
12种桉树焦枯病病原菌对10种桉树无性系致病性初步分析
2014-07-12 12:27李国清李洁琼刘菲菲李天会陈帅飞
桉树科技 2014年4期
李国清,李洁琼,刘菲菲,李天会,陈帅飞
(国家林业局桉树研究开发中心,广东 湛江 524022)
12种桉树焦枯病病原菌对10种桉树无性系致病性初步分析
李国清,李洁琼,刘菲菲,李天会,陈帅飞*
(国家林业局桉树研究开发中心,广东 湛江 524022)
为了明确在中国发现的桉树叶片焦枯病病原菌Calonectria spp.对桉树的致病力大小,并测定不同桉树无性系对Calonectria病原菌抗病性的强弱,本试验采用在中国发现的12种Calonectria属(Ca. cerciana、Ca. chinensis、Ca. hongkongensis、Ca. microconidialis、Ca. papillata、Ca. parakyotensis、Ca. pauciramosa、Ca. pentaseptata、Ca. pseudoreteaudii、Ca. seminaria、Ca. terrestris、Ca. tetraramosa)共29株致病菌对10个桉树无性系(DH32-22、DH32-29、EC152、EC153、EC155、G1、K31、OC14、U6、W5)进行了室内离体叶片致病性测定。结果显示:12种被测试Calonectria病原菌均能在不同程度上使10个桉树无性系离体叶片产生病斑;不同种Calnectria病原菌对同一桉树无性系的致病性差异显著,不同桉树无性系对同种Calonectria致病菌的抗病能力也存在显著差异。
桉树;焦枯病;丽赤壳属;致病性;抗病育种
桉树(Eucalyptus)具有生长快、轮伐期短、适应性强等特点而成为我国南方速生人工林建设的主要树种[1-3]。近年来,随着国家对林业产业支持力度加大和人们对桉树研究认识不断加深,桉树在我国南方地区发展迅速[3]。至2013年8月,我国桉树面积已达450万hm2[4],相较于2000年(154万hm2)增长将近两倍,桉树人工林为我国经济发展和生态安全做出了巨大贡献[2]。在我国桉树快速发展的同时,部分地区由于桉树经营管理模式不当、种植结构不合理、忽视检验检疫等因素导致桉树人工林受病虫害威胁越来越大[5-7]。
桉树焦枯病是由丽赤壳属Calonectria(Ca.)真菌引起的桉树重大病害之一,可导致桉树叶片焦枯、脱落,致使桉树生长活力下降,生产量降低[8-9]。迄今在我国桉树主要种植区——广西[10]、广东[11]、海南[12]、福建[13-14]等区域均有桉树焦枯病发生,并且已给我国桉树产业造成了巨大的经济损失。截至2014年,已发现丽赤壳属有 90种[13,15-17],其中多个种能引起桉树焦枯病,并能造成巨大危害[8,13]。本文在前期研究的基础上[13,18],利用在我国发现的12种丽赤壳属真菌对 10个桉树无性系进行离体叶片致病性测定并分析不同桉树无性系的抗病差异,以期为今后抗焦枯病桉树无性系选育提供科学依据。
1 材料与方法
1.1 供试材料
供试菌株为我国南方地区发现并鉴定的 12种29株桉树焦枯病病原菌(Calonectria)[11,13,17],其中包括各个种的正模标本菌株(表1)。所有菌株由国家林业局桉树研究开发中心(China Eucalypt Research Centre, Chinese Academy of Forestry)菌种保藏库(CERC)和南非比勒陀利亚—林农生物技术研究所(Forestry and Agricultural Biotechnology Institute, University of Pretoria, South Africa)菌种保藏中心(CMW)提供。
试验所用10个桉树无性系详见表2。
2%MEA培养基:20 g麦芽粉,20 g琼脂,1 L水。

表1 试验所用桉树焦枯病病原菌菌株信息

表2 试验所用10个桉树无性系信息
1.2 试验方法
测定方法和步骤采用李国清等[18]离体叶片致病性测定方法,保持温度30℃、湿度70%。试验在2014年8—9月完成,采用随机区组设计,以无菌MEA培养基菌饼为对照,每株菌接种5片桉树离体叶片。整个试验重复1次以降低试验结果的随机性。
1.3 数据分析
采用Excel 2010、SPSS 20软件进行数据处理和分析。
2 结果与分析
试验结果表明:在接种24 h后,部分桉树无性系叶片上开始产生病斑;4 d后测量病斑大小。通过SPSS单因素方差分析,两次试验结果差异不显著,因此两次试验结果数据合并后进行数据分析。各菌株在不同桉树无性系上病斑长度以及结果差异分析见表3。
除菌株CMW 25283 (Ca. pauciramosa)外,28株菌均在10个桉树无性系离体叶片上产生病斑。其中,菌株 CMW 25283 (Ca. pauciramosa)、CMW 25310 (Ca. pseudoreteaudii)在所有10个无性系上与对照相比均无显著差异(表3)。
同一桉树无性系叶片上,各菌株产生的平均病斑长度差异显著。其中在无性系EC152、EC155、G1、K31、OC14和W5上造成病斑平均长度最大的菌株是CMW 25290 (Ca. cerciana);在无性系 DH32-22、DH32-29和 U6上造成的病斑平均长度最大的菌株是CMW 35179 (Ca. chinesis);在无性系EC153上造成的平均病斑长度最大的是CMW 35180 (Ca. terrestris) (表3,纵向比较)。
同一菌株在不同的桉树无性系上的平均病斑长度也有所不同。如CMW 25311 (Ca. pauciramosa)、CMW 31399 (Ca. hongkongensis)、CMW 31487 (Ca. microconidialis)、CMW 35165 (Ca. papillata)、CMW 35451 (Ca. pentaseptata)、CMW 31449 (Ca. seminaria)、CMW 35366 (Ca. terrestris)、CMW 31474 (Ca. tetraramosa) 和CMW 31476 (Ca. tetraramosa)在无性系 DH32-22较其余无性系上的平均病斑长度最大(表3,横向比较)。
综合比较分析不同种Calonectria致病菌在试验叶片上产生的病斑大小表明,12种Calonectria致病菌在10个桉树无性系离体叶片上均能产生病斑,且均与对照存在显著差异。其中 Ca. cerciana和 Ca. chinensis产生的病斑平均长度最大,其他病原菌产生病斑长度顺序为Ca. parakyotensis > Ca. terrestris > Ca. papillata > Ca. hongkongensis > Ca. seminaria、Ca. microconidialis > Ca. pentaseptata > Ca. tetraramosa > Ca. pseudoreteaudii > Ca. pauciramosa (图1)。
综合分析比较所测试桉树无性系对 12种Calonectria致病菌的抗性差异表明,不同无性系对Calonectria致病菌的抗性存在一定的差异,G1抗性最强,其次是OC14、EC153、EC152、W5、U6、EC155、K31和DH32-22,DH32-29抗性最弱(图2)。

W 5 .08 6.21±0a 1/a2 .31 3.59±0d 1-f 1/c2 .13 6.31±0a 1/a2 .82 4.22±0c 1-e 1/a2b2 .00 0.50±0k 1/a2 .09 1.81±0h 1-j 1/a2b2 .68 3.33±0e 1-g 1/a2-c 2 .23 1.92±0h 1-j 1/d2 .27 4.52±0b 1-d 1/a2b2 .26 2.07±0h 1i1/b2c2 .37 1.70±0h 1-k 1/b2 .12 1.16±0i 1-k 1/d2e2 .08 1.16±0i 1-k 1/e2 .38 5.21±0a 1-c 1/a2b2 .38 4.18±0c 1-e 1/b2c2 cmU 6 4.62±0.59 b 1-d 1/c2 5.19±0.19 a 1-c 1/a2 6.11±0.10 a 1/a2b2 4.28±0.78 c 1-e 1/a2b2 0.50±0.00 m 1/a2 1.53±0.05 i 1-m 1/c2 3.71±0.64 d 1-g 1/a2-c 2 2.72±0.14 g 1-j 1/c2d2 4.42±0.48 b 1-d 1/a2b2 2.44±0.31 g 1-j 1/a2b2 2.18±0.29 h 1-k 1/a2b2 1.67±0.19 h 1-m 1/b2c2 1.34±0.06 j 1-m 1/d2e2 4.81±0.42 b 1-d 1/a2-c 2 4.47±0.37 b 1-d 1/a2-c 2 OC145.29±0.29 a 1/b2c2 5.01±0.12 a 1b1/a2 5.06±0.27 a 1b1/c2 4.84±0.18 a 1b1/a2b2 0.50±0.00 i 1/a2 1.65±0.10 f 1-h 1/b2c2 2.34±0.36 f 1/a2-c 2 3.39±0.42 d 1e1/b2c2 4.39±0.23 a 1-d 1/a2b2 2.35±0.33 f 1/a2-c 2 1.86±0.19 f 1g1/b2 0.78±0.08 g 1-i 1/e2f2 1.76±0.10 f 1g1/a2-c 2 3.63±0.30 c 1d1/c2 3.69±0.33 c 1d1/c2 K 316.58±0.07 a 1b1/a2 5.67±0.17 a 1-c 1/a2 6.74±0.11 a 1/a2 3.79±0.91 d 1-g 1/b2 0.50±0.00 m 1/a2 1.81±0.04 i 1-m 1/a2b2 3.80±0.61 d 1-g 1/a2b2 3.93±0.51 d 1-f 1/b2 4.39±0.41 c 1-e 1/a2b2 2.13±0.30 h 1-l 1/b2c2 2.56±0.39 f 1-k 1/a2b2 2.25±0.22 h 1-k 1/a2 1.42±0.08 k 1-m 1/c2-e 2 5.93±0.29 a 1b1/a2 5.29±0.22 b 1c1/a2况G 1 a 1/c2 a 1/c2 i 1/a2 i 1/f2情系5.01±0.31 4.31±0.41 a 1-c 1/b2 4.95±0.34 4.34±0.33 a 1-c 1/a2b2 0.50±0.00 1.68±0.09 f 1-i 1/a2-c 2 2.18±0.41 f 1-h 1/b2c2 2.41±0.17 e 1-g 1/c2d2 4.22±0.29 a 1-c 1/a2b2 2.53±0.28 d 1-f 1/a2b2性2.55±0.19 d 1-f 1/a2b2 0.62±0.07 1.54±0.20 f 1-i 1/b2-e 2 3.69±0.48 b 1-d 1/c2 3.65±0.40 b 1-d 1/c2度长无斑树病桉上EC155 a 1/a2 h 1/a2 c 1/b2 d 1/a2片6.18±0.23 5.50±0.15 a 1-c 1/a2 5.43±0.35 a 1-c 1/c2 5.64±0.13 a 1b1/a2 0.50±0.00 1.86±0.12 d 1-g 1/a2b2 2.91±0.52 d 1/a2-c 2 4.30±0.59 5.15±0.14 a 1-c 1/a2 2.89±0.25 d 1/a2b2 2.87±0.15 1.48±0.17 e 1-h 1/c2d2 1.86±0.17 d 1-g 1/a2b2 4.81±0.28 b 1c1/a2-c 2 4.23±0.49 c 1/b2c2叶系性无树EC153 j 1/a2桉5.30±0.22 a 1b1/b2c2 5.12±0.13 a 1b1/a2 5.42±0.25 a 1b1/c2 4.52±0.27 b 1-d 1/a2b2 0.50±0.00 1.85±0.04 f 1-h 1/a2b2 2.00±0.36 f 1-h 1/c2 1.83±0.16 f 1-h 1/d2 3.53±0.55 d 1e1/b2 1.50±0.17 g 1-j 1/c2 2.81±0.34 e 1f1/a2 0.98±0.06 g 1-j 1/e2f2 1.77±0.11 f 1-i 1/a2-c 2 4.00±0.42 c 1d1/c2 4.02±0.37 c 1d1/c2个10在菌原EC152 a 1/a2b2 a 1b1/a2 a 1/b2c2 k 1/a2 e 1-h 1/d2 e 1/a2b2病5.84±0.20 5.26±0.11 5.57±0.13 5.14±0.16 a 1-c 1/a2b2 0.50±0.00 1.86±0.05 e 1-j 1/a2b2 2.13±0.21 e 1-h 1/b2c2 2.11±0.38 4.43±0.27 b 1-d 1/a2b2 2.30±0.30 e 1f1/a2-c 2 2.62±0.12 1.14±0.10 g 1-k 1/d2e2 1.72±0.16 e 1-j 1/a2-d 2 4.14±0.34 c 1d1/b2c2 4.14±0.28 c 1d1/b2c2 Calonectria 6.15±0.09 DH32-29 a 1b1/a2 5.44±0.16 a 1-c 1/a2 6.39±0.08 a 1/a2 4.55±0.59 c 1d1/a2b2 0.50±0.00 l 1/a2 1.80±0.04 i 1j1/a2b2 3.63±0.59 5.48±0.22 a 1-c 1/a2 4.96±0.21 c 1/a2 3.20±0.12 e 1-g 1/a2 d 1-f 1/a2-c 2 2.56±0.33 g 1-j 1/a2b2 2.07±0.20 h 1-j 1/a2b2 1.60±0.18 j 1k1/a2-d 2 5.32±0.38 b 1c1/a2b2 5.35±0.22 b 1c1/a2种312表DH32-22 5.86±0.20 a 1-c 1/a2b2 5.22±0.15 b 1-d 1/a2 6.29±0.10 a 1/a2 3.67±0.65 f 1-h 1/b2 0.000.5±k 1/a2 1.91±0.05 j 1/a2 3.98±0.73 e 1-g 1/a2 5.47±0.21 a 1-d 1/a2 3.26±0.80 g 1-i 1/b2 2.46±0.36 i 1j1/a2b2 2.87±0.32 h 1-j 1/a2 1.89±0.16 j 1/a2-c 2 1.99±0.09 j 1/a2 5.29±0.38 b 1-d 1/a2b2 5.13±0.23 b 1-d 1/a2b2株529053095179536752835311139914385175147114871492516579727976菌W2W2W3W3W2W2W3W3W3W3W3W3W3W3W3CMCMCMCMCMCMCMCMCMCMCMCMCMCMCMgensis种菌Ca.cerciana Ca.cerciana Ca.chinensisCa.chinensisciramosaCa.pauciramosaCa.pauCa.hongkongkongensisCa.hongkongensisCa.honicroconidialisillataillataillataCa.microconidialisCa.microconidialisCa.mCa.papCa.papCa.pap

5 W 5.55±0.27 a 1b1/a2b2 1.23±0.04 i 1-k 1/c2 1.62±0.10 h 1-k 1/c2d2 2.24±0.38 g 1-i 1/d2 6.21±0.35 h 1i1/a2-c 2 0.71±0.03 j 1k1/c2d2 2.67±0.11 f 1-h 1/a2-c 2 2.04±0.06 h 1i1/b2 2.30±0.07 g 1-i 1/a2 5.30±0.25 a 1-c 1/a2-d 2 5.21±0.30 a 1-c 1/b2 1.80±0.25 h 1-j 1/c2d2 1.53±0.10 h 1-k 1/b2 1.40±0.12 i 1-k 1/b2 0.50±0.00 k 1/b2在株菌一U 6 3.07±0.44 e 1-h 1/c2 1.41±0.11 j 1-m 1/b2c2 2.17±0.25 h 1-k 1/b2c2 2.63±0.52 g 1-j 1/c2d2 2.40±0.38 g 1-k 1/a2b2 0.99±0.10 k 1-m 1/a2-c 2 2.89±0.13 f 1-i 1/a2b2 2.37±0.14 g 1-k 1/a2 2.97±0.11 e 1-h 1/a2b2 5.75±0.28 a 1b1/a2b2 4.14±0.35 c 1-f 1/c2 1.98±0.22 h 1-l 1/b2-d 2 1.79±0.20 h 1-m 1/a2b2 1.96±0.17 h 1-l 1/a2b2 0.61±0.04 l 1m1/a2b2同示f 2表OC144.05±0.83 b 1-d 1/a2-c 2 1.65±0.25 f 1-h 1/a2-c 2 0.95±0.10 g 1-i 1/e2 1.49±0.28 f 1-i 1/d2 1.01±0.13 g 1-i 1/d2 0.63±0.13 h 1i1/d2 2.31±0.10 f 1/b2-d 2 0.75±0.11 g 1-i 1/c2 2.56±0.27 e 1f1/a2b2 4.56±0.24 a 1-c 1/d2 4.68±0.15 a 1-c 1/b2c2 1.74±0.15 f 1-h 1/c2d2 0.94±0.12 g 1-i 1/d2 2.21±0.27 f 1/a2 0.52±0.02 i 1/a2b2 a 2,b2,c2…);向K 314.59±0.79 c 1d1/a2-c 2 1.38±0.08 k 1-m 1/b2c2 1.77±0.23 i 1-m 1/c2 3.34±0.33 d 1-h 1/b2c2 2.77±0.35 f 1-k 1/a2 0.77±0.04 l 1m1/b2-d 2 3.14±0.19 e 1-i 1/a2 2.45±0.07 g 1-k 1/a2 2.94±0.13 f 1-j 1/a2-c 2 5.67±0.41 a 1-c 1/a2-c 2 6.51±0.20 a 1b1/a2 1.60±0.15 j 1-m 1/d2 1.67±0.12 j 1-m 1/b2 2.09±0.24 h 1-l 1/a2 0.63±0.06 m 1/a2b2 (纵性著显异差的上系G 1 f 1-i 1/d2 i 1/d2 i 1/c2 i 1/a2平性3.50±0.82 c 1-e 1/b2c2 1.62±0.23 f 1-i 1/a2-c 2 1.16±0.11 g 1-i 1/d2e2 1.71±0.36 1.41±0.24 f 1-i 1/c2d2 0.57±0.05 2.25±0.16 f 1-h 1/b2-d 2 0.58±0.06 2.34±0.21 e 1-h 1/b2-d 2 4.78±0.29 a 1b1/c2d2 4.68±0.18 a 1-c 1/b2c2 1.70±0.40 f 1-i 1/c2d2 1.06±0.18 h 1i1/c2d2 2.19±0.23 f 1-h 1/a2 0.71±0.15水无树0.05桉EC155 4.73±0.67 b 1c1/a2-c 2 2.06±0.28 d 1-g 1/a2 2.09±0.31 d 1-f 1/b2c2 2.24±0.26 d 1e1/d2 1.83±0.19 d 1-g 1/b2-d 2 0.92±0.19 e 1-h 1/a2-d 2 2.31±0.19 d 1e1/b2-d 2 0.78±0.10 e 1-h 1/c2 2.35±0.22 d 1e1/b2-d 2 4.99±0.57 a 1-c 1/b2-d 2 5.45±0.40 a 1-c 1/b2 2.94±0.44 d 1/a2 1.40±0.13 e 1-h 1/b2c2 2.30±0.21 d 1e1/a2 0.53±0.03 h 1/a2b2在度长斑病的EC153 3.50±0.75 d 1e1/b2c2 1.61±0.27 g 1-j 1/a2-c 2 1.00±0.12 g 1-j 1/e2 1.82±0.42 f 1-h 1/d2 1.30±0.21 g 1-j 1/c2d2 1.08±0.17 g 1-j 1/a2b2 2.13±0.16 f 1-h 1/c2d2 0.65±0.08 g 1-j 1/c2 2.05±0.15 f 1-h 1/c2d2 5.98±0.13 a 1/a2 4.99±0.22 a 1-c 1/b2 2.52±0.20 e 1-g 1/a2-d 2 1.03±0.16 g 1-j 1/c2d2 2.09±0.21 f 1-h 1/a2 0.60±0.06 j 1/a2b2生产)。上向片叶(横系性EC152 3.81±0.83 d 1/b2c2 1.88±0.20 e 1-j 1/a2b2 0.99±0.09 h 1-k 1/e2 1.92±0.46 e 1-j 1/d2 2.03±0.30 e 1-i 1/a2-c 2 1.20±0.15 f 1-k 1/a2 2.25±0.18 e 1-g 1/b2-d 2 0.80±0.10 j 1k1/c2 2.25±0.25 e 1/c2d2 5.89±0.15 a 1/a2b2 5.34±0.18 a 1b1/b2 2.77±0.39 e 1/a2b2 0.94±0.12 i 1-k 1/d2 2.09±0.31 e 1-h 1/a2 0.60±0.03 k 1/a2b2性著无显树异桉差一的DH32-29 5.97±0.11 a 1b1/a2 1.65±0.13 j 1/a2-c 2 2.80±0.24 f 1-i 1/a2 3.84±0.23 d 1e1/a2b2 2.80±0.36 f 1-i 1/a2 0.70±0.00 k 1l1/c2d2 1.99±0.37 h 1-j 1/d2 2.62±0.05 f 1-j 1/a2 2.85±0.05 f 1-h 1/c2d2 6.14±0.19 a 1b1/a2 6.29±0.16 a 1b1/a2 2.63±0.26 f 1-j 1/a2-c 2 1.81±0.13 h 1-j 1/a2b2 2.33±0.14 g 1-j 1/a2 0.63±0.07同上l 1/a2b2在平株水同0.05菌不在DH32-22 4.86±0.35 c 1-e 1/a2-c 2 1.86±0.13示度j 1/a2b2 2.61±0.21 i 1j1/a2b2 4.57±0.13 d 1-f 1/a2 2.63±0.32 i 1j1/a2b2 0.88±0.07 k 1/a2-d 2 2.58±0.30 i 1j1/a2-d 2 2.64±0.15 i 1j1/a2 3.35±0.12 g 1-i 1/d2 5.90±0.17 a 1b1/a2b2 5.10±0.39 b 1-d 1/b2 3.39±0.28 g 1-i 1/a2 2.20±0.09 j 1/a2 2.61±0.16 i 1j1/a2 0.59±0.04 k 1/a2b2 a 1,b1,c1…病m1表长斑的生产株25292 25310上母片菌W35169CMW35377CMW35436CMW35451CMMWMWW31446CMW31449CMW31450CMW35180CMW35364CMW35366CMW31474CMW31476CM字叶的系方性下无字树数桉中同表种表不上菌inariainariainaria:续Ca.parakyotensisCa.pentaseptata Ca.pentaseptata Ca.pentaseptata Ca.pseudoreteaudiiCCa.pseudoreteaudiiCCa.semCa.semCa.semCa.terrestris Ca.terrestris Ca.terrestris Ca.tetraramosaCa.tetraramosa注CK
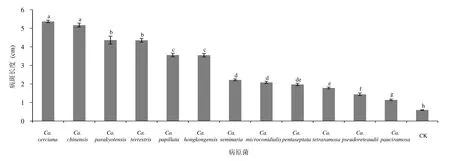
图1 12种Calonectria致病菌对桉树无性系的致病力差异比较

图2 10个桉树无性系对Calonectria致病菌抗病性差异比较
3 结论与讨论
本研究对我国南方地区(主要为广东省)发现的12种Calonectria致病菌在10个桉树无性系上进行致病力测试,结果表明:不同Calonectria致病菌对同一桉树无性系的致病性存在显著差异,不同桉树无性系对同一Calonectria致病菌的抗病性也存在差异。
选育抗病无性系是防控丽赤壳属真菌危害桉树人工林的重要措施之一。陈全助[19]对福建省永安市某一地区的11个桉树种(系)上桉树焦枯病的发病情况进行调查,结果表明这一地区不同桉树种(系)对焦枯病的抗病性存在明显差异。Rodas等[20]对哥伦比亚地区的Ca. spathulata (无性型:Cy. spathulatum)在42个巨桉无性系上的致病性测试结果证明不同无性系对Ca. spathulata的抗性存在显著差异。陈帅飞等[13]对在福建省发现的4种Calonectria致病菌在2个桉树无性系上进行了致病力测定试验,发现4种Calonectria对桉树无性系均有一定的致病性,且在不同无性系上存在差异。李国清等[18]通过对在我国广东、福建发现的5种Calonectria致病菌在4个桉树无性系上的致病性测定,发现在所测试桉树无性系上,Ca. cerciana的致病性最强,G1的抗病性较其他无性系强,这与本文研究结果一致。因此,通过针对不同种焦枯病病原菌在不同桉树无性系上的致病力测定,可阐明焦枯病病原菌对特定桉树无性系的致病力强弱,并阐明桉树不同无性系对特定焦枯病病原菌的抗性差异,从而为桉树焦枯病的防控提供依据。
由丽赤壳属真菌引起的桉树焦枯病是危害我国桉树人工林健康可持续发展的重大病害之一,由于焦枯病病原存在物种多样性,不同的省份和区域病原可能存在差异,因此该病的防控较困难。目前,桉树焦枯病对我国桉树大面积危害已有多年,但针对不同桉树焦枯病病原菌进行桉树无性系致病力测定尚处于起步阶段,因此十分有必要对Calonectria病原菌在我国华南地区桉树人工林的分布进行系统的调查,对其物种多样性和种群多样性进行系统研究,并进一步选育出特定区域抗特定焦枯病病原菌的无性系,同时要严格防控病原菌通过桉树苗木、育苗基质等在桉树种植不同区域之间的传播[21],以防控不同桉树焦枯病病原菌大面积蔓延和危害。
[1] 谢耀坚.世纪初的桉树研究[M].北京:中国林业出版社, 2006.
[2] 祁述雄.中国桉树(第 2版)[M].北京:中国林业出版社, 2002.
[3] 杨民胜,谢耀坚,刘杰锋.中国桉树研究三十年(1981—2010)[M].北京:中国林业出版社,2011.
[4] 陈少雄,陈小菲.我国桉树经营的技术问题与思考[J].桉树科技,2013,30(3):52-59.
[5] 谢耀坚.中国桉树人工林可持续经营战略初探[J].世界林业研究,2003,16(5):59-64.
[6] Zhou X D,Wingfield M J.Eucalypt diseases and their management in China[J].Australasian Plant Pathology, 2011,40(4):339-345.
[7] 陈帅飞.中国桉树真菌病原汇录:2006—2013[J].桉树科技,2014,31(1):37-65.
[8] Old K M,Wingfield M J,Yuan Z Q.A Manual of Diseases of Eucalypts in South-East Asia[M].Indonesia:Centre for International Forestry Research,2003.
[9] Lombard L,Crous P W,Wingfield B D,et al.Species concepts in Calonectria (Cylindrocladium)[J].Studies in Mycology,2010,66(1):1-13.
[10] 黄翠流,陈唯王,罗基同,等.广西速生桉叶部真菌性病害的病原鉴定[J].中国森林病虫,2012,31(3):1-6.
[11] Lombard L,Zhou X D,Crous P W,et al.Calonectria species associated with cutting rot of Eucalyptus[J].Persoonia, 2010(24):1-11.
[12] 唐真正,周诗萍.海南省儋州市桉树主要病虫害发生与防控对策[J].热带林业,2007,35(3):42-44.
[13] Chen S F,Lombard L,Roux J,et al.Novel species of Calonectria associated w ith Eucalyptus leaf blight in Southeast China[J].Persoonia,2011(26):1-12.
[14] 朱建华,郭文硕,陈红梅,等.桉树焦枯病对桉树生长量的损失估计研究[J].中国森林病虫,2011,30(5):6-10.
[15] Xu J J,Qin S Y,Hao Y Y,et al.A new species of Calonectria causing leaf disease of water lily in China[J].Mycotaxon,2012,122(1):177-185.
[16] Lombard L,Crous P W,Wingfield B D,et al.Phylogeny and systematics of the genus Calonectria[J].Studies in Mycology, 2010,66(1):31-69.
[17] Lombard L,Chen S F,Mou X T,et al.New species, hyperdiversity and potential importance of Calonectria spp. From Eucalyptus in South China[J].Studies in Mycology, 2014.DOI:10.1016/j.simyco.2014.11.003.
[18] 李国清,陈帅飞,吴志华,等.中国桉树焦枯病病原菌物种多样性及致病力初步分析[J].热带作物学报,2014,35(6): 1183-1191.
[19] 陈全助.桉树种(系)对焦枯病抗性生理的研究[D].福州:福建农林大学,2007.
[20] Rodas C A,Lombard L,Gryzenhout M.Cylindrocladium blight of Eucalyptus grandis in Colombia[J].Australasian Plant Pathology,2005,34(2):143-149.
[21] 陈帅飞.林木病原菌的传播途径概述[J].桉树科技,2014, 31(2):50-56.
Prelim inary Analyses on Pathogenicity of Twelve Calonectria spp. on Ten Eucalyptus Clones in China
LI Guo-qing, LI Jie-qiong, LIU Fei-fei, LI Tian-hui, CHEN Shuai-fei
(China Eucalypt Research Centre, Zhanjiang 524022, Guangdong, China)
Calonectria leaf blight, caused by species of the genus Calonectria, is w idely distributed in Eucalyptus plantations in tropical and subtropical regions of the world. The aims of this study were to clarify the pathogenicity of different species of Calonectria on a range of Eucalyptus hybrid clones and understand the resistance of different clones against Calonectria species in China. Twenty-nine isolates representing 12 species of Calonectria (Ca. cerciana, Ca. chinensis, Ca. hongkongensis, Ca. microconidialis, Ca. papillata, Ca. parakyotensis, Ca. pauciramosa, Ca. pentaseptata, Ca. pseudoreteaudii, Ca. seminaria, Ca. terrestris, Ca. tetraramosa) were tested on 10 Eucalyptus clones (DH32-22, DH32-29, EC152, EC153, EC155, G1, K31, OC14, U6, W5). Leaf inoculation showed that all 12 tested Calonectria species produced lesions on all 10 Eucalyptus clones, to varying degrees. The pathogenicity of different Calonectria species to the same Eucalyptus clone differed significantly, and the resistance of different Eucalyptus clones to the same species of Calonectria also differed significantly.
Eucalyptus; leaf blight; Calonectria; pathogenicity; resistance breeding
S763.15
A
2014-11-10
广东省林业科技创新专项“桉树重大病虫害控制技术研究与示范”(2010KJCX015-03);广东省自然科学基金“广东桉树焦枯病及无性系选择研究”(10145624536-400000);国家国际科技合作专项项目“重要林业病害致病力及多样性研究”(2012DFG31830)
李国清(1988— ),硕士,助理工程师,主要从事森林病害研究
*陈帅飞为通讯作者,E-mail:cerccsf@126.com
猜你喜欢
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02
环球时报(2019-01-03)2019-01-03
中成药(2017年12期)2018-01-19
上海农业学报(2017年4期)2017-04-10
湖南畜牧兽医(2016年3期)2016-06-05
现代食品(2016年24期)2016-04-28
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年4期)2016-03-16
中华老年口腔医学杂志(2016年5期)2016-03-01
川北医学院学报(2015年5期)2015-12-05
