
HPV16 E6/E7对凋亡调控基因表达影响的研究
2014-07-13 07:37沙亚哈提别尔克哈之吉别克瓦提别克刘伊宁李晓苗来雯婷美丽吾尔提达吾列提汗谌宏鸣李惠武
新疆医科大学学报 2014年11期
沙亚哈提·别尔克哈之, 李 卉, 吉别克·瓦提别克, 刘伊宁, 李晓苗, 来雯婷,美丽吾尔提·达吾列提汗, 谌宏鸣, 李惠武
(新疆医科大学基础医学院, 乌鲁木齐 830011)
目前,越来越多的证据表明人乳头状癌病毒(Human papilloma virus,HPV)的感染与食管鳞癌的发生有关。Guo等[1]对安阳300例食管癌患者和900例正常人的食管组织进行研究,结果表明HPV的感染与食管癌的发生关系非常密切,这为HPV感染是中国食管癌高发区病因提供了有力的证据,并且指出HPV16的感染与食管癌的关系最为密切。Lu等[2]将HPV16 E6E7蛋白融合疫苗注入HPV16 E7表达的食管鳞癌细胞株中,结果明显降低了食管癌细胞的生长增殖。这一结果从另一方面证明HPV16 在食管癌的发生、发展中起到重要作用。但尚无HPV E6/E7与凋亡调控因子关系的研究报道。
HPV16 是一类小型无胞膜的环状双链DNA病毒,具有高度的宿主特异性,其早期表达的2个蛋白E6和E7扰乱正常细胞周期活动和信号通路。Longworth等[3-4]报道,高风险E7通过锌指状区域蛋白序列与组蛋白脱乙酰酶(HDACs)结合并直接调节病毒复制。作为促凋亡基因Bad 含有BH3结构域的前凋亡蛋白,与Bcl-2抗凋亡蛋白形成二聚体,调控细胞凋亡。Bcl-2基因主要通过抑制后者发挥抗凋亡作用,是Bcl-2家族抗凋亡蛋白中的一员。本实验通过用课题组前期构建的 HPV16 E6 及E6E7 融合基因的真核表达载体,转染食管癌细胞 KYSE450细胞,过表达E6 及E6E7,检测转录水平上Bcl-2和Bad 的表达变化,进一步探讨HPV16 E6/E7对凋亡调控因子Bcl-2和Bad表达的影响,以及凋亡调控因子与HPV的联系。
1 材料和方法
1.1细胞株和培养基食管癌细胞株KYSE450购自中国科学院细胞中心,小牛血清购自杭州四季青生物工程材料有限公司,RPMI1640 培养基和胰蛋白酶购自美国GIBCO公司,DMSO(二甲基亚砜)购自SIGMA公司,HDAC Inhibitor购自美国SIGMA公司。
1.2细胞的培养食管癌细胞株KYSE450使用RPMI1640培养基(含10%胎牛血清及青霉素、链霉素和两性霉素各 100 U/mL),置 37℃、饱和湿度、5%CO2孵育箱内培养,隔天观察。脂质体转染食管癌细胞KYSE450:实验共分正常培养组(C)、空载体pLVX-AcGFP1-N1转染组(P)、空载体pLVX-AcGFP1-N1转染抑制组(iP)、pLVX-AcGFP1-N1-E6转染组(iE6)及pLVX-AcGFP1-N1-E6E7融合基因转染组(E6E7)5组。12孔板内培养细胞至80%融合度时,用低血清培养基洗涤细胞,以 4 μL LipofectamineT M 2000转染 1.6 μg质粒 DNA, 6 h后更换含10%胎牛血清的1640培养基,同时对于转染组用2 nm/L的TSA的培养液培养,孵箱内继续孵育48 h。
1.3试剂和仪器Trizol、异丙醇、溴化乙锭(Sigma公司),M-MLV cDNA合成试剂盒(Promega公司),PCR均用试剂盒、DNA Marker、6×Loading buffer、琼脂糖、50×TAE(上海生工生物技术有限公司),其他试剂为国产分析纯。
opti-MEM®I 低血清培养基(美国 Invitrogen公司),低温高速离心机(Beckman CoulterTM,美国),梯度PCR仪( iCyclerTM Thermal Cycler,美国),凝胶成像系统(Bio-Rad,美国),生物安全柜(HERA safe,德国),电热恒温培养箱(上海华达器材厂)。
1.4RT-PCR检测及结果判定
1.4.1 总RNA提取 按Trizol一步法提取RNA,紫外分光光度计测定各组RNA浓度和A260/280比值,RNA保存于-80℃。
1.4.2 引物设计与合成 根据Genebank查找 Bad与Bcl-2序列,利用Primer 5.0软件设计其引物,引物序列如下:Bad上游引物:5′-CAGAGTTTGAGCCGAGTGAGC-3′,下游引物:5′-CCCATCCCTTCGTCGTCCT-3′,片段长度为247 bp。Bcl-2 上游引物:5′-CGCTGGGAGAACAGGGTA-3,下游引物:5′-GGGCTGGGAGGAGAAGAT-3′,片段长度为151 bp。GAPDH 上游引物:5′-GGGAAACTGTGGCGTGAT-3′,下游引物:5′-AAAGGTGGAGGAGTGGGT-3′,片段长度为309 bp。
1.4.3 反转录合成cDNA 总RNA 1 μL、oligo(dT)引物1 μL、RNase-free Water 9 μL混匀后置于72℃ 10 min,取出后置于冰上逐次加入MgCl24 μL、dNTP 2 μL、10倍反应缓冲液2 μL、RNA酶抑制剂0.5 μL、逆转录酶0.5 μL,最终体积达到20 μL。混匀后置于42℃ 1 h,用95℃ 5 min终止反应。取出后置冰上,加80 μL双蒸水,得到100 μL逆转录产物,于-20℃冰箱保存备用。
1.4.4 RT-PCR扩增反应 逆转录产物cDNA模板2 μL,待扩增的各基因上下引物各1 μL,即用PCR反应试剂盒反应溶液10 μL,ddH2O 6 μL,总体积20 μL。
1.4.5 RT-PCR反应条件 用cDNA为模板,以GAPDH作为内参对照,进行PCR扩增,反应体系20 μL:上下游引物(10 μmol/L)各1.0 μL,模板 cDNA 2.0 μL,ddH2O 6.0 μL,Mg2+0.4 μL,退火温度60℃,35个循环。
目的基因的扩增: 取2×Taq PCR Master Mix 10 μL,向其中加入目的基因正向和反向引物各1 μL、6 μL ddH2O,再加入2 μL的 cDNA模板,最终的PCR反应体系为20 μL,Bcl-2与Bad 的退火温度分别为58℃和60℃。
1.4.6 PCR产物凝胶电泳 10 μL产物加入1 μL 5×Loading Buffer上样缓冲液,经2%的琼脂糖凝胶电泳后,在紫外光凝胶成像分析仪下分析结果,用内参照光密度值标化Bad、Bcl-2的吸光度值,得到目的基因表达的相对含量。

2 结果
2.1Bad和Bcl-2基因的表达Bad mRNA在食管癌细胞KYSE450中表达较低,E6E7过表达对细胞内Bad 基因 mRNA水平无影响,各转染组之间无差异。Bcl-2 mRNA 在正常组(C)和空载体组(P)中均有表达,E6E7融合基因转染组(E6E7)的表达量高于其他各组,E6/E7对Bcl-2有轻度上调作用(表1)。
2.2HDAC抑制剂(TSA)对E6调控Bcl-2基因表达的影响经过HDAC抑制剂TSA处理后,尽管Bcl-2基因的表达水平受到了抑制,但TSA却明显增加了E6对Bcl-2基因表达的诱导调控作用(表1、图1)。

图1 转染KYSE450细胞后目的基因的检测
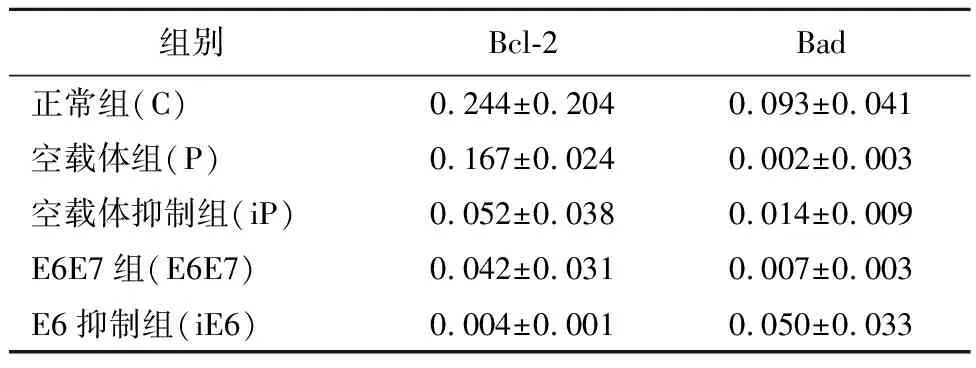
组别Bcl-2Bad正常组(C)0.244±0.2040.093±0.041空载体组(P)0.167±0.0240.002±0.003空载体抑制组(iP)0.052±0.0380.014±0.009E6E7组(E6E7)0.042±0.0310.007±0.003E6抑制组(iE6)0.004±0.0010.050±0.033
3 讨论
肿瘤的发生是一个复杂的过程,伴随多种可能的机制。HPV16早期表达基因可能参与细胞周期的肿瘤发生、发展过程,但对于E6/E7是否对肿瘤细胞凋亡过程进行调控尚未见报道。HPVs 是一类小型无胞膜的环状双链DNA病毒,具有高度的宿主特异性,并对鳞状上皮细胞极具感染力[5-6]。最近的研究发现抑制HPV E6和E7致癌基因可引起细胞凋亡,并激活肿瘤细胞抑制通路[7]。Bad蛋白可能通过与凋亡抑制基因Bcl-2和Bcl-xl等的表达产物形成异源二聚体而达到促凋亡效果[8-9]。Bcl-2基因是Bcl-2基因家族的一个重要成员, Bcl-2可以通过阻止caspase的活化在线粒体细胞色素C释放的上游发挥其抗凋亡的作用[10-11]。研究发现,Bcl-2在肾脏肿瘤、胃肠肿瘤中高表达,细胞凋亡受到抑制,加速肿瘤生长,预后差[12-13]。Schimmer等[14]研究发现,在Bcl-2基因和Bcl-xl基因过表达的情况下,Bad蛋白可直接诱导凋亡,具有治疗意义。Hattori等[15]认为,促凋亡的Bad蛋白与抗凋亡的Bcl-2蛋白竞争性结合形成异二聚体调节凋亡过程。
HDAC抑制剂是一类新型抗肿瘤药物,其通过改变染色质结构而调控基因表达。HDAC抑制剂的主要生物效应包括诱导肿瘤细胞分化、细胞周期阻滞和细胞凋亡、增强化疗和放疗敏感性、逆转转移性肿瘤的恶性表型等抗肿瘤效应[16-17]。本实验对食管癌KYSE450细胞中Bad mRNA水平进行检测,结果显示Bad mRNA水平在 E6 载体转染组、E6E7 融合基因载体转染组、正常培养组和空载体组间没有明显变化。推测该食管癌细胞中,E6、E7 与Bad之间在转录水平上可能不存在调控关系,Bcl-2 mRNA 在正常培养组和空载体转染组中均有表达, E6E7融合基因转染组的表达量高于其他各组,说明E6/E7对Bcl-2有轻度调控作用,使该基因表达增高, 这与目前研究报道一致,本研究中,E6/E7的表达促进Bcl-2 在转录水平上调,但具体机制还有待进一步验证。 HDAC抑制剂 TSA处理后,尽管Bcl-2基因的表达受到了抑制,但TSA却明显增加了E6对Bcl-2基因表达的诱导调控作用,提示HDAC在细胞中E6与Bcl-2在转录水平上存在调控关系。
综上所述,病毒感染后早期表达致癌基因 E6、E7 可能刺激肿瘤转移因子 Bcl-2 在转录水平上的表达,促进了肿瘤的恶性发展。Bcl-2作为主要的调亡调控因子,在肿瘤的生物学行为中起着重要的作用。进一步研究E6、E7调控Bcl-2在肿瘤发生、发展中的作用机制,对肿瘤早期诊断和临床治疗具有有意义的价值。
参考文献:
[1] Guo FC, Liu Y, Wang XE, et al. Human papillomavirus infection and esophageal squamous cell carcinoma:a Case-Control study[J]. Cancer Epidemiol Biomarkers Prev, 2012, 21(5):780-785.
[2] Lu YZ, Zhang ZX, Liu Q, et al. Immunological protection against HPV16 E7-expressing human esophageal Cancer cell challenge by a novel HPV16-E6/E7 fusion protein based-vaccine in a Hu-PBL-SCID mouse model[J]. Biol Pharm Bull, 2007, 30(1):150-156.
[3] Longworth MS, Laimins LA.The binding of histone deacety-lases and integrity of Zinc finger-like motifs of the E7 protein are essential for the Life cycle of human papillomavirus type 31[J]. J Virol, 2004, 78:3533-3541.
[4] Longworth MS. Regina wilson and laimonis a laimins[J]. EMBO J, 2005, 24:1821-1830.
[5] Hausen H.Papillomaviruses in the causation of human cancers-a brief historical account[J]. Virology, 2009, 384(2):260-265.
[6] Moody CA,Laimins LA.Human papillomavirus oncoproteins: pathways to transformation[J]. Nat Rev Cancer, 2010, 10(8):550-560.
[7] Magaldi TG, Almstead LL, Bellone SA, et al. Primary human cervical carcinoma cells require human papillomavirus E6 and E7 expression for ongoing proliferation[J]. Virology, 2012, 422(1):114-124.
[8] Zha J, Harada H, Yang E, et al. Korsmeyer SJ serine phosphorylation of death agonist BAD in response to survival factor results in binding to 14-3-3 not BCL-X(L)[J]. Cell,1996,87:619-628.
[9] Datta SR, Katsov A, Hu L, et al. 14-3-3 proteins and survival kinases cooperate to inactivate BAD by BH3 domain phosphorylation[J]. Mol Cell, 2000, 6:41-51.
[10] Burlacu A. Regulation of apoptosis by Bcl-2 family proteins[J]. Cell Mol Med, 2003, 7:249-257.
[11] Sprick MR, Walczak H. The interplay between the Bcl-2 family and death receptor-mediated apoptosis[J]. Biochim Biophys Acta, 2004, 1644(2/3):125-132.
[12] Kim R, Emi M. Therapeutic potential of antisense Bcl-2 as a chemo sensitizer for patients with gastric carcinoma[J]. J Clin Oncol, 2005,25:40-50.
[13] Molto L, Rayman P. The Bcl-2 transgene protects T cells from renal cell carcinoma-mediated apoptosis[J]. Clin Cancer Res, 2003,45:4060-4068.
[14] Schimmer AD, Hedley DW, Pham NA, et al. BAD induces apoptosis in cells over-expressing Bcl-2 or Bcl-xL without loss of mitochondrial membrane potential[J]. Leuk Lymphoma, 2001, 42(3):429-443.
[15] Hattori T, Ookawa N, Fujita R, et al. Heterodimerizationof bcl-2 and bcl-xl with bax and bad in colorectal Cancer[J]. Aeta Oncol, 2000, 39(4):495-499.
[16] Brown R, Strathdee G. Epigenomics and epigenetic therapy of cancer[J]. Trends Mol Med Suppl, 2002, 8(4):43-48.
[17] Kim YB, Ki SW, Yoshida M, et al. Mechanism of cell cycle arrest caused by histone deacetylase inhibitors in human carcinoma cells[J]. Antibiot(Tokyo), 2000, 53:1191-1200.
猜你喜欢
华人时刊(2022年9期)2022-09-06
华人时刊(2020年15期)2020-12-14
天然产物研究与开发(2018年7期)2018-08-21
上海农业学报(2017年3期)2017-04-10
腹腔镜外科杂志(2016年9期)2016-06-01
西南军医(2016年1期)2016-01-23
中国继续医学教育(2015年4期)2016-01-07
川北医学院学报(2015年5期)2015-12-05
中国当代医药(2015年16期)2015-03-01
中国药理学通报(2014年2期)2014-05-09
