
四种鱼蛉触角感器的超微形态
2014-06-12 03:10刘星月
植物保护 2014年5期
岳 璐, 刘星月, 杨 定
(中国农业大学农学与生物技术学院昆虫学系,北京 100193)
昆虫体表感受器是维持其正常生活行为的重要器官,而触角和口器是感受器分布最集中的结构[1-2]。触角作为昆虫最重要的感觉器官之一,其表面具有种类繁多的感受器,这些感受器有助于昆虫完成正常的生命活动,如识别、定位、觅食、求偶、交配、产卵、繁殖、栖息、防御与寻找寄主等[3-7]。同时,不同类群昆虫触角感器往往在形态、着生位置等方面存在显著差异,已有研究表明触角感器在昆虫分类中具有一定的应用价值[8-9]。因此,近年来,利用扫描电镜对昆虫触角感器超微结构观察的研究日益增加,并在昆虫形态分类及昆虫生理行为等方面的研究中起到了一定促进作用[10]。
鱼蛉亚科(Chauliodinae)隶属于昆虫纲广翅目(Megaloptera)齿蛉科(Corydalidae),其幼虫水生,捕食其他水生无脊椎动物,对蚊科、摇蚊科等害虫具有一定的控制作用。鱼蛉亚科世界性分布,目前已知17属约130种,我国已知5属46种。鱼蛉亚科触角形状和长度变异较大[11],为分属的重要特征,包括两大类型:触角丝状,约为前翅长的2/3,如异鱼蛉属(Apochauliodes)、始鱼蛉属(Archichauliodes)、原鱼蛉属(Protochauliodes)、马达鱼蛉属(Madachauliodes)、宽鱼蛉属(Platychauliodes)、梯鱼蛉属(Taeniochauliodes)、新鱼蛉属(Neohermes)、幻鱼蛉属(Nothochauliodes)、帝鱼蛉属(Dysmicohermes)和山鱼蛉属(Orohermes),其中新鱼蛉属雄蛉触角特化为念珠状;触角栉状或近锯齿状,约为前翅长的1/2,如臀鱼蛉属(Anachauliodes)、鱼蛉属(Chauliodes)、栉 鱼 蛉 属 Ctenochauliodes)、斑 鱼 蛉 属(Neochauliodes)、黑 鱼 蛉 属 (Nigronia)、准 鱼 蛉 属(Parachauliodes)和华鱼蛉属(Sinochauliodes)。在第二种类型中,触角形状在各属间又存在变异。在臀鱼蛉属、斑鱼蛉属和华鱼蛉属中,雄蛉触角为栉状而雌蛉近锯齿状;栉鱼蛉属雌雄蛉触角均栉状,只是雄蛉触角各节的分支更长;准鱼蛉属雌雄触角均近锯齿状;鱼蛉属触角雌雄二型或均栉状;黑鱼蛉属触角雌雄二型或均近锯齿状。鱼蛉亚科中的触角雌雄二型可能与其求偶过程中信息交流相关,触角感器可能在此过程中发挥作用并在不同属种间以及同种两性间存在差异[12]。此外,鱼蛉亚科作为较为古老原始的完全变态类昆虫,对其触角感器的研究将有助于理解触角的形态、功能乃至相关生物学行为在完全变态类昆虫中的演化。然而,目前有关鱼蛉亚科触角感器的研究仅见于杨定和刘星月对斑鱼蛉属的初步研究[12]。
笔者利用扫描电镜观察了鱼蛉亚科4个属的代表种:属模栉鱼蛉[Ctenchauliodes nigrovenosus(van der Weele)]、西华斑鱼蛉(Neochauliodes occidentalis van der Weele)、锯角黑鱼蛉[Nigronia serricornis (Say)]与全色新鱼蛉[Neohermes concolor(Davis)]成虫的触角感器,描述了4种鱼蛉触角感器的形态、种类、数量及分布,从不同属种间和同种雌雄两性间的触角感器特征进行比较。
1 材料与方法
1.1 虫源
本研究所用虫源均为95%乙醇浸泡标本。属模栉鱼蛉标本分别采自广西壮族自治区武鸣县大明山国家森林公园(雄)和贵州省遵义市宽阔水自然保护区白哨沟(雌);西华斑鱼蛉标本采自四川省峨眉山;锯角黑鱼蛉标本采自美国俄亥俄州 (USA,Ohio State,Delaware County,Alum Creek);全色新鱼蛉标本采自美国西弗吉尼亚州 (USA,West Virginia State,Hardy County,Northeastern Mathias)。
1.2 样品的制备与观察
取触角完整的4种鱼蛉雌雄各1头,在放大镜下用镊子和刀片将触角完整地取下,置于吸水纸上晾干,待标本完全干燥后,使用双面导电胶带将样品粘在样品台上,部分样品采用离子溅射器喷金。应用QUANTA 200F(荷兰FEI公司)扫描电子显微镜在15~20kV加速电压下观察和拍照。参考前人的研究对观察到的化学感受器进行鉴别和命名[1,3-10,12-16]。
2 结果与分析
2.1 触角一般形态
属模栉鱼蛉雌雄虫触角都呈栉状,鞭节各节具明显分支,基部和端部几节的分支短,中部的分支逐渐变长,雄虫分支平均为1 300~1 350μm,雌虫分支平均1 030~1 190μm。触角鞭节各亚节表面光滑无褶皱,而其分支表面具褶皱,且雄虫分支的褶皱比雌虫明显。鞭节各亚节长度相似,雄虫亚节长约288.1~305.2μm,雌虫亚节长约280.8~286.2μm。
西华斑鱼蛉雄虫触角呈栉状,鞭节各节具明显分支,但基部和端部几节的分支短,中部的分支逐渐变长,平均长约1 350μm,雌虫触角近锯齿状,即鞭节各亚节的外侧部分顶端向外突出,雌雄虫触角表面均无褶皱。雄虫触角鞭节亚节长约300.0μm,雌虫触角鞭节亚节长约315.7~326.9μm。
锯角黑鱼蛉雌雄虫触角均近锯齿状,雄虫触角齿状突伸较雌虫明显,雌雄虫触角表面均无褶皱。雄虫触角鞭节亚节长约338.9~350.0μm,基部与端部锯齿小,中部最大,突出部分宽度约94.4~97.2μm,雌虫触角鞭节亚节长约321.4~338.1μm。
全色新鱼蛉雄虫触角呈念珠状,雌虫触角呈丝状,表面光滑无褶皱。雄虫触角鞭节亚节长约366.7μm,雌虫触角鞭节亚节长约319.4~322.2μm。
2.2 属模栉鱼蛉触角感器
属模栉鱼蛉雌雄两性成虫触角具相同的5种感器,分别为:毛形感器Ⅱ型 (sensilla trichodeaⅡ,STⅡ)、毛形感器Ⅲ型 (sensilla trichodeaⅢ,STⅢ)、锥形感器 (sensilla basiconica,SB)、长锥形感器 (sensilla long-basiconica,SLB)、纽扣形感器 (sensilla buttonshaped,SBT)。主轴上5种感器均存在,触角分支上存在毛形感器Ⅱ型、毛形感器Ⅲ型、锥形感器和长锥形感器4种,无纽扣形感器 (图1)。

图1 属模栉鱼蛉雌雄成虫触角感器Fig.1 Sensilla on the antennae of adult males and females of Ctenchauliodes nigrovenosus
毛形感器 (sensilla trichodea,ST):毛形感器是属模栉鱼蛉触角上分布最多的感觉器,有2种类型,按形状分为Ⅱ型、Ⅲ型。Ⅱ型长于其他感器,劲直挺立,向顶端渐尖细,数量多,表面有明显的向上螺旋的纵沟,着生于触角鞭节各亚节的表皮毛窝中,在亚节主轴无分支一侧较多,而其在分支上集中分布于每个分支的下部,该种感器尖端在鞭节末端与分支末端处向内弯曲,长度约85.4~136.8μm,基部直径2.8~4.5μm;Ⅲ型直接生于触角各节,基部无凹形窝,呈弯曲纤毛状,较短,30°~60°角匍匐于触角表面,多数由基部伸向端部,长度约为11.2~15.7μm,基部直径1.1~1.7μm (图1:a,b,c)。
锥形感器 (sensilla basiconica,SB):锥形感器较短小,圆锥形,端部钝圆,垂直着生于触角鞭节各亚节表皮毛窝中,分支上较少,主要分布在触角亚节腹面两侧,长度约5.0~9.0μm,基部直径2.5~3.0μm(图1:a,b,c)。
长锥形感器 (sensilla long-basiconica,SLB):长锥形感器比锥形感器更粗长,近似锥形,端部钝圆,近垂直着生于触角鞭节各亚节表皮毛窝中,数目较少,在亚节端部具分支一侧少量分布,其在分支上的数量比主轴上更多,长度不等,基部直径3.4~4.2μm(图1:a,b,c)。
纽扣形感器 (sensilla button-shaped,SBT):形似纽扣,圆形,主要着生在触角鞭节各亚节端部以及亚节与分支的交界处,数量较少,感器直径4.2~5.7μm(图1:b,d)。
雌虫、雄虫在感器种类上无显著差别,毛形感器Ⅲ型分布最密、最多,其次是毛形感器Ⅱ型,其他3种感器相对较少。在数量上雌雄存在差异:在触角分支上,雄虫的毛形感器Ⅱ型比雌虫更多、更长,锥形感器与雌虫相比更多一些,其他感器差异不大;从每一节触角主轴上比较,雌虫毛形感器Ⅱ型与锥形感器略多于雄虫,其他感器数量相当。
2.3 西华斑鱼蛉触角感器
西华斑鱼蛉成虫触角感器共有5种,分别为:毛形感器Ⅰ型 (sensilla trichodeaⅠ,STⅠ)、毛形感器Ⅱ型 (sensilla trichodeaⅡ,STⅡ)、刺形感器 (sensilla chaetica,SC)、锥形感器 (sensilla basiconica,SB)和柱形感器(sensilla cylindric,SCL),其中雄虫触角具以上5种感器,而雌虫具其中4种感器且无刺形感器(图2)。
毛形感器 (sensilla trichodea,ST):毛形感器是在西华斑鱼蛉触角上分布最多的感器,有2种类型,按形状分为Ⅰ型、Ⅱ型。Ⅰ型着生于触角鞭节各亚节表皮毛窝中,短小,直毛状,表面光滑无纵沟,由基部伸向端部,长度约10.0~16.8μm,基部直径2.0~3.0μm;Ⅱ型着生于触角鞭节各亚节表皮毛窝中,劲直挺立,向顶端渐尖细,数量多,表面有明显向上螺旋的纵沟,长度约84.3~134.8μm,基部直径5.0~6.3μm (图2:a,b,c,d,f)。
刺形感器 (sensilla chaetica,SC):刺形感器主要着生在雄虫触角鞭节各亚节分支表皮毛窝中,细长,表面光滑无纵沟,端部钝圆,长为毛形感器Ⅰ型的2~3倍,约19.1~29.2μm,基部直径2.0~2.8μm(图2:a)。
锥形感器 (sensilla basiconica,SB):锥形感器较短小,圆锥形,端部钝圆,垂直着生于触角鞭节各亚节表皮毛窝中,各节上数量均不超过10个,长度约5.3~6.3μm,基部直径2.5~3.0μm (图2:c,d)。
柱形感器 (sensilla cylindric,SCL):柱形感器着生于触角鞭节各亚节表皮毛窝中,短粗,近圆柱形,末端钝圆,雌虫触角上分布不多,雄虫触角分支上相对较多,长度约为7.3~10.7μm,基部直径1.7~2.2μm (图2:a,c,e)。
比较雌雄虫触角鞭节的一个亚节,发现雄虫触角感器比雌虫更密集。雌虫的毛形感器Ⅱ型较雄虫多,雄虫在有分支的一侧几乎无毛形感器Ⅱ型而在其相对一侧则较多;毛形感器Ⅰ型在雌虫触角各亚节端部的侧面明显居多,雄虫则在近分支处居多,且与其他部分密度差异不大。从毛形感器Ⅰ型与毛形感器Ⅱ型在一个亚节上整体数量比例来看,雌虫为7∶10,而雄虫为10∶3。
在雄虫触角分支上,毛形感器Ⅰ型、毛形感器Ⅱ型和刺形感器整体密度较高,毛形感器Ⅰ型与毛形感器Ⅱ型数量较刺形感器明显减少,三者数量比例 (毛形感器Ⅰ型:毛形感器Ⅱ型:刺形感器)约为5∶1∶15。
2.4 锯角黑鱼蛉触角感器
锯角黑鱼蛉雌雄两性成虫触角具有相同的4种感器,分别为:毛形感器Ⅲ型 (sensilla trichodeaⅢ,STⅢ)、毛形感器Ⅳ型 (sensilla trichodeaⅣ,STⅣ)、锥形感器 (sensilla basiconica,SB)和柱形感器(sensilla cylindric,SCL)(图3)。
毛形感器 (sensilla trichodea,ST):按形状分为Ⅲ型、Ⅳ型。毛型感器Ⅲ型直接生于触角各节,基部无凹形窝,呈弯曲纤毛状,较短,30°~60°角匍匐于触角表面,多数由基部伸向端部,长度约为12.2~19.0μm,基部直径1.4~1.7μm;毛形感器Ⅳ型着生于触角鞭节各亚节表皮毛窝中,与Ⅱ型形状类似,长度较Ⅱ型短,劲直挺立,向顶端渐尖细,表面明显有向上纵沟,长约30.0~59.4μm,基部直径4.4~5.0μm (图3:a,b,c,d)。
锥形感器 (sensilla basiconica,SB):锥形感器相对短小,但该种鱼蛉触角的锥形感器比本文观察其他各种者略长,近圆锥形,端部钝圆,垂直着生于触角鞭节各亚节表皮毛窝中,雌虫各节数量均不超过10个,雄虫上数量相对较多,均匀分布在亚节各面,长约4.6~9.7μm,基部直径2.2~2.6μm (图3:a,b,d)。
柱形感器 (sensilla cylindric,SCL):该种鱼蛉触角的柱形感器比本文观察其他各种者略短,雌虫触角上很少,雄虫上的数量与锥形感器相当,长度约6.0~6.7μm,基部直径1.9~2.3μm (图3:a,b,c)。
比较雌雄虫触角鞭节的一个亚节,雌虫与雄虫感器分布密度相当。雌虫毛形感器Ⅳ型在触角上分布比较均匀,而雄虫毛形感器Ⅳ型主要分布在触角腹面的中部与侧面,总体数目比雌虫少;雌雄虫毛形感器Ⅲ型均主要在毛形感器Ⅳ型之间均匀分布,其次在各亚节连接部位也分布较密,雌虫上该感器的长度比雄虫略长;在柱形感器方面,雄虫柱形感器明显多于雌虫,雌虫的柱形感器和锥形感器零散地分布在毛形感器Ⅲ型与毛形感器Ⅳ型中间,而雄虫触角主轴上这两种感器较多,数量与其上的毛形感器Ⅲ型相当,在锯齿突出处略有增多。

图2 西华斑鱼蛉雌雄成虫触角感器Fig.2 Sensilla on the antennae of adult males and females of Neochauliodes occidentalis
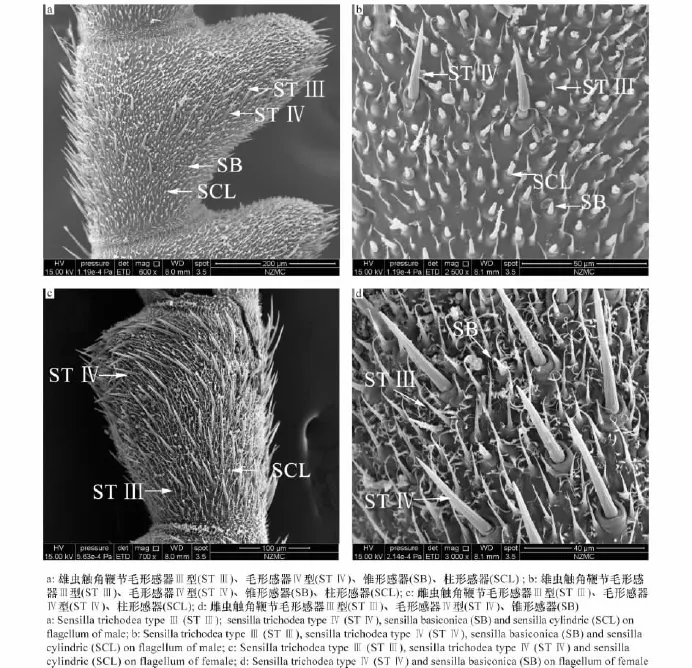
图3 锯角黑鱼蛉雌雄成虫触角感器Fig.3 Sensilla on the antennae of adult males and females of Nigronia serricornis
2.5 全色新鱼蛉触角感器
全色新鱼蛉成虫触角感器共有5种,分别为毛形感器Ⅱ型 (sensilla trichodeaⅡ,STⅡ)、毛形感器Ⅴ型(sensilla trichodea V,ST V)、柱形感器(sensilla cylindric,SCL)、锥形感器 (sensilla basiconica,SB)和长锥形感器 (sensilla long-basiconica,SLB),其中雄虫触角具以上5种感器,而雌虫具其中4种感器且无毛形感器Ⅴ型(图4)。
毛形感器 (sensilla trichodea,ST):按形状分为Ⅱ型、Ⅴ型。毛形感器Ⅱ型直立着生于表皮窝中,顶端渐细,表面有明显纵沟,长度约77.8~130.5μm,基部直径5.0~6.9μm;毛形感器Ⅴ型着生于触角表面表皮窝中,纤细弯曲,比Ⅱ型更长,长度与亚节长度相当或略长于亚节,其上有螺旋纵纹,长度约291.0~375.0μm,基部直径8.3~9.2μm(图4:a,b,c,d)。
柱形感器 (sensilla cylindric,SCL):雌雄虫触角上都比较少,分布在锥形感器中间,长度约7.8~10.0μm,基部直径2.5~3.3μm (图4:a,c,d)。
锥形感器 (sensilla basiconica,SB):锥形感器相对短小,雌虫触角腹面分布较多,长度约4.2~8.3μm,基部直径3.1~4.2μm(图4:a,b,c,d)。
长锥形感器 (sensilla long-basiconica,SLB):在雌虫触角两侧分布较多,雄虫触角上有少数该种感器,长度约13.8~25.0μm,基部直径3.3~4.2μm(图4:b,c,d)。

图4 全色新鱼蛉雌雄成虫触角感器Fig.4 Sensilla on the antennae of adult males and females of Neohermes concolor
比较雌雄虫触角鞭节的一个亚节,雄虫触角感器的分布密度明显小于雌虫。在雄虫触角亚节上,毛形感器Ⅱ型很少,毛形感器Ⅴ型最多,环绕分布在触角各面,柱形感器比较少,锥形感器比柱形感器多,在毛形感器Ⅴ型中间不均匀分布。雌虫触角亚节上,均匀分布较多的毛形感器Ⅱ型,长锥形感器主要分布在亚节侧面,锥形感器主要分布在亚节腹面,相对于锥形感器和长锥形感器,柱形感器比较稀少。从触角整体上看,柱形感器、锥形感器、长锥形感器这3种感器在末端节分布较少,在柄节、梗节和鞭节的前几节分布明显增多。
3 讨论
在感器类型方面,属模栉鱼蛉和锯角黑鱼蛉雌雄两性触角感器无差异;西华斑鱼蛉雄虫触角具有毛形感器Ⅰ型、毛形感器Ⅱ型、刺形感器、锥形感器和柱形感器,雌虫触角则缺少刺形感器;全色新鱼蛉雄虫触角具有毛形感器Ⅱ型、毛形感器Ⅴ型、柱形感器、锥形感器和长锥形感器,雌虫触角则缺少毛形感器Ⅴ型。由此可见,触角形态具有明显雌雄二型的种类,在触角感器类型上也具有性别间的差异[13]。4种鱼蛉雌雄虫触角上共同具有的唯一一种感器是锥形感器,属模栉鱼蛉、西华斑鱼蛉和全色新鱼蛉雌雄虫触角均具毛形感器Ⅱ型,西华斑鱼蛉、锯角黑鱼蛉和全色新鱼蛉雌雄虫触角均具柱形感器。属模栉鱼蛉触角特有纽扣形感器,西华斑鱼蛉触角上特有毛形感器Ⅰ型,西华斑鱼蛉雄虫触角上还特有刺形感器,锯角黑鱼蛉触角特有毛形感器Ⅳ型,全色新鱼蛉只有雄虫触角上特有毛形感器Ⅴ型。
在触角鞭节各亚节全部感器整体密度上比较,锯角黑鱼蛉触角感器密度最高,其次是属模栉鱼蛉和西华斑鱼蛉,全色新鱼蛉密度最小。分布于北美的黑鱼蛉属与亚洲的鱼蛉近缘,本研究中发现的这些属代表种具密集的触角感器也可能是反映其近缘关系的特征之一。在同种两性间,属模栉鱼蛉和锯角黑鱼蛉雌雄虫触角感器密度相当,而西华斑鱼蛉雄虫触角感器密度大于雌虫,全色新鱼蛉雄虫触角感器密度小于雌虫。因此表明触角形态具有明显雌雄二型的种类,在触角感器密度上也具有性别间的差异。
在各感器的数量与分布上比较,锥形感器在属模栉鱼蛉触角分支上少量分布且雄虫者略多于雌虫,在触角主轴腹面两侧相对分支略多且雌虫者多于雄虫;西华斑鱼蛉雌雄触角各亚节锥形感器均不超过10个;锯角黑鱼蛉雌虫触角各亚节锥形感器不超过10个,雄虫者稍多,尤其是锯齿状突出部分数量明显增多;全色新鱼蛉锥形感器主要分布在触角腹面。据报道锥形感器的功能主要集中在机械感受和嗅觉上[4,14-15],在这4种鱼蛉上锥形感器数量均不多,且同种雌雄差异不明显,因此4种鱼蛉触角锥形感器可能也具有类似的机械感受和嗅觉功能。柱形感器为西华斑鱼蛉、锯角黑鱼蛉和全色新鱼蛉所共有,但数量较少,雄虫触角上该感器明显多于雌虫,表明柱形感器在西华斑鱼蛉、锯角黑鱼蛉和全色新鱼蛉中可能具有接受雌虫性信息素的功能。毛形感器作为最常见的触角感器,有报道表明具有机械受体和触觉受体双重功能[16],此外还有接收性信息素的功能[17]。毛形感器Ⅱ型和毛形感器Ⅲ型在4种鱼蛉上数量较多,因此可能具有机械受体和触觉受体的功能,全色新鱼蛉雄虫特有的毛形感器Ⅴ型则可能具有接收同种雌虫性信息素的功能。
本文对鱼蛉亚科4个属代表种触角感器超微形态的观察表明,触角感器在鱼蛉亚科不同属种间无论在类型还是数量分布等方面均存在差异且多样化。同时,同种雌雄两性的触角感器在一些具有雌雄触角二型性的物种中也具有差异,表明触角感器在鱼蛉亚科求偶交配过程中确实发挥了信息交流的作用。但目前有关鱼蛉亚科乃至整个广翅目触角感器在生理生态方面的作用机制还尚未研究。今后有必要对广翅目更多代表属种以及同种更多个体的触角感器进行观察,并深入探讨触角感器作用的生理机制。
[1]Schneider D.Insect antennae[J].Annual Review of Entomology,1964,9:103-122.
[2]Zhao L Q,Dai W,Zhang C N,et al.Morphological characterization of the mouthparts of the vector leafhopper Psammotettix striatus (L.)(Hemiptera:Cicadellidae)[J].Micron,2010,41:754-759.
[3]王桂荣,郭予元,吴孔明.棉铃虫触角感器的超微结构观察[J].中国农业科学,2002,35(12):1479-1482.
[4]那杰,于维熙,李玉萍,等.昆虫触角感器的种类及其生理生态学意义[J].沈阳师范大学学报(自然科学版),2008,26(2):213-216.
[5]王攀,郑霞林,雷朝亮,等.豇豆荚螟幼虫头部形态及化学感受器观察[J].植物保护,2011,37(2):72-75.
[6]洪慧金,任顺祥,杜予州,等.三种粉虱触角、口针及跗节感受器的超微结构观察[J].应用昆虫学报,2011,48(1):77-83.
[7]Zhang D.Wang Q K,Hu D F,et al.Sensilla on the antennal funiculus of the horse stomach bot fly,Gasterophilus nigricornis [J].Medical and Veterinary Entomology,2012,26:314-322.
[8]王娜,魏劲松,党露,等.三种夜蛾成虫口器感器的超微形态[J].昆虫学报,2012,55(7):877-884.
[9]王颖娟,李子忠.锈翅蚁蛉头部附器的扫描电镜观察[J].应用昆虫学报,2012,49(6):1636-1642.
[10]李竹,陈立.触角感器特征应用于昆虫分类的研究进展[J].昆虫分类学报,2008,32:113-118.
[11]刘星月,李文亮,杨定.脉翅总目系统发育研究进展[J].昆虫知识,2007,44(5):626-631.
[12]杨定,刘星月.中国动物志,昆虫纲,第51卷,广翅目[M].北京:科学出版社,2010.
[13]杨啸,赵奎军,王克勤,等.大豆食心虫触角感器的扫描电镜观察[J].应用昆虫学报,2012,49(5):1321-1326.
[14]马瑞燕,杜家伟.昆虫的触角感器[J].昆虫知识,2000,37(3):179-182.
[15]支海美,刘星月,杨定.苹果蠹蛾头部感器的电镜扫描结构[J].应用昆虫学报,2012,49(1):43-48.
[16]余海忠.昆虫触角感受器研究进展[J].安徽农业科学,2007,35(14):4238-4240,4243.
[17]Hallberg E,Hansson B S,Steinbrecht R A.Morphological characteristics of antennal sensilla in the European cornborer Ostrinia nubilalis(Lepidoptera:Pyralidae)[J].Tissue Cell,1994,26:489-502.
猜你喜欢
广西林业科学(2022年1期)2022-03-03
生物学杂志(2021年4期)2021-08-24
甘肃农业科技(2021年6期)2021-07-25
环境昆虫学报(2020年5期)2020-11-11
延安大学学报(自然科学版)(2019年3期)2019-10-11
科学导报(2019年35期)2019-09-03
河南农业科学(2019年5期)2019-05-28
植物保护(2018年3期)2018-05-14
江苏农业科学(2014年3期)2014-07-16
