
杨树光合特性及叶片营养分布的空间异质性1)
2014-08-02 03:54:08井大炜邢尚军马海林杜振宇刘方春马丙尧
东北林业大学学报 2014年6期
井大炜 邢尚军 马海林 杜振宇 刘方春 马丙尧
(德州学院,德州,253023) (山东省林业科学研究院)
杨树光合特性及叶片营养分布的空间异质性1)
井大炜 邢尚军 马海林 杜振宇 刘方春 马丙尧
(德州学院,德州,253023) (山东省林业科学研究院)
以I-107欧美杨为研究对象,观测了不同冠层净光合速率的日变化、月变化及叶片叶绿素和营养元素质量分数的变化规律。结果表明:冠层I、冠层II和冠层III的净光合速率日变化均呈单峰型,且具有明显的分层性,其日均值的大小次序为:冠层III>冠层Ⅱ>冠层I,差异均达显著水平;同一冠层叶片的净光合速率在不同月份之间呈先增大后减小的趋势,7月和8月的净光合速率明显高于5月、6月和9月,同时随冠层的升高净光合速率呈递增的趋势,冠层Ⅱ和冠层III叶片净光合速率均显著高于冠层I;同一冠层4个方位叶片叶绿素质量分数的平均值表现为:冠层III≈冠层Ⅱ>冠层I,即冠层III和冠层Ⅱ未达到显著性差异,但均显著高于冠层I。此外,不同营养元素在杨树叶片中质量分数差异较大,其中氮质量分数较高,而磷元素、钾元素、钙元素和镁元素质量分数较低,并且随冠层的升高,各营养元素质量分数均呈递增的趋势。
I-107欧美杨;净光合速率;叶绿素质量分数;叶片营养;分层性
An experiment was conducted to determine the diurnal variation and monthly variation of net photosynthetic rate, and contents of chlorophyll and nutrient elements in different canopy. The diurnal variation of net photosynthetic rate in Canopy I, II, III is unimodal type and has obvious stratification. The diurnal average of net photosynthetic rate of the canopy is in an ascending order of Conopy I, II and III, and the differences among different canopy are significant. The leaf net photosynthetic rate in the same canopy displays the variation tendency of first increases and then decreases in different months, and the net photosynthetic rates in July and August are significantly higher than those in May, June and September. The net photosynthetic rate shows a trend of increasing with the increase of canopy, which the net photosynthetic rates of Canopy Ⅱ and III significantly increase compared with the Canopy I. The average value of leaf chlorophyll content for four different orientations in the same canopy shows the variation tendency: Canopy III≈Canopy II>Canopy I. The contents of different nutrient elements in poplar leaves have larger differences, so the nitrogen content is higher, and the elements of phosphorus, potassium, calcium and magnesium content are lower, and the contents of nutrient elements are increasing trend with the increase of canopy.
绿色植物的光合作用是森林生态系统固定碳的唯一途径和碳素循环过程的开始。因此,光合作用在植物界乃至全球生态系统的能量流动与物质循环中都具有非常重要的作用[1-2]。光合作用是植物生长发育的基础及生产力高低的决定性因素,同时又是一个对环境变化很敏感的生理过程[3]。树冠是林木与外界环境相互作用最直接和最活跃的界面层,由于光环境特征[4]和叶片氮质量分数[5]等因素的影响,光合特性在树冠的垂直层次与水平层次上存在着时间、空间上的差异性,并且树冠的结构和功能特性在不同的光环境下呈现出一定的可塑性[6]。
杨树具有适应性强,生长速度快和易繁殖成活的特点,再加上林木的质量轻、强度高、弹性好、纤维长且含量高、易加工等优点,在山东省已成为速生丰产林工程的主要造林树种[7]。通过修枝培育无节胶合板材,是提高杨树木材质量和经济效益的一项重要营林技术措施,但是目前对杨树修枝的研究仅限于不同修枝强度、修枝年龄、修枝季节等对树木生长的影响[8],而没有对不同冠层光合特性等问题进行系统的研究,因而所提出的修枝技术仍属经验性或推理性。此外,营养元素是植物营养和生长的物质基础,且作为植物体的组成成分,具有调节植物生理功能的作用。营养元素的供应状况、体内贮藏分布等既受周围环境和自身生长发育阶段的影响,也对植物的各种生理生化过程起一定的促进和干扰作用[9]。可见,营养元素对植物来说是非常重要的。目前,对杨树修枝技术的研究,尚没有学者从冠层光合特性及叶片营养元素质量分数变化的角度开展研究,而明确不同冠层在垂直方向上的光合特性及叶片营养元素的变化规律,能从机理上探讨杨树修枝的重要性和必要性,这对于科学合理的指导控冠具有举足轻重的意义。为此,本试验以欧美107杨为研究对象,从其光合特性的日变化、月变化及叶片营养元素质量分数等角度来探讨不同冠层光合作用的生理生态过程,为完善杨树的修枝技术、合理地进行冠层管理提供科学依据。
1 材料与方法
试验地点设在山东省济南市北郊林场,属暖温带大陆性季风气候区,四季分明,日照充分,年平均气温14 ℃,年平均降水量675 mm。供试土壤为潮土,土壤速效N 19.65 mg·kg-1,速效P为14.32 mg·kg-1,速效K为45.79 mg·kg-1,有机质质量分数为7.83 g·kg-1。试验林地每年施肥一次,所用化肥为尿素、磷酸一铵和氯化钾,肥料用量相当于N为205.28 kg·hm-2,P2O5为70.38 kg·hm-2和K2O为58.65 kg·hm-2的施肥水平。选择5年生I-107欧美杨为研究对象,株行距2.5 m×5.0 m,南北行向,林木生长均匀,平均树高12.75 m,平均胸径12.36 cm。
1.1 树冠的分层
在对30棵I-107欧美杨每木检尺的基础上,选出5株冠形完好的标准木,根据该品种杨树每年生长的枝条基本呈轮生状的特性,将造林第1年、第2年和第3年所生的轮生枝分别设为冠层I、冠层II和冠层III,并把每个冠层分为内部、中部和外部(完全不遮光部分为外部,中部和内部为遮光部分的二等分),同时结合4个方位(东、南、西、北)来测定不同冠层的光合特性及叶片营养指标。
1.2 叶片光合作用及叶绿素质量分数的测定
2013年5—9月,选择晴朗的天气,在树旁架10 m高的人字梯,利用英国产的ADC Lci型便携式光合作用仪测定杨树冠层各部分枝条(内—中—外)不同部位(东、南、西、北)成熟叶片的净光合速率,并于2013年8月26日(晴天)的08:00—18:00每隔2 h测定一次叶片的净光合速率(日变化),同时在相应部位选取有代表性的叶片迅速带回实验室采用80%丙酮浸提,分光光度计比色法测定叶绿素质量分数[10],重复3次。为保持其它环境因子稳定且适宜,将叶室温度设置为25 ℃,相对湿度设为60%。
然后,净光合速率的日变化与月变化分别结合不同部位(内—中—外)的净光合速率和不同部位的叶片数采用加权平均公式计算。

式中:Pni为不同冠层的净光合速率,i为冠层I、II、III,Pni-外、Pni-中、Pni-内分别为枝条外部、中部和内部对应的净光合速率,ni-外、ni-中、ni-内分别为枝条外部、中部和内部对应的叶片数。
净光合速率日变化与月变化的测量方法及仪器参数的设置均保持一致。此外,不同冠层的叶绿素质量分数也采用类似的加权平均公式来计算。
1.3 叶片营养元素质量分数的测定
2013年5—9月分别采用高枝剪分冠层(I、Ⅱ、III)、部位(内—中—外)和方位(东、南、西、北)随机剪取枝条中成熟的叶片,分开处理保存,在杀青、烘干后分别采用H2SO4-H2O2-蒸馏法、钒钼黄比色法、火焰光度计法测定其N、P和K质量分数,并用原子吸收分光光度计测定钙、镁质量分数[11]。
最后结合不同月份营养元素质量分数的平均值和不同部位的叶片数采用加权平均公式计算。
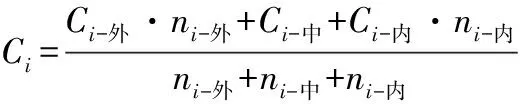
式中:Ci为冠层不同方位(东、南、西、北)的各营养元素质量分数,i为冠层I、II、III,Ci-外、Ci-中、Ci-内分别为枝条外部、中部和内部对应的各营养元素质量分数,ni-外、ni-中、ni-内分别为枝条外部、中部和内部对应的叶片数。
1.4 数据分析
采用SPSS17.0统计软件进行数据统计,利用Excel2007和Sigmaplot10.0分别进行数据计算和图形制作。
2 结果与分析
2.1 不同冠层Pn的日变化
表1显示了杨树不同冠层净光合速率的日变化,可知,3个冠层叶片净光合速率的日变化趋势基本一致,从08:00至12:00,冠层I、II和III的净光合速率随着光强的增加而增大,12:00时达到最大,分别为9.89、13.43和15.97μmol·m-2·s-1,冠层III分别比冠层I和II提高61.48%和18.91%,差异均达显著水平。从12:00之后,各冠层的净光合速率均呈下降的趋势,在18:00时达到最小值。从08:00至18:00,3个冠层净光合速率的平均值分别为5.30、7.11和9.49μmol·m-2·s-1,差异亦达到显著水平。由此可见,3个冠层的净光合速率日变化均呈单峰型,并且具有明显的分层性,其中大到小变化规律为冠层III、冠层Ⅱ、冠层I。
由表2可见,杨树不同冠层光合有效辐射的日变化表现为单峰曲线;从08:00至18:00,光合有效辐射呈先增大后减小的变化趋势,在12:00时达到最大值。方差分析表明,在08:00时,冠层III的光合有效辐射显著高于冠层I和II,而冠层I和II差异不显著;从10:00至14:00,不同冠层光合有效辐射的大小次序为:冠层III、冠层II、冠层I,差异均达显著水平;在18:00时,冠层I、II和III之间均无显著性差异。从日变化的平均值来看,冠层III的光合有效辐射最大,其次是冠层II,冠层I最小,且冠层之间的差异均达到显著水平。由此可见,净光合速率与光合有效辐射的日变化规律基本保持一致。
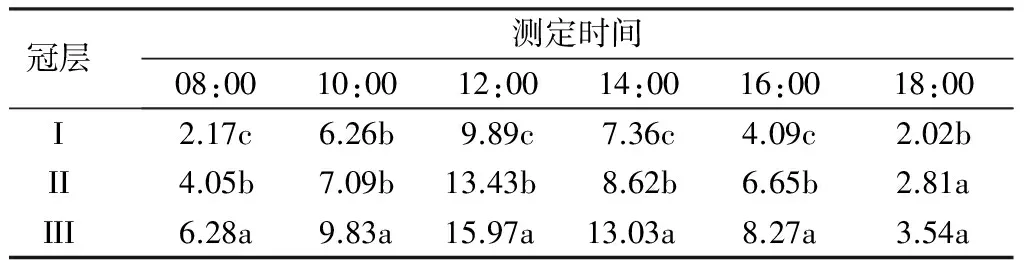
表1 杨树不同冠层净光合速率的日变化μmol·m-2·s-1
注:同列不同小写字母表示处理间差异显著(P<0.05)。
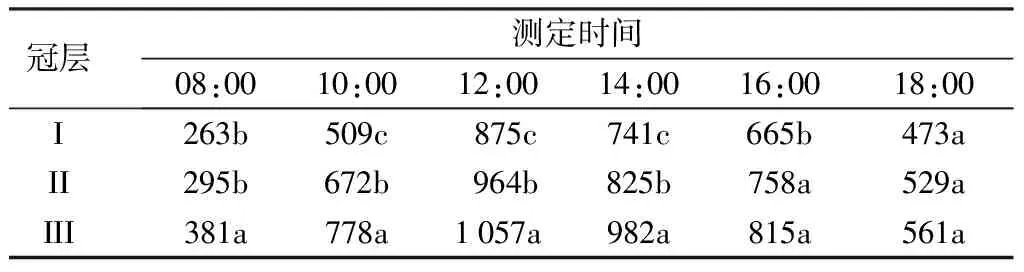
表2 杨树不同冠层光合有效辐射的日变化 μmol·m-2·s-1
注:同列不同小写字母表示处理间差异显著(P<0.05)。
2.2 不同冠层Pn的月变化
杨树不同冠层净光合速率的月变化如表3所示。可以看出,同一冠层叶片的净光合速率在不同月份间差异均达到显著水平,3个冠层叶片净光合速率在5月时最低,随后逐渐增大并在8月时达到峰值,之后即下降。叶片净光合速率随冠层的升高而增大,冠层I、Ⅱ和III叶片净光合速率的平均值分别为7.78、10.65和12.32 μmol·m-2·s-1,冠层III分别比冠层I和Ⅱ提高58.27%和15.66%,差异均达到显著水平。数据表明,同一冠层叶片的净光合速率在不同月份之间呈先增大后减小的趋势,7月和8月的净光合速率明显高于5、6和9月;同时随冠层的升高净光合速率呈递增的趋势,冠层Ⅱ和冠层III叶片净光合速率均显著高于冠层I。
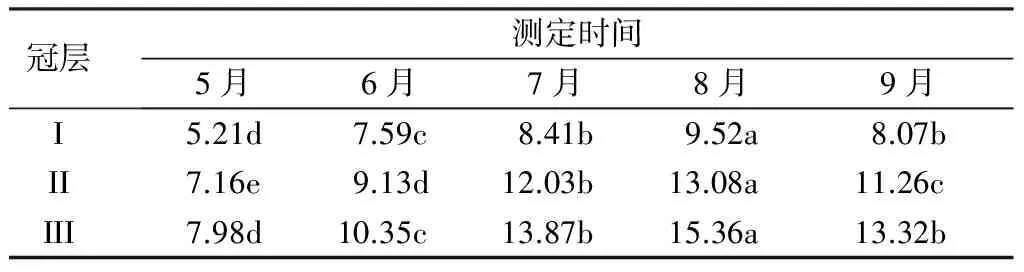
表3 杨树不同冠层净光合速率的月变化 μmol·m-2·s-1
注:同列不同小写字母表示处理间差异显著(P<0.05)。
2.3 不同冠层的叶绿素质量分数
叶片是光合作用的主要器官,叶绿体是进行光合作用的主要细胞器[2],而叶绿素是参与光合作用光能吸收、传递和转化的重要色素,因此叶片的光合速率和叶绿素质量分数有密切关系。由表4可知,不同方位叶片的叶绿素质量分数在冠层间变化趋势不完全一致。冠层I不同方位叶片的叶绿素质量分数无显著性差异,变化幅度为1.25~1.58 mg·g-1;冠层II不同方位叶片叶绿素质量分数的变化幅度为2.32~2.72 mg·g-1,从大到小次序为:南、东、西≈北,南方位的叶绿素质量分数最高,并显著高于其它方位;其次是东方位,也显著高于西和北方位,而西方位和北方位差异不显著。冠层III叶片叶绿素质量分数的变化幅度为2.45~2.93 mg·g-1,不同方位的变化规律与冠层II基本一致。同一冠层4个方位叶片叶绿素质量分数的平均值从大到小依次为冠层III≈冠层Ⅱ、冠层I,冠层III和冠层Ⅱ未达到显著性差异,但均显著高于冠层I。可见,最下层叶片的叶绿素质量分数最低,这与净光合速率的变化趋势基本一致。
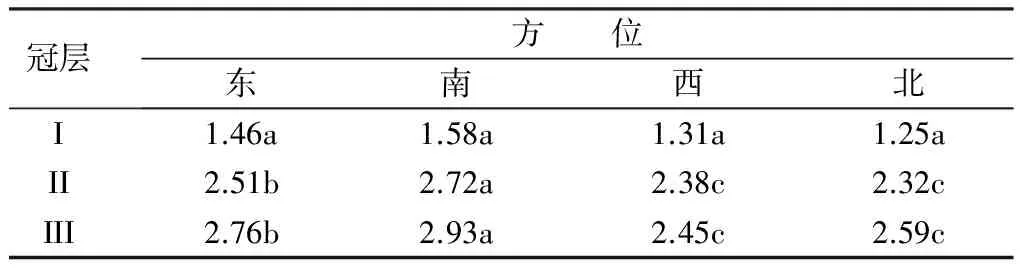
表4 杨树不同冠层不同方位的叶绿素质量分数 mg·g-1
注:同列不同小写字母表示处理间差异显著(P<0.05)。
2.4 不同冠层的叶片营养
氮是叶绿素、核酸、酶、维生素和生物碱等的主要成分;磷是核酸和核苷酸的组成部分,也是原生质和细胞核的主要成分;钾不仅提高光合作用的强度和增强抗逆性,而且对植物体内糖类的形成和运输、氮代谢等也有影响[12]。不同冠层叶片营养元素质量分数如表5所示。不同营养元素在叶片中质量分数差异较大,其中氮质量分数较高,而磷元素、钾元素、钙元素和镁元素质量分数较低。就氮元素而言,4个不同方位的叶片氮质量分数及其平均值从冠层I到冠层III呈上升趋势:冠层III>冠层Ⅱ>冠层I,即氮质量分数随树冠位置的上升而增加,且冠层间差异均达到显著水平。同一冠层不同方位之间的氮质量分数也有差异,但未达到显著水平。
冠层I、冠层Ⅱ、冠层III叶片磷质量分数的平均值分别为1.64、2.28和2.43 g·kg-1,叶片钾质量分数的平均值分别为1.50、2.10和2.26 g·kg-1,且各冠层之间的差异均达显著水平。同叶片氮质量分数相比,叶片磷和钾质量分数均有明显的下降,并且各冠层之间的差异也在减小。同一冠层不同方位的磷和钾质量分数也有差异,但未表现出明显的规律性。
钙元素和镁元素在不同冠层间呈现出基本一致的变化规律:冠层Ⅲ≈冠层Ⅱ>冠层I,即冠层Ⅲ和冠层Ⅱ的钙、镁质量分数均差异不显著,但显著高于冠层I。总体而言,杨树不同冠层之间的营养元素质量分数具有明显的分层性,随着冠层的升高,各营养元素质量分数均呈递增的趋势。
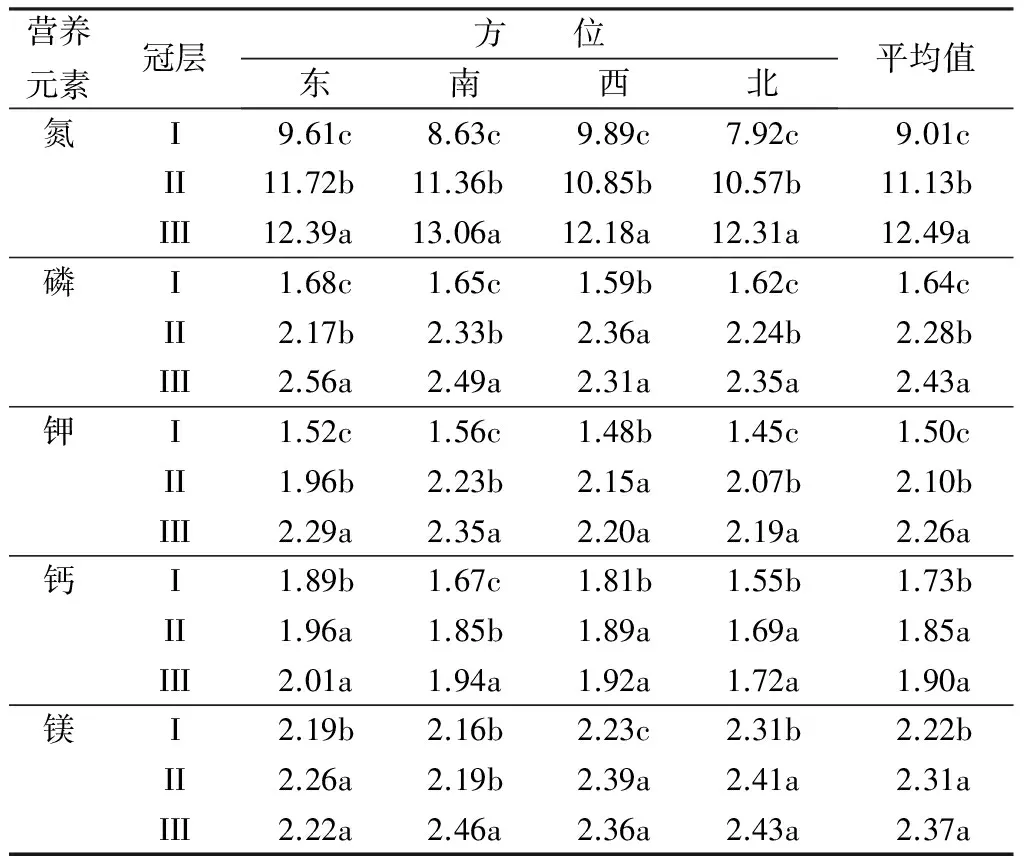
表5 杨树不同冠层不同方位的叶片营养状况
注:同列不同小写字母表示处理间差异显著(P<0.05)。
3 结论与讨论
树冠是树木进行光合作用、制造干物质的载体,增加产量的基础就是增强树冠截获有效太阳辐射的能力和提高光能转化效率。由于树冠结构和几何特征对太阳辐射和降水的影响,使能量传输和分配呈现冠层的空间异质性,导致光合作用在冠层空间上存在差异[13-14]。本试验研究发现,杨树不同冠层净光合速率日变化的规律为:冠层III>冠层Ⅱ>冠层I,表明冠层叶片的光合能力由强到弱依次为冠层III、冠层Ⅱ、冠层I。同时通过对净光合速率的月变化分析得出,7月和8月的净光合速率明显高于5月、6月和9月,并且不同冠层之间具有明显的分层性:冠层III>冠层Ⅱ>冠层I。这与张小全等[15]对杉木针叶光合特性的季节变化规律基本一致。同时叶绿素质量分数在不同冠层间呈现出与净光合速率一致的变化趋势,这与马永春[16]在I-69杨上的研究结果一致。
植物的化学成分是反映植物在一定生境下积蓄营养元素或从土壤中吸取营养元素的能力,也受树种的分类学地位和遗传性控制[2,17]。梁智等[18]等在核桃树上的研究表明,核桃树对氮、钾、钙的需求较多,对磷、镁的需求较少。也有人对马占相思树的研究发现[19],各器官中的大量营养元素质量分数以氮最高,由大到小次序为:氮、钙、钾、镁、磷。本研究得出,杨树在不同冠层叶片的营养元素均以氮素质量分数为最高,而磷、钾、钙和镁质量分数较低,这可能与欧美107杨对不同营养元素的需求特性有关。本试验还得出,随着树冠的升高,各营养元素质量分数总体呈递增的趋势,表明叶片的营养元素质量分数在垂直方向上具有明显的分层性。这与刘广全和土晓宁[20]在栎林叶片上的研究结果一致。这可能是由于杨树叶片的营养元素质量分数与叶片的净光合速率、蒸腾速率及空气相对湿度密切相关,而这些因素在垂直方向上具有明显的成层性所致。
马永春[16]也对杨树不同冠层叶片的营养元素质量分数进行了研究,但在每个冠层中只选择了有代表性的两片叶子,具有一定的偶然性和片面性。而本研究把每个冠层分为“内—中—外”3个部分,结合不同部分的叶片数采用加权平均的方法来计算各个冠层的相关指标,相比而言能更全面、系统地描述不同冠层的光合特性及营养状况,结果更具有代表性和说服力。通过本研究得出,冠层I的光合特性最弱,并且营养元素质量分数也是最低,这表明下部冠层的枝条达到一定树龄后,该冠层叶片的整体光合生产能力会显著下降。所以建议对杨树进行修枝时,应该以冠层为对象,然后逐年从下往上修掉达到一定树龄、叶片光合生产能力显著下降的冠层的枝条。
[1] Boonen C, Samson R, Janssens K, et al. Scaling the spatial distribution of photosynthesisfrom leaf to canopy in a plant growth chamber[J]. Ecological Modelling,2002,156(2/3):201-212.
[2] 潘瑞炽.植物生理学[M].6版.北京:高等教育出版社,2008.
[3] 彭晓邦,仲崇高,沈平,等.玉米大豆对农林复合系统小气候的光合响应[J].生态学报,2010,30(3):710-716.
[4] 肖文发.杉木人工林单叶至冠层光合作用的扩展与模拟研究[J].生态学报,1998,18(6):621-628.
[5] Hikosaka K, Hirose T. Nitrogen uptake and use by competing individuals in a Xanthium canadense stand[J]. Oecologia,2001,126(2):174-181.
[6] Vanderklein D W, Wilens R T, Cartier A, et al. Plant architecture and leaf damage in bear oak. I: physiological responses[J]. North-eastern Naturalist,2004,11(3):343-356.
[7] 郑世锴.杨树丰产栽培[M].北京:金盾出版社,2005.
[8] 方升佐,徐锡增,严相进,等.修枝强度和季节对杨树人工林生长的影响[J].南京林业大学学报,2000,24(6):6-10.
[9] 操国兴,谢德体.多枝桉叶营养元素质量分数的季节变化与施肥效应研究(Ⅱ)[J].生物学杂志,2003,20(4):17-19.
[10] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[11] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:24-214.
[12] 方升佐,徐锡增,吕士行.杨树定向培育[M].合肥:安徽科学技术出版社,2004.
[13] Anten N P R. Optimal photosynthetic characteristics of individual plants in vegetation stands and implications for species coexistence[J]. Annals of Botany,2005,95(3):495-506.
[14] 郑元,赵忠,周慧,等.刺槐树冠光合作用的空间异质性[J].生态学报,2010,30(23):6399-6408.
[15] 张小全,徐德应.杉木中龄林不同部位和叶龄针叶光合特性的日变化和季节变化[J].林业科学,2000,36(3):19-26.
[16] 马永春.杨树人工林修枝机理及修枝技术体系的研究[D].南京:南京林业大学,2012.
[17] Munson A D, Margolis H A, Brand D G. Seasonal nutrient dynamics in white pine and white spruce in response to environmental manipulation[J]. Tree Physiology,1995,15(3):141-149.
[18] 梁智,周勃,邹耀湘.核桃树体生物量构成及矿质营养元素累积特性研究[J].果树学报,2012,29(1):139-142.
[19] 温中林,郑卫,陈日美.马占相思人工林生物量及营养元素分布研究[J].安徽农业科学,2010,38(27):15040-15045.
[20] 刘广全,土晓宁.秦岭锐齿栎林叶内营养元素质量分数的时空分布[J].西北林学院学报,1999,14(4):1-8.
Spatial Heterogeneity of Photosynthesis Characteristics and Leaf Nutrition Distribution in Poplar/
Jing Dawei(Dezhou University, Dezhou 253023, P. R. China); Xing Shangjun, Ma Hailin, Du Zhenyu, Liu Fangchun, Ma Bingyao(Shandong Forestry Academy)//
Journal of Northeast Forestry University.-2014,42(6).-34~37
Populus×euramericanacv. ‘Neva’; Net photosynthetic rate; Chlorophyll content; Leaf nutrition; Stratification
井大炜,男,1982年1月生,德州学院,博士研究生。
邢尚军,男,山东省林业科学研究院,研究员。E-mail:xingsj-126@126.com。
2014年1月22日。
S792.11;Q945.78
1) 山东省农业重大应用技术创新课题资助。
责任编辑:潘 华。
猜你喜欢
中国农业信息(2022年1期)2022-05-25 13:31:46
农业机械学报(2021年11期)2021-12-07 05:36:44
大气科学(2021年1期)2021-04-16 07:34:18
现代园艺(2017年23期)2018-01-18 06:57:44
新课程·下旬(2017年7期)2017-08-14 11:12:50
农业环境科学学报(2017年2期)2017-03-20 14:57:37
分析测试学报(2015年11期)2015-02-28 16:41:15
西北林学院学报(2015年5期)2015-01-03 05:12:20
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
植物营养与肥料学报(2011年3期)2011-10-24 06:14:44
