
miR-31对人皮肤鳞状细胞癌生长的影响及作用机制的研究
2014-05-25 05:49:58常东方段绍坤李渝凉
中国肿瘤临床 2014年9期
陈 凯 常东方 段绍坤 李渝凉
皮肤鳞状细胞癌(鳞癌)是我国常见的皮肤恶性肿瘤,具有高度的侵袭能力,可能与紫外照射损伤、免疫抑制及其他皮肤疾病有关[1]。因此,寻找其发生过程中发挥关键作用的分子对其治疗是必要的。研究发现微小RNA(microRNA,miRNA)在肿瘤的发生发展中起着重要的作用[2]。
miRNA是一类比较保守的单链非编码小RNA分子,主要通过与靶基因mRNA 3'UTR结合,在转录后水平调控基因的表达[3,4]。miRNA参与一系列的生理过程,如生长、发育、分化及凋亡[5]。研究发现约50%miRNA基因定位于染色体上肿瘤基因相关区域,并且在肿瘤中经常伴随着miRNA的异常表达[6]。关于miRNA与皮肤鳞癌的研究在国内外报道甚少,有研究表明在皮肤鳞癌中miR-125b通过靶定MMP-13抑制鳞癌细胞的生长和侵袭迁移[7]。此外,通过对皮肤鳞癌和其他皮肤肿瘤患者中miRNA表达水平的分析,发现miR-31在皮肤鳞癌中高表达[8]。然而,关于miR-31在皮肤鳞癌中的作用还未明确。
本研究探索了miR-31对细胞增殖以及肿瘤生长的影响,并对其靶基因进行验证。
1 材料与方法
1.1 材料
本研究20对皮肤鳞癌和癌旁正常组织来自2011年1月至2013年6月重庆医科大学永川医院收治患者,并得到患者的知情同意。鳞癌细胞A431和SLC-1的培养液是含有10%FBS的DMEM(Gibco公司)。脂质体LipofectamineTM2000购自Invitrogen公司,ASO和siRNA购自上海吉玛公司。兔抗LATS2和GAPDH抗体购自Abcam公司。RNA提取试剂盒购自Qiagen公司,反转录酶购自Fermentas公司,实时定量PCR的2×SYBR Green Master Mix购自Takara公司。
1.2 方法
1.2.1 细胞转染 按照脂质体的说明进行。细胞转染4 h后,换含有血清的培养液,在37℃、5%CO2的孵箱中继续培养。
1.2.2 平板克隆形成实验 将转染的细胞种到12孔板,每隔3天换液至到大部分克隆形成。用PBS清洗后5%浓度结晶紫染色,并进行计数。
1.2.3 体外成瘤实验 裸鼠购自北京维通利华有限责任公司(动物合格证号:SCXK(京)2013-0002),并得到动物管理委员会的许可。裸鼠在无菌的条件下正常饮食,将106个转染的鳞癌细胞重悬在100 μL无血清培养液中,经皮下注射途径植入裸鼠。大概3周后,将小鼠麻醉处死取出肿瘤组织,并量取体积。体积=(长×宽2)/2。
1.2.4 Western印迹 细胞中转染LATS2的siRNA(20 nM)以及对照,48 h后提取蛋白进行Western印迹分析LATS2的水平。转染48 h后,用PBS洗细胞后用RIPA缓冲液裂解细胞。利用BCA法进行蛋白浓度测定,取20 μg蛋白进行SDS-PAGE胶分析。最后利用ECL显影,并分析条带的强度。以GAPDH作为内参。
1.2.5 RNA提取和实时定量PCR 利用RNA提取试剂盒进行RNA提取,采用NanoDrop ND-1000分光光度计测定RNA的浓度。取500 ng RNA进行cDNA的合成后进行PCR反应。反应条件是:95℃30 s,之后40个循环95℃ 30 s,57℃ 30 s,72℃ 30 s。以U6 RNA和β-actin作为内参。
1.2.6 免疫组织化学 将石蜡包埋的组织切片进行脱蜡水合处理,3%H2O2室温孵育5 min。蒸馏水冲洗,PBS浸泡5 min后,用5%山羊血清封闭10 min,滴加一抗工作液孵育1 h。然后用生物素标记的二抗工作液37℃孵育10 min。最后滴加辣根酶标记的链霉卵白素工作液,37℃孵育30 min。PBS冲洗后显色剂显色3 min,在显微镜下观察。
1.2.7 GFP报告载体实验 将含有miR-31结合位点的LATS2 3'UTR克隆到pcDNA3/GFP载体的GFP下游。在细胞中共转染miR-31 ASO和LATS2 3'UTR,同时转染RFP质粒作为内参。转染48 h后,用RIPA裂解细胞提取蛋白,荧光分光光度计检测细胞GFP荧光强度。
1.3 统计学处理
用SPSS 18.0软件进行统计分析,比较两组数据间的差异采用双侧样本的Student's t检验,比较相关性采用Pearson相关系数分析。P<0.05被认为是差异具有统计学意义。
2 结果
2.1 平板克隆形成实验分析miR-31对细胞生长的影响
将miR-31 ASO以及对照转染A431和SLC-1细胞,利用平板克隆形成实验检测细胞生长的情况。转染miR-31 ASO的A431细胞克隆数较对照组减少约50%,且差异具有统计学意义(P<0.05);在SLC-1细胞中也得到了类似的结果。
2.2 体外成瘤实验分析miR-31对肿瘤生长的影响
将转染miR-31 ASO以及对照的细胞经皮下注射入裸鼠体内,利用体内成瘤模型检测miR-31对肿瘤生长的影响。结果显示转染miR-31 ASO的鳞癌细胞成瘤能力降低,肿瘤体积明显小于对照组(图1),且差异具有统计学意义(P<0.05)。
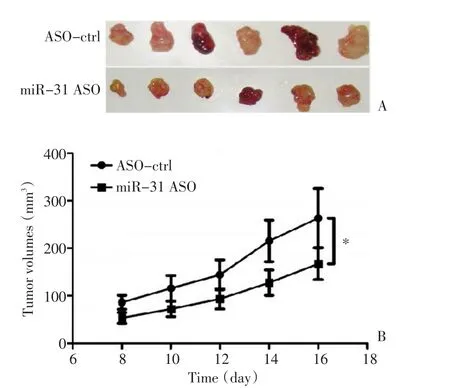
图1 体外成瘤实验检测miR-31对鳞癌生长的影响Figure 1 In vitrotumor formation assay indicates that miR-31 inhibition suppresses cSCC growth
2.3 Western blot印迹检测miR-31对LATS2蛋白水平的影响
研究证实miR-31可以靶定抑癌基因LATS2和PPP2R2A[9]。为了检测在皮肤鳞癌中miR-31是否也通过这两个基因发挥作用,首先通过Western印迹检测miR-31 ASO对LATS2和PPP2R2A表达水平的影响。实验显示鳞癌细胞转染miR-31 ASO后使LATS2的水平升高约1.5倍,差异具有统计学意义(图2,P<0.05),而对PPP2R2A的表达水平无影响(P>0.05)。
2.4 生物信息学预测miR-31的候选靶基因
利用生物信息学方法(TargetScan以及Pictar预测网站)对miR-31与候选靶基因的结合位点进行验证,图3显示miR-31与候选靶基因LATS2 3'UTR的结合位点互补。
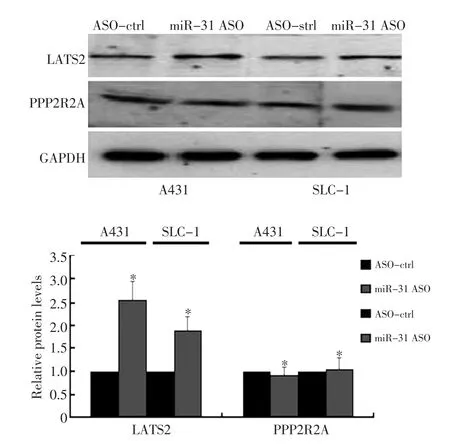
图2 Western blot检测miR-31对候选靶基因蛋白表达的影响Figure 2 Western blot shows that miR-31 inhibition increases LATS2 protein levels
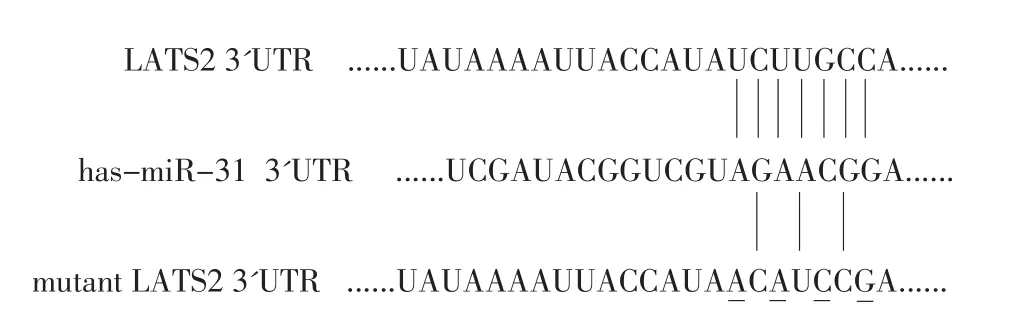
图3 生物信息学预测miR-31的靶基因Figure 3 Bioinformatics indicates that LATS2 3'UTR has binding sites with miR-31
2.5 GFP报告载体实验验证miR-31的靶基因
在A431和SLC-1细胞中共转染miR-31 ASO和野生型或者突变型的LATS2 3'UTR(突变位点如图3所示),通过GFP报告载体实验检测miR-31对LATS2 3'UTR荧光强度的影响。结果显示转染miR-31 ASO的细胞中LATS2 3'UTR的荧光强度较对照组增强50%(图4),且差异具有统计学意义(P<0.05),且在两种鳞癌细胞中结果一致。然而miR-31 ASO对突变的3'UTR荧光强度无明显影响(P>0.05,图4)。
2.6 Western blot检测siRNA对LATS2表达水平的影响
在A431和SLC-1细胞中转染LATS2 siRNA以及对照,转染48 h后利用Western blot检测细胞中LATS2蛋白水平。结果显示转染LATS2 siRNA的细胞中LATS2的水平较对照组降低约80%,且差异具有统计学意义(图5,P<0.05)。
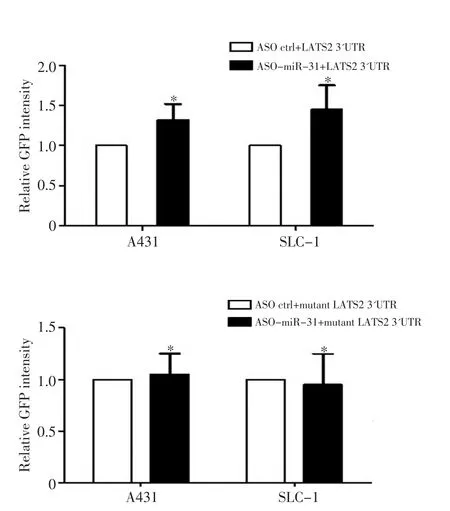
图4 GFP报告实验验证miR-31的靶基因Figure 4 GFP reporter assay shows that miR-31 inhibition increases the GFP intensity of LATS2 3'UTR
2.7 平板克隆形成实验分析LATS2对细胞生长的影响
在A431和SLC-1细胞中转染LATS2 siRNA以及对照,利用平板克隆形成实验检测细胞的生长情况,结果显示转染LATS2 siRNA的细胞克隆数分别升高约70%和1.3倍,且差异具有统计学意义(图6,P<0.05)。
2.8 实时定量PCR分析miR-31的表达水平
利用实时定量PCR检测miR-31和靶基因在鳞癌以及癌旁正常组织中的表达水平。图7显示,与癌旁正常组织相比,鳞癌组织中miR-31的水平升高约2.5倍,差异具有统计学意义(P<0.05),并且miR-31与LATS2的表达呈负相关(P<0.05)。
2.9 免疫组织化学法检测LATS2在鳞癌组织中的表达
利用免疫组织化学检测LATS2在鳞癌组织和癌旁正常组织中的表达水平。图8显示,与正常组织相比,皮肤鳞癌组织中LATS2的表达水平明显降低。
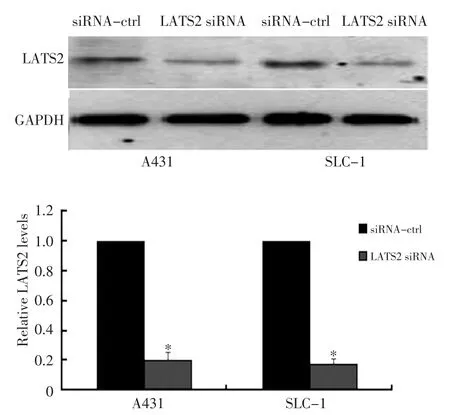
图5 Western blot检测LATS2的表达以及平板克隆形成实验检测细胞生长Figure 5 Western blot shows that LATS2 expression is repressed by in⁃troduction of siRNA against LATS2
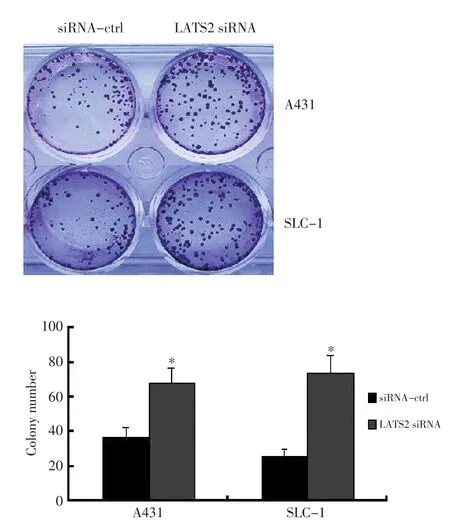
图6 平板克隆形成实验检测LATS2对细胞生长的影响Figure 6 Colony formation assay shows that knocking down of LATS2 in⁃creases the number of colonies
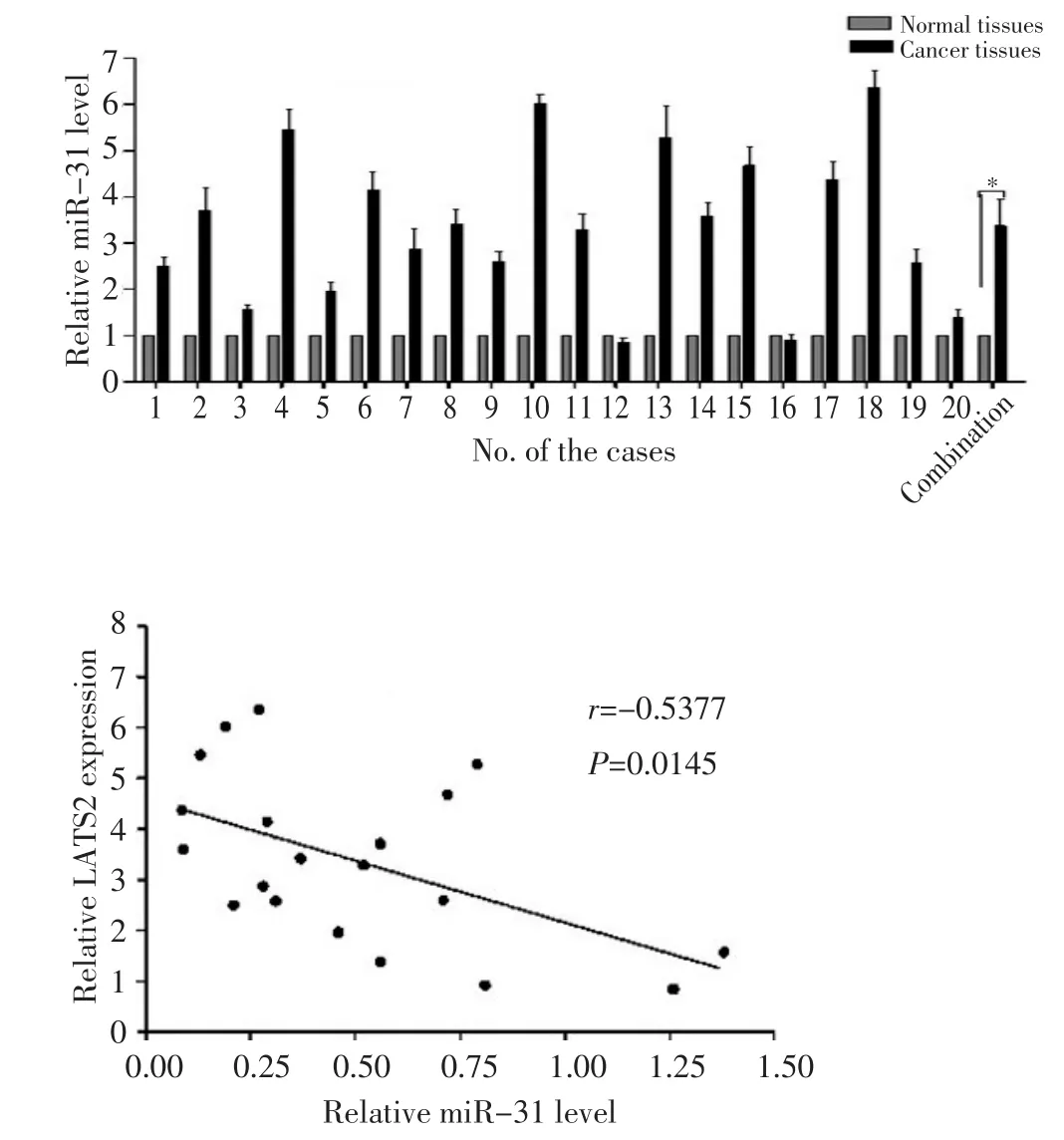
图7 实时定量PCR检测miR-31和LATS2在鳞癌组织和正常对照中的表达水平Figure 7 Real-time PCR shows that miR-31 is upregulated in cSCC tis⁃sues,and an inverse relationship exists between miR-31 and LATS2 ex⁃pression levels
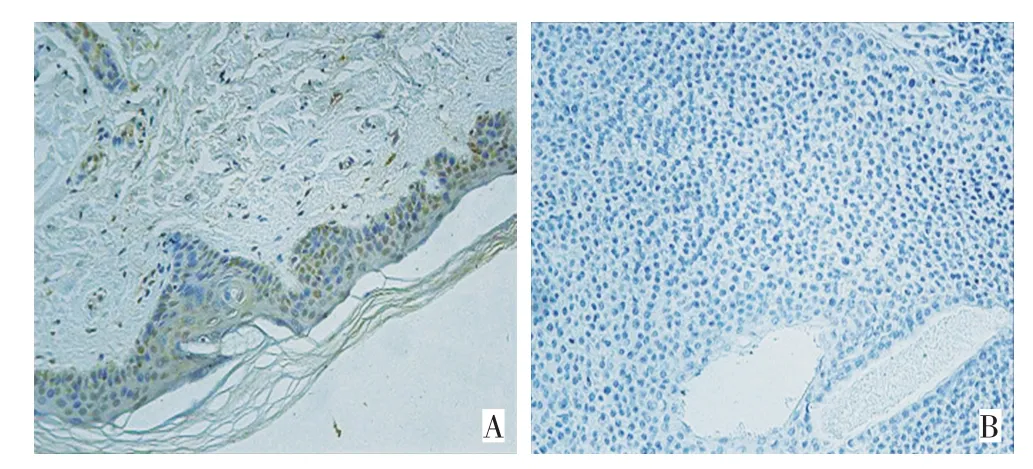
图8 免疫组织化学检测LATS2在鳞癌组织和正常对照中的表达水平(SP×200)Figure 8 Immunohistochemistry shows that LATS2 is downregulated in cSCC tissues(SP×200)
3 讨论
本研究结果发现miR-31在鳞癌组织中高表达,抑制miR-31的表达后细胞克隆形成能力下降,肿瘤生长受抑制。已有研究发现miR-31可促进血管平滑肌细胞生长[10]。此外,在食管鳞癌和结肠癌中,miR-31也是高表达,并且与肿瘤的进展程度相关[11-12],这些均提示miR-31发挥着癌基因的角色。
miRNA与靶基因3'UTR的不完全互补,是导致一个miRNA调控多个靶基因表达的原因之一[13]。研究证实在肺癌和血管平滑肌细胞中miR-31可以通过靶定PPP2R2A和LATS2参与细胞的生长。在本研究中,抑制miR-31的表达后,LATS2蛋白水平升高,而PPP2R2A的表达不受影响。这可能是PPP2R2A的3'UTR在不同的肿瘤细胞中有差异,导致miRNA的抑制作用消失。此外,通过GFP报告实验进一步验证LATS2是miR-31的直接靶基因,miR-31通过与其3'UTR直接结合而抑制其表达。
LATS2定位于13号染色体,可编码丝氨酸苏氨酸蛋白激酶。研究发现该酶属于Hippo肿瘤抑制信号途径的成员,可以通过磷酸化的形式使癌基因YAP失活,从而抑制恶性间皮瘤细胞的生长[14]。在胃癌中,癌基因miR-372可以通过抑制LATS2的表达从而抑制细胞的生长,促进细胞凋亡[15]。说明LATS2在抑制肿瘤细胞生长中起着重要的作用。本研究通过免疫组织化学及实时定量PCR发现LATS2在皮肤鳞癌组织中表达降低。敲除LATS2的表达后细胞的克隆形成能力增强,与抑制miR-31的作用相反,进一步证实LATS2是miR-31的靶基因。
总之,本研究在皮肤鳞癌中miR-31通过抑制LATS2的表达调控鳞癌的生长,发挥着癌基因的作用,可为皮肤鳞癌的分子治疗提供更多的依据和理论基础。
1 Garcia-Zuazaga J,Olbricht SM.Cutaneous squamous cell carcino⁃ma[J].Adv Dermatol,2008,24:33-57.
2 Du B,Wang Z,Zhang X,et al.MicroRNA-545 Suppresses Cell Proliferation by Targeting Cyclin D1 and CDK4 in Lung Cancer Cells[J].PloS One,2014,9(2):e88022.
3 Bartel DP.MicroRNAs:genomics,biogenesis,mechanism,and function[J].Cell,2004,116(2):281-297.
4 Dalmay T.Mechanism of miRNA-mediated repression of mRNA translation[J].Essays Biochem,2013,54:29-38.
5 Ambros V.The functions of animal microRNAs[J].Nature,2004,431(7006):350-355.
6 Calin GA,Sevignani C,Dumitru CD,et al.Human microRNA genes are frequently located at fragile sites and genomic regions in⁃volved in cancers[J].Proc Natl Acad Sci U S A,2004,101(9):2999-3004.
7 Xu N,Zhang L,Meisgen F,et al.MicroRNA-125b down-regu⁃lates matrix metallopeptidase 13 and inhibits cutaneous squamous cell carcinoma cell proliferation,migration,and invasion[J].J Biol Chem,2012,287(35):29899-29908.
8 Bruegger C,Kempf W,Spoerri I,et al.MicroRNA expression differs in cutaneous squamous cell carcinomas and healthy skin of immu⁃nocompetent individuals[J].Exp Dermatol,2013,22(6):426-428.
9 Liu X,Sempere LF,Ouyang H,et al.MicroRNA-31 functions as an oncogenic microRNA in mouse and human lung cancer cells by repressing specific tumor suppressors[J].J Clin Invest,2010,120(4):1298-1309.
10 Liu X,Cheng Y,Chen X,et al.MicroRNA-31 regulated by the ex⁃tracellular regulated kinase is involved in vascular smooth muscle cell growth via large tumor suppressor homolog2[J].J Biol Chem,2011,286(49):42371-42380.
11 Luo J,Ling ZQ,Peng BF,et al.Expression of miR-31 in esopha⁃geal squamous cell carcinomaand its relation to prognosis[J].Chi⁃nese Journal of Cancer,2013,(7):487-492.[罗 君,凌志强,彭兵锋,et al.MicroRNA-31在食管鳞状细胞癌中的表达及其与预后的关系[J].中国癌症杂志,2013,(7):487-492.]
12 Wang CJ,Ma N,Zhou Y.Expression and functions of miR-31 in colorectal cancer tissues and cells[J].Journal of zhengzhou universi⁃ty(Medical medicine),2013,(5):625-629.[王朝杰,马 宁,周 云.结直肠癌组织和结肠癌细胞中miR-31的表达及功能[J].郑州大学学报(医学版),2013,(5):625-629.]
13 Carroll AP,Goodall GJ,Liu B.Understanding principles of miRNA target recognition and function through integrated biological and bioinformatics approaches[J].Wiley interdiscip Rev RNA,2014,doi:10.1002/wrna.1217.[Epub ahead of print]
14 Murakami H,Mizuno T,Taniguchi T,et al.LATS2 is a tumor sup⁃pressor gene of malignant mesothelioma[J].Cancer Res,2011,71(3):873-883.
15 Cho WJ,Shin JM,Kim JS,et al.miR-372 regulates cell cycle and apoptosis of ags human gastric cancer cell line through direct regula⁃tion of LATS2[J].Mol Cells,2009,28(6):521-527.
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
现代装饰(2022年3期)2022-07-05 05:56:36
中老年保健(2021年10期)2021-11-30 09:34:06
皮肤病与性病(2021年3期)2021-07-30 08:07:40
云南医药(2021年3期)2021-07-21 05:41:06
今日农业(2020年24期)2020-12-15 16:16:00
四川文理学院学报(2020年5期)2020-02-12 02:05:58
兽医导刊(2016年12期)2016-05-17 03:51:50
少儿科学周刊·儿童版(2016年2期)2016-03-19 11:54:12
癌变·畸变·突变(2015年3期)2015-02-27 06:15:13
