳定碳同位素对气候要素的响应.林业科学,2013,49(7):10-15.
[14] 彭剑峰,勾晓华,陈发虎,刘普幸,张永,方克艳.阿尼玛卿山地不同海拔青海云杉(Picea crassifolia)树轮生长特性及其对气候的响应.生态学报,2007,27(8):3268-3276.
[18] 邵雪梅,范金梅.树轮宽资料所指示的川西过去气候变化.第四纪研究,1999,1(1):81-89.
[19] 秦宁生,时兴合,邵雪梅,汪青春.川西高原树木年轮所指示的平均最高气温变化.高原山地气象研究,2008,28(4):18-24.
[20] 李宗善,刘国华,张齐兵,胡婵娟,罗淑政,刘兴良,何飞.利用树木年轮宽度资料重建川西卧龙地区过去159年夏季温度的变化.植物生态学报,2010,34(6):628-641.
[24] 郑绍伟,黎燕琼,何飞,陈泓,宿以明,刘兴良.卧龙森林生态站林内外主要气象因子对比分析研究.成都大学学报:自然科学版,2006,25(3):210-213.
[25] 徐庆,蒋有绪,刘世荣,安树青,段正峰.卧龙巴郎山流域大气降水与河水关系的研究.林业科学研究,2007,20(3):297-301.
[26] 刘禹,马利民,蔡秋芳,安芷生,刘卫国,高玲瑜.采用树轮稳定碳同位素重建贺兰山1890年以来夏季(6-8月)气温.中国科学(D辑),2002,32(8):667-674.
[27] 范玮熠,王孝安.树木年轮稳定碳同位素与气候变化的关系研究进展.陕西师范大学学报:自然科学版,2004,32(9):148-151.
[28] 尚华明,魏文寿,袁玉江,喻树龙,张瑞波,陈向军.天山北坡东部树轮宽度和稳定碳同位素的环境响应分析.沙漠与绿洲气象,2010,4(5):6-10.
[29] 刁明霞,张霞,饶良臣.理论统计学[M] .北京:中国科学技术出版社,1998,165-170.
[30] 陈拓,陈发虎,安黎哲,刘晓宏.不同海拔祁连园柏树轮和叶片δ13C值的变化.冰川冻土,2004,26(6):767-771.
[32] 刘晓宏,秦大河,邵雪梅,任贾文,王瑜.西藏林芝冷杉树轮稳定碳同位素对气候的响应.冰川冻土,2002,24(5):574-578.
[34] 吕军,屠其璞,钱君龙.树木年轮碳稳定同位素在气象中的应用.气象,2001,27(1):9-13.
[35] 商志远,王建,张文,李彦彦,崔明星,陈振举,赵兴云.大兴安岭北部樟子松树轮δ13C的高向变化及其树轮宽度的关系.应用生态学报,2013,24(1):1-9.
[40] 张同文,袁玉江,喻树龙,魏文寿,杨青,尚华明.树木年轮重建阿勒泰西部1481—2004年6—9月降水量序列.冰川冻土,2008,30(4):659-667.
猜你喜欢
中国农业科技导报(2024年1期)2024-04-29 00:42:04
山西林业科技(2022年4期)2022-03-12 04:39:48
广西农学报(2021年2期)2021-11-21 14:08:38
生态学报(2021年14期)2021-09-01 10:55:12
花火A(2019年1期)2019-03-22 02:27:42
现代养生·上半月(2017年7期)2017-07-07 21:43:25
环球人文地理·评论版(2016年6期)2016-08-23 01:08:48
西南农业学报(2016年6期)2016-04-16 05:13:01
湖北农业科学(2014年21期)2014-12-22 11:42:23
沙漠与绿洲气象(2014年3期)2014-03-25 13:11:50
