
旱涝交替胁迫对水稻抗折力的影响及其机理分析
2014-07-07 15:38杨静晗郭相平杨骕王振昌
山东农业大学学报(自然科学版) 2014年3期
杨静晗,郭相平,杨骕,王振昌
河海大学南方地区高效灌排与农业水土环境教育部重点实验室,江苏南京210098
旱涝交替胁迫对水稻抗折力的影响及其机理分析
杨静晗,郭相平,杨骕,王振昌*
河海大学南方地区高效灌排与农业水土环境教育部重点实验室,江苏南京210098
采用盆栽试验研究了水稻分蘖期和拔节期旱涝交替胁迫后的对水稻细观结构、微观结构、茎秆充数度及其抗折力的影响规律。研究结果表明:旱涝胁迫后茎粗、叶鞘厚度、与抗折力无正相关。大维管束的数目与抗折力呈正相关,拔节期轻旱使茎秆干物质累积量和所占比例提高,对水稻抗折力影响较大;重度旱胁迫无法提高水稻的抗折力。分蘖期的干旱胁迫虽然大维管束的面积有所增大,但总体抗折力无增加。因此蓄水控灌模式下对水稻拔节期进行旱涝胁迫程度适宜控制,可获得较传统浅水勤灌更高的抗倒伏能力。
水稻;旱涝交替胁迫;茎秆物理性状;抗折力;叠加效应
倒伏是影响水稻产生的重要因素,能使水稻减产在10%甚至更高[1-4]。水稻倒伏有根倒伏和茎倒伏,其中茎倒伏在江苏较为普遍。一般认为水稻茎秆的抗倒伏能力,与茎粗、茎壁厚、大小维管束数目以及截面积尺寸有关[5-7];水稻茎秆节间充实度、茎壁厚度、茎秆横截面等指标的增加,可改善水稻茎秆抗倒伏性能[8],而上述指标的变化,除受品种特性、种植密度、气象条件等因素影响外,还与灌排管理密切相关。大部分研究成果表明,单纯的干旱胁迫能够增加基部茎粗、茎秆强度和壁厚[9-10],从而提高水稻茎秆抗倒伏能力,而淹水胁迫的作用则相反[11-13]。
蓄水控灌是最近几年发展起来的一种节水灌排模式。该模式下,较低的灌水下限会导致一定程度的干旱胁迫发生,而为提高雨水利用效率所采用的高蓄水上限,在暴雨发生频率较高的南方地区,水稻会受到干旱和淹水两种不同性质的交替胁迫。在具有相反作用的两种胁迫交替模式下,水稻茎秆的抗倒伏指标会如何变化?目前该方面的研究不多。郭相平等人的小区试验结果表明,水稻茎秆倒伏能力增减与胁迫程度有关,并从茎秆的力学指标方面对影响机理进行了初步分析[14]。旱涝交替胁迫不仅影响水稻干物质累积量,而且影响干物质沿茎秆的垂向分布,从而改变倒伏力矩和茎秆强度;旱涝交替模式下,基部叶鞘的厚度发生变化,并对茎秆的抗倒伏产生影响。上述指标如何影响水稻的抗倒伏指标,目前尚未可知。本文利用盆栽试验资料,分析了蓄水控灌模式下,旱涝交替胁迫对水稻基部茎粗、壁厚、叶鞘厚度、充实度、维管束等参数对水稻抗倒伏能力的影响,分析该模式下水稻茎秆抗倒伏指标的变化规律和机理,为制定水稻合理灌排制度提供理论依据。
1 试验材料与方法
1.1 试验区简介
试验于2013年4~10月在河海大学节水园区进行。试区位于长江下游,属于亚热带湿润气候,多年平均降雨量为1022 mm,夏季暴雨多发。
试验所采用干土取自临近稻田耕作层的黏壤土。土壤田间持水量为30.9%(重量含水量),土壤类型为黏壤土,中等肥力。其主要农化性状如表1

表1 试验地主要农化性状(单位:mg/kg)Table 1 Testing main agrochemical properties
1.2 试验材料与方法
供试水稻品种为当地高产品种“南粳44”,采用桶栽试验。桶底部直径18 cm,上部直径24 cm,高度25.5 cm。桶置于遮雨棚内。
所用土壤经过晒干、打碎、过筛后,每桶装干土7.5㎏。每桶的施肥量分别为P2O50.10 g、N 0.15 g、k2O 0.10 g。
水稻4月11日开始育种,三叶一心移栽,每盆移栽5株,分蘖初期视苗情长势,每盆留长势相近的3株。并在分蘖期和拔节期进行控水处理,用雨棚防雨。分蘖期于7月22日干旱胁迫处理,拔节期于8月15日干旱胁迫,胁迫5 d结束并进行旱涝快速转换处理。
各处理水稻通过腾发和排水进行干旱。在达到胁迫程度和时间后,立即植入塑料方盒内模拟涝胁迫。淹水胁迫结束后,再次通过排水和腾发进行干旱胁迫,达到胁迫程度和时间后,恢复至对照处理。除水分胁迫外,其他农技措施相同。为避免水温变幅过大,淹水胁迫前预先放入附近坑塘水(与灌溉用水同),保持3 d以上。根据倒伏发生的时间,各处理植株茎秆在黄熟期(10月11日)齐地面收割后测试。
1.3 试验设计
试验采用2因素2水平,即水分胁迫分蘖期和拔节期;胁迫历时:5 d,胁迫程度:轻旱、重旱。共设置5种处理,在干旱胁迫期间采用称重法控制土壤水分,下限控制如表2所示。除水分外,其它农技措施相同。

表2 水稻盆栽试验设计方案Table 2 The rice pot experiment design
1.4 测定指标和方法
根据相关资料,水稻茎秆基部0~10 cm茎段(通常为基部1~3伸长节)和抗倒伏能力有极显著的相关关系[7],水稻的倒伏经常是发生在基部的第二节间,故本试验选用第二伸长节间作为研究对象。由于倒伏主要发生在成熟期,故测试时间为10月11日。
(1)水稻生长指标。在测量株高后,取出植株,用游标卡尺测量基部第二伸长节间中部最大的直径和最小的直径,取平均数,即:茎粗d=(最长直径+最短直径)/2。剪开第二伸长节间,用游标卡尺测量茎壁厚度,每测一次将样品转动90°,连续4次读数,取4次的均值。
(2)干重测定:置于恒温箱内105℃杀青30 min,80℃烘干至恒重,放置感量为0.01 g的电子天平上称重。
(3)抗折力测定:茎秆抗折力是水稻茎秆倒伏的主要形式,故本试验中取基部以上10 cm茎秆进行测试。抗折力参考了马均和濑古秀生[15-16]的方法。茎秆水平放置,两侧被固定为简支梁,中间10 cm测试部分悬空。采用电子茎秆强度试验机(型号为CMT6104),其特制的压力应变传感器置于茎秆中心位置,自上而下施加垂向压力(相当于文献[5]中的砝码),模拟茎秆折力。传感器以0.1 mm/s的速度向下位移。随着传感器不断下移,茎秆压力和挠度不断增加,当茎秆发生屈服破坏时压力达到最大值,此时的压力即为茎秆的最大抗折力。
(4)茎秆微观结构观察及测量:将做好的茎秆切片放在奥林巴斯显微镜CX41下观察。在20倍镜头下拍照,并记录一个镜头中大小维管束的个数。在40倍镜头下测量大小维管束的面积。
2 结果与分析
2.1 旱涝交替胁迫对水稻细观结构及其抗折力的影响
水稻茎秆的茎鞘是叶片基部较宽或扩大成片状,形成圆筒体,将茎秆的一部分或者是全部紧密包围,担负着支撑、保护幼芽及节间生长。旱涝交替胁迫处理中,基部茎秆的茎鞘厚度均显著低于对照,且分蘖期最小,并显著低于CK和拔节期处理。茎鞘在茎秆抗折力中具有重要影响[17],本试验结果表明,在去除茎鞘后,茎秆抗折力下降36%~74%,其中旱涝胁迫处理的为36%~68%,尤其是分蘖重度胁迫仅为36%,表明旱涝交替胁迫下,茎鞘对抗折力的影响较对照有所降低。这与旱涝交替胁迫模式下茎鞘的厚度减小有关(见表3)。本试验还表明,茎秆的抗折力与茎鞘厚度并无明显的正相关关系,与其他学者的观点不符。这可能旱涝交替胁迫改变了茎鞘成分或结构有关,有待于进一步研究。
正常栽培条件下,茎粗与茎秆的抗折力正相关[7]。但在旱涝交替胁迫下,本试验的结果似乎并不支持上述观点,这可能与茎秆的成分[18]、微观结构和充实度有关(分析见后)。一般认为,干旱胁迫能够抑制茎秆的延伸生长,增加茎秆的厚度[14];而在淹水胁迫下,茎秆会加速生长,导致茎秆直径减小[19],与干旱胁迫具有相反的作用。表3结果表明,除了拔节期重度干旱处理外,在旱涝交替胁迫下,其他处理茎秆直径(茎粗)与对照差异不显著。这表明可以利用干旱胁迫拮抗淹水胁迫对茎秆发育和抗折力下降的不利影响。拔节期是营养生长和生殖生长并行时期,该阶段重度干旱会降低叶片光合作用和干物质累积,进而抑制茎秆生长,导致茎粗降低,而淹水胁迫则放大了这种趋势[20]。因此,拔节期重度干旱与淹水胁迫下表现为联合效应而非拮抗效应,最终导致茎粗的显著降低。可能是拔节期随着干旱胁迫程度的加剧,植物通过降低生长速率,抑制株高生长[21],即使是后期的淹水补偿效应仍然无法消除重度干旱后对作物的影响,以至于水稻吸收和运输营养物质的能力降低、茎秆变细。
2.2 旱涝交替胁迫对水稻微观结构及其抗折力的影响
试验发现拔节期维管束面积显著下降,其中重度干旱胁迫处理下降最为显著,而分蘖期大维管束的面积均显著增加(见表4)。无论分蘖期还是拔节期的维管束面积的变化都与胁迫程度成负相关。稻株内维管束是一个相互连通的系统组织,地上器官(叶、叶鞘、茎)中都有大小两种维管束。大小维管束主要根据维管束横截面积的大小、功能、分布及其延伸的长度来划分的,大维管束之间分布有数条小维管束,从显微镜下观察,其大维管束的横截面积大,有许多与后生导管相连的木质部薄壁细胞,小维管束的横截面积小,其中后生韧皮部和后生木质部间的伴胞和韧皮薄壁细胞能够从自由空间吸收光合产物到细胞内[23-24]。重度干旱胁迫后抑制了叶片的光合作用,抑制了水分、营养物质的运输,导致营养供应不足、维管束面积变小;轻度旱胁迫后的水稻具有较强的补偿生长能力,促进茎秆吸收营养物质、维管束分化加快、数目或面积增加。
维管束组织由初生木质部和初生韧皮部所组,其中在木质部中含大量木纤维;是水稻植株由根部向穗部输送水分、矿质和有机养分的主要通道,因此维管束为增强性材料,相当于混凝土中的钢筋[23]。相关学者[15]认为茎秆中维管束数目、面积与水稻抗折力成正相关。本试验与其他学者的研究基本一致。水稻茎秆的抗折力与大维管束的数目呈正相关,但小维管束的数目及面积并无显著性差异(见表4)。从显微镜下观察水稻茎秆切片发现,轻旱胁迫处理能够增加大维管束面积(如TLD),或者增加大维管束个数(如SLD),维管束之间排列相对紧密,重度的干旱抑制大维管束细胞的生长,排列相对松散(如图1)。其原因可能是轻度旱胁迫加快营养物质运输,促进维管束的发育,木质部中所含有的木薄壁细胞和木纤维的含量增加,进而导致茎秆机械强度提高。因此适度的轻旱胁迫有利于水稻茎秆的生长发育、增加大维管束面积或个数、维管束更加坚硬而富有弹性,提高茎秆的机械强度。
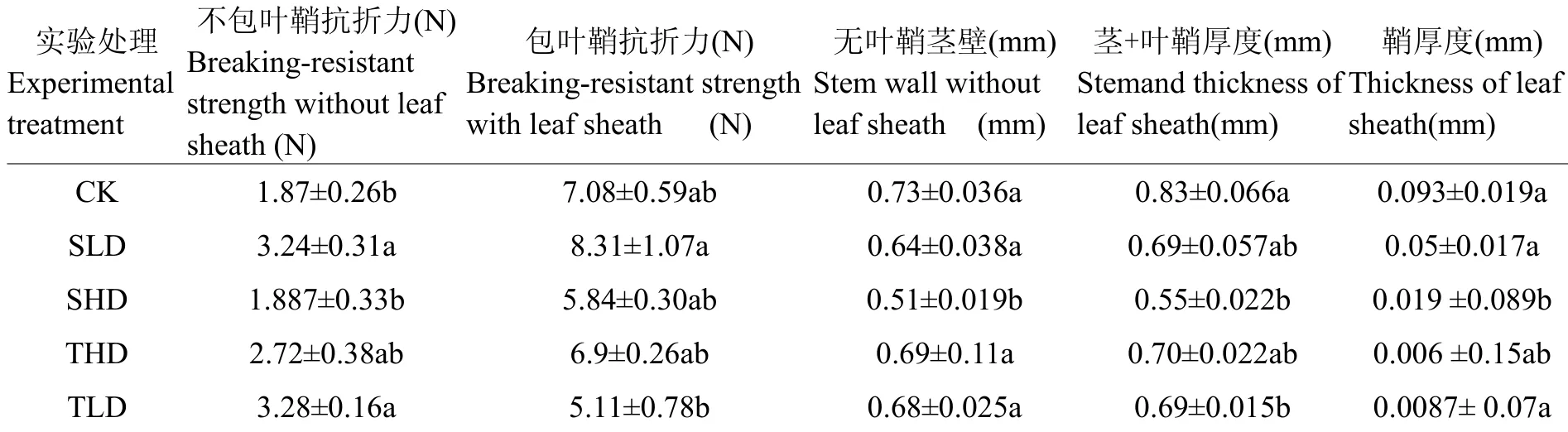
表3 水稻基部抗折力和壁厚之间的关系Table 3 Rice base bending force and the relationship between the wall thickness

表4 不同处理的水稻茎秆微观结构参数Table 4 Haulm microstructure of different processing parameters
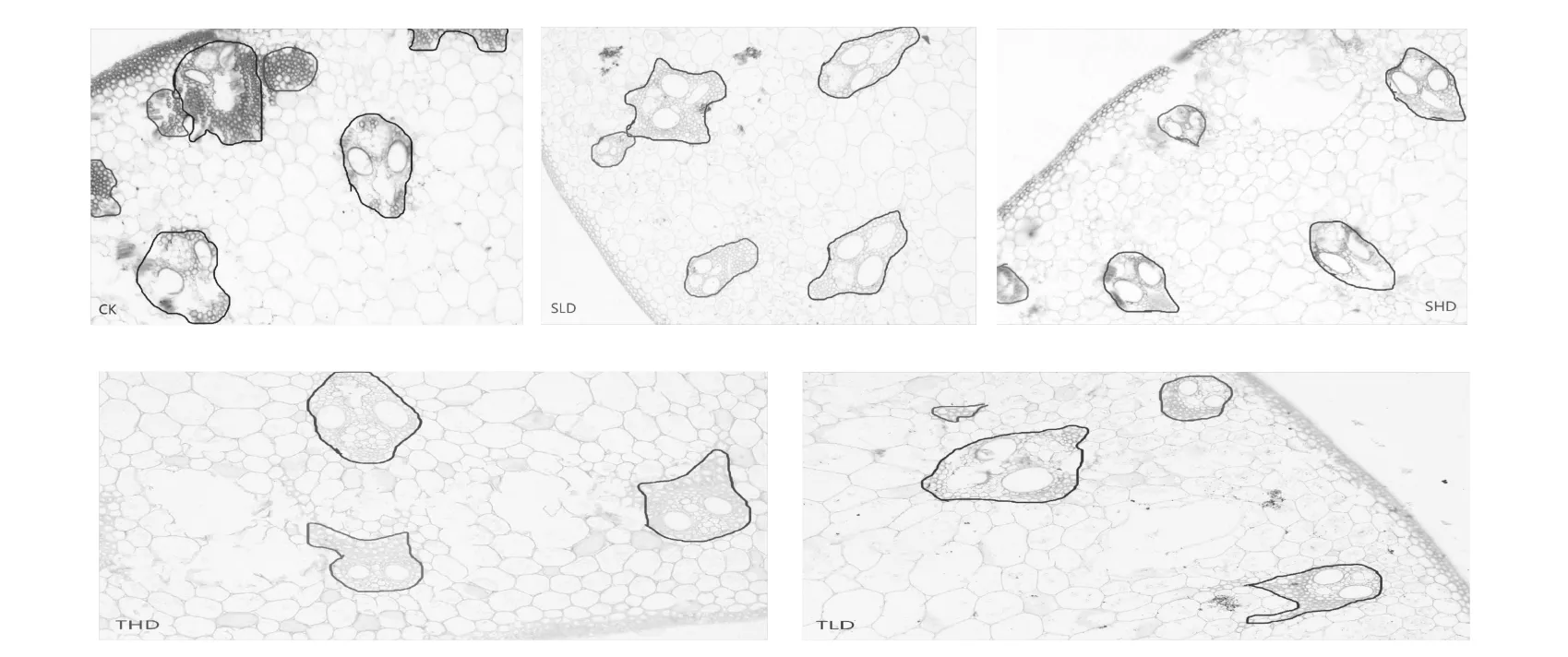
图1 细胞维管束Fig.1 Cells in the vascular bundle
2.3 旱涝交替胁迫对水稻干物质积累量及其抗折力的影响
相关研究发现作物不同生育阶段干物质积累对干旱胁迫的响应存在明显差异,干物质分配比率在一定程度上反映了水稻在受到水分胁迫时的生存对策[25]。分蘖期是以营养生长为主的时期,旱涝胁迫均抑制作物生长,导致干物质累积量减少。但水稻的生殖器官分化和生长主要发生在分蘖期以后,在其后生育阶段恢复正常水分管理,对水稻生殖器官的抑制作用较大。即在库源关系中,库的容量受到的影响较小,使得后继生育阶段中,更多的干物质向穗部运输(见表5),导致茎秆干物质累积减少。拔节期是水稻营养生长与生殖生长并行时期,旱涝胁迫会抑制幼穗发育及颖花分化,进而降低水稻的穗数和穗粒数,导致库的容量降低,对源的吸力降低,更多的干物质滞留于茎秆中,使得茎秆干物质累积量和所占比例提高,较分蘖期处理有提高的趋势。尤其是重度干旱胁迫处理,其干物质累积量和所占比例均显著高于对照和分蘖期处理。
单位长度茎秆的质量反映了茎秆充实度。Kashiwagi T等[26]认为40 cm以下茎秆干物质重与抗倒伏能力呈显著正相关。即茎秆抗折力与茎秆的充实度呈正相关。但在旱涝交替模式下,本试验与上述研究结果不完全吻合[10]。与对照和拔节期处理相比,分蘖期的旱涝胁迫使得茎秆干物质积累量显著下降,并与抗折力降低的趋势一致;拔节期不同程度的旱胁迫处理,其茎秆物质积累量和抗折力与对照无显著差异。在轻度干旱下,茎秆抗折力与茎秆干物质积累量成正相关(SLD和TLD);在重度干旱胁迫下,茎秆中干物质量的增加,未能提高茎秆的抗折力。这可能是重度旱胁迫处理大小维管束面积或数目的降低(见表4)、叶鞘厚度变薄等有关(见表3)。
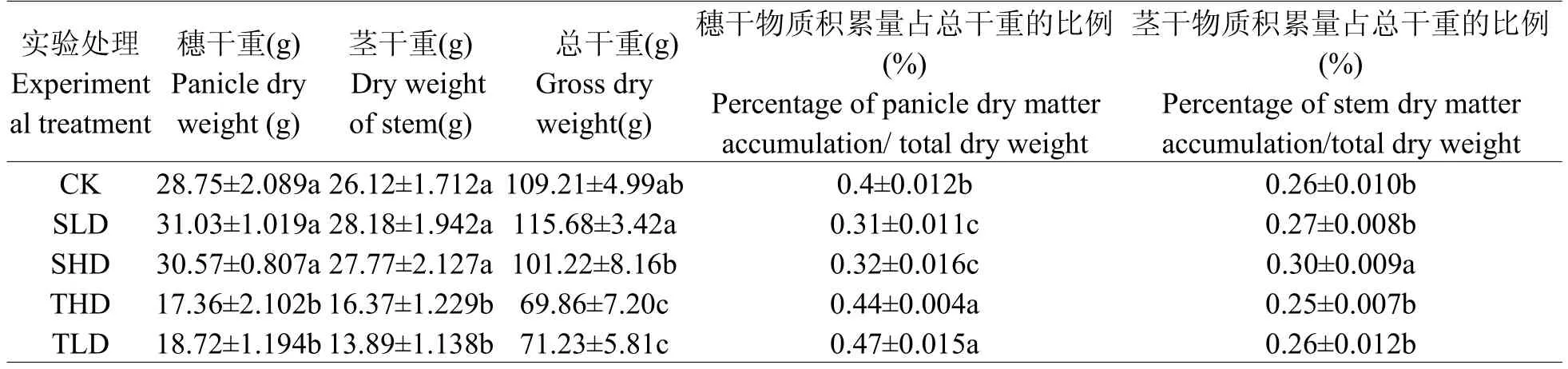
表5 不同旱涝交替胁迫对物质干积累量的影响Table 5 Different drought and flood alternating stress on the influence of dry matter accumulation
3 讨论与结论
茎秆倒伏是水稻减产的主要因素,为了降低水稻的倒伏,探讨了蓄水控灌模式下,旱涝交替胁迫对水稻抗折力的影响,通过对水稻分蘖期及拔节期的干旱胁迫处理,观察对穗重、茎秆厚度、叶鞘厚度、维管束的面积及数目、单位干物质重的影响,研究表明:茎鞘厚度、茎粗与抗折力无明显正相关,这与正常栽培下水稻其他学者研究观点不符,这可能旱涝交替胁迫改变了茎鞘成分或结构有关,有待于进一步研究。拔节期轻旱对水稻抗折力影响较大;重度旱胁迫无法提高水稻的抗折力。分蘖期的干旱胁迫能够促进水稻的增产,由于营养物质的运移导致叶鞘厚度及相应的单位干物质减少,虽然大维管束的面积有所增大,但总体抗折力无增加。
本试验中干旱胁迫对水稻基部倒伏能力的影响主要体现在两方面:一是通过改变茎秆大维管束面积或数目增加、使得水稻维管束内部的木质部中所含有的木薄壁细胞和木纤维的含量增加,提高水稻茎秆的硬度。二是通过抑制茎秆的延伸生长,提高茎秆充实度。上述因素综合使得水稻抗倒伏能力提高。
水稻基部抗折力是一个综合指标,其抗折力增减与胁迫方式有关。不同时期的旱涝交替胁迫,在水稻抗倒伏能力具有拮抗作用,可以降低彼此的不利影响,增强水稻茎秆的机械强度。为了增强茎秆的抗折断能力,必须在增加茎秆充实度的同时,进一步增强茎秆内在的微观结构。干旱胁迫具有提高水稻抗倒伏能力的作用,利用其拮抗效应,能够补偿淹水胁迫导致的抗倒伏能力下降。本试验表明:适当的轻度干旱和淹水胁迫能够提高水稻的抗折力;重度干旱胁迫未能增加水稻的抗倒伏能力。因此在蓄水控灌模式下对拔节期进行旱涝胁迫程度适宜控制,旱涝交替胁迫处理的水稻可获得较传统浅水勤灌更高的抗倒伏能力;为制定水稻合理灌排制度提供了理论依据,并对水稻抗倒伏及增产具有一定的指导意义。
[1]吕纯波,王福林,郭彦文.控制灌溉条件下水稻抗逆能力分析[J].中国农村水利水电,2006(10):139-141
[2]袁志华,赵安庆,苏宗伟,等.水稻茎秆抗倒伏的力学分析[J].生物数学学报,2003,18(2):234-237
[3]周丽华.水稻茎秆性状与抗倒伏关系的研究综述[J].中国稻米,2006(3):10-11
[4]杨长明,杨林章,颜廷梅,等.不同养分和水分管理模式对水稻抗倒伏能力的影响[J].应用生态学报,2004,15(4):646-650
[5]段传人,王伯初,王凭青.水稻茎秆的结构及其性能的相关性[J].重庆大学学报(自然科学版),2003,11:38-40
[6]杨艳华,朱镇,张亚东,等.不同水稻品种(系)抗倒伏能力与茎秆形态性状的关系[J].江苏农业学报,2011,27(2):231-235
[7]张忠旭,陈温福.水稻抗倒伏能力与茎秆物理性状的关系及其对产量的影响[J].沈阳农业大学学报,1999,30(2):81-85
[8]赵安庆,袁志华,曹晴.水稻茎秆抗倒伏的综合评价[J].生物数学学报,2006,21(4):554-556
[9]彭世彰,张正良,庞桂斌.控制灌溉条件下寒区水稻茎秆抗倒伏力学评价及成因分析[J].农业工程学报,2009,25(1):6-10
[10]杨长明,杨林章,颜廷梅,等.不同养分和水分管理模式对水稻抗倒伏能力的影响[J].应用生态学报,2004,15(4):646-650
[11]冯道俊.植物水涝胁迫研究进展[J].中国水运,2006,6(10):252-254
[12]胡田田,康绍忠.植物淹水胁迫响应的研究进展[J].福建农业大学学报(自然科学版),2005,34(1):18-24
[13]潘摇,澜摇,薛摇立.植物淹水胁迫的生理学机制研究进展[J].生态学杂志,2012,31(10):2662-2672
[14]郭相平,甄博,王振昌.旱涝交替胁迫增强水稻抗倒伏性能[J].农业工程学报,2013,29(12):130-135
[15]马均,马文波,田彦华,等.重穗型水稻植株抗倒伏能力的研究[J].作物学报,2004,30(2):143-148
[16]濑古秀生.水稻の倒伏に关する研究[J].九州农试汇报,1962,7:419-495
[17]杨惠杰,杨仁崔,李义珍,等.水稻茎秆性状与抗倒伏能力的关系[J].福建农业学报,2000,15(2):1-7
[18]金不换.干旱胁迫对不同品种早熟禾形态和生理特性影响的研究[D].哈尔滨:东北农业大学,2009
[19]郭相平,张烈君,王琴,等.作为水分胁迫补偿效应研究进展[J].河海大学学报(自然科学版),2005,33(6):634-647
[20]李阳生,李绍清.淹涝胁迫对水稻生长发育的生理特性和产量性状的影响[J].武汉植物学研究,2000,18(2):117-122
[21]郭相平,张烈君,王琴,等.拔节孕穗期水分胁迫对水稻生理特性的影响[J].干旱地区农业研究,2006,24(2):125-129
[22]潘学燕,苗芳.小麦维管组织结构研究概况[J].中国农学通报,2006,21(9):121-123
[23]胡婷,焦群英,付志一,等.小麦茎秆的抗弯复合材料力学模型[J].应用力学学报,2007,24(1):279-283
[24]王忠.植物生理学[M].北京:中国农业出版社,2000:221-256
[25]黄璜.水稻大穗形成机制研究-茎秆维管束数目与大穗形成的关系[J].湖南农业科学,1983,3(2):18-27
[26]李莹,张利民.植物甜菜碱及甜菜碱合成酶研究进展[J].杂粮作物,2006,26(3):191-193
Impact of Droughts Alternating Stress on the Fracture Resistance against Rice and Its Mechanism Analysis
YANG Jing-han,GUO Xiang-ping,YANG Sui,WANG Zhen-chang*
Key Laboratory of Efficient Irrigation-Drainage and Agricultural Soil-Water Environment in Southern China,Hohai University,Ministry of Education,Nanjing 210098,China
Rice tillering stage and jointing stage were studied by potted plant experiments of droughts alternating stress on rice after the microscopic structure,microstructure,stem sucks,and the influence law of bending force.The results showed that the stem diameter and leaf sheath thickness after drought stress were without a positive correlation with bending force. Dave bundle was positively correlated with the number of bending force,made the stem elongation stage light drought dry matter accumulation amount and the higher the percentage and the greater influence on the flexural strength of rice.The severe drought stress can't improve rice bending force.Tillering stage of drought stress while dave the area of the tube bundle increased,but not increased overall bending force.So water filling stages of rice control irrigation mode for appropriate control and drought stress level,can be obtained than traditional shallow water frequently filling and higher resistance to lodging.
Rice;alternative stress of drought and water logging(ASDW);culm physical characteristics;lodging index resistance snapping;pile-up effects
Q945.78
A
1000-2324(2014)03-0328-06
2012-10-23
2013-04-05
国家自然基金:滨海盐碱土隔离层条件下水盐不均匀分布及作物响应规律(51309080)
杨静晗(1990-),女,汉族,山东济宁人,硕士研究生,主要从事农业水土工程研究.
*通讯作者:Author for correspondence.E-mail:165548159@qq.com
猜你喜欢
水科学进展(2022年4期)2022-09-14
中国农业科学(2022年8期)2022-05-16
河北农业大学学报(2021年5期)2021-11-10
新疆农业科学(2020年9期)2020-10-13
水利规划与设计(2020年1期)2020-05-25
人民长江(2019年12期)2020-01-02
河南农业科学(2019年9期)2019-09-24
吉林农业·下半月(2017年7期)2017-07-15
农民致富之友(2017年6期)2017-05-10
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23
