
重金属胁迫对两种海洋饵料微藻的急性毒性效应研究*
2014-04-17 07:48于小娣王其翔张鑫鑫唐学玺
中国海洋大学学报(自然科学版) 2014年2期
于小娣,师 玥,刘 泳,周 斌,王其翔,张鑫鑫,唐学玺,王 悠**
(1.中国海洋大学海洋生命学院,山东 青岛266003;2.青岛市海泊河污水处理厂,山东 青岛266021;3.山东省海水养殖研究所,山东 青岛266002)
重金属是海洋三大污染物之一,其危害已受到人们广泛关注。据报道,2009年我国海域受重金属污染的面积继续增大,对海洋生态系统危害极大[1]。海洋微藻是海洋初级生产力的重要组成部分,也是海洋生态系统中食物链的起点。当水体受到重金属污染时,海洋微藻是首要的污染对象,并通过食物链富集、传递作用对食物链各营养级的生物产生毒害作用[2-3]。有关重金属对海洋微藻毒性效应的研究有许多报道,重金属胁迫会破坏微藻的叶绿体结构,使其变形,同时使细胞壁松散,细胞内液泡数量增多,体积增大[4];还能破坏海藻叶片细胞的膜结构和非膜结构,对其呼吸、代谢等重要生理过程产生影响[5]。
铜(Cu2+)、镉(Cd2+)和锌(Zn2+)是3种典型的污染重金属,其中Cu2+和Zn2+是微藻生长所必需的元素,痕量的Cu2+或Zn2+能维持微藻体内正常的生理代谢过程,但水体中过高浓度的重金属会破坏其膜系统及细胞器,影响微藻的光合作用和正常物质代谢过程。Cd2+的毒性很强,且具有高生物富集性,可造成生物体的急性、亚急性致死作用[6-8]。从目前的研究来看,重金属对饵料微藻的毒性效应研究相对集中于某一生物组织层次上,从亚细胞、细胞和种群等多生物组织层次系统上来分析重金属的毒性效应及损伤途径的研究较少[7]。
本研究以山东近岸常见的2种海洋饵料微藻:青岛大扁藻(Platymonashelgolandicavar.tsingtaoensis)和等鞭金藻8701(Isochrysisgalbana8701)为目标,在实验生态条件下研究3种重金属离子:Cu2+、Cd2+和Zn2+的急性毒性胁迫对其种群增长的影响,结合透射电镜技术(Transmission Electronic Microscope,TEM)分析重金属胁迫对微藻亚细胞结构的损伤,从不同生物组织层次上分析重金属的毒性效应。研究结果为研究重金属污染对微藻群落结构的变化影响提供数据。
1 材料与方法
1.1 实验材料
实验用2种常见的饵料微藻:绿藻门的青岛大扁藻(Platymonashelgolandicavar.tsingtaoensis)和金藻门的等鞭金藻8701(Isochrysisgalbana8701),均取自中国海洋大学生态毒理学实验室。
微藻培养用的海水均为取自青岛市鲁迅公园的自然海水,经沉淀、过滤(0.45μm 微孔滤膜),121.3℃灭菌20min后用于藻类的培养。微藻培养的三角瓶预先用1mol/L的HCL浸泡24h,再用蒸馏水冲洗干净后于121.3℃灭菌20min。培养过程中,将已知起始密度的目标微藻接种于经f/2培养液[9]加富的灭菌海水中,培养温度(19±1)℃,光照66μmol/(m2·s),光暗周期12h∶12h。培养过程中定时摇动以防止其附壁生长。
1.2 实验设计
CuSO4·5H2O(AR),ZnSO4·7H2O(AR),3CdSO4·8H2O(AR)分别用重蒸水配成相应的母液。在预实验基础上等间距设置5个浓度组(见表1)。

表1 急性毒性实验设计Table 1 Acute toxic effects design /mg·L-1
实验微藻的初始密度为1.0×105cells/mL,实验总体积为120mL,每个实验组设3个平行样(n=3),以不加入重金属污染物的一组为对照组。整个实验周期为120h,每24h取样1mL,先用Lugol碘液固定,再用血球计数板计数,根据计数结果计算微藻种群的密度、相对增长率、相对抑制率及48h-EC50和96h-EC50。
1.3 Cu2+胁迫下2种海洋微藻的透射电镜观察
取10mL Cu2+胁迫了96h的藻液,用0.22μm的滤膜过滤,收集滤膜上的藻细胞,用0.1mol/L磷酸钾缓冲液(pH=7.4)冲洗藻体,再过滤,重复2次。在室温下用5%的戊二醛固定4 8h后进行透射电镜观察(HITACHI-700TEM)。

图1 Cd2+胁迫对2种微藻种群增长及相对增长率的影响Fig.1 Acute toxic effect of Cd2+ on population growth and relative growth rates of two species of microalgae
实验中,Cu2+对等鞭金藻8701的胁迫浓度为0.2和0.6mg/L;Cu2+对青岛大扁藻的胁迫浓度为0.5和1.5mg/L。
1.4 数据分析
用图像处理软件Singmaplot 10.0作图。相对增长率计算公式如下:
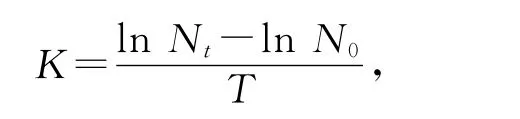
其中:Nt为培养t时刻的细胞密度;N0为起始细胞密度;T为培养时间(h)。
相对抑制率计算公式如下:

其中:Kc为对照组相对增长率;Ki为处理组相对增长率。
半数有效抑制浓度EC50利用统计软件Spss16.0中的概率单位-浓度对数回归法计算求得。
2 结果
2.1 3种重金属离子对2种微藻的毒性效应
在实验浓度范围内,青岛大扁藻的种群密度随着Cd2+胁迫时间的增加而不断增加,但相对增长率却随着时间的延长不断降低,48h后各处理组的相对增长率均明显低于对照组(Paired t-test,P<0.05)。96h时,青岛大扁藻(见图1a)对照组的相对增长率为16%,低浓度组(Cd2+=14mg/L)的相对增长率为11%左右,而高浓度组(Cd2+=22mg/L)的相对增长率为4%,差异显著。等鞭金藻8701的种群密度及相对增长率的变化趋势同青岛大扁藻类似(见图1b),与对照组相比(相对增长率为32%),96h时低浓度组(Cd2+=0.3mg/L)的相对增长率为28%(P<0.05),而高浓度组(Cd2+=3.1mg/L)生长几乎停止,相对增长率只有1%,差异极显著(P<0.01)。青岛大扁藻对Cd2+的敏感性要低于等鞭金藻8701。
Zn2+对2种微藻生长的影响情况与Cd2+基本相同:与对照组相比,低浓度(Zn2+=8mg/L)的Zn2+胁迫对青岛大扁藻的生长抑制不明显(Paired t-test,P>0.05),在24h内还有一定的促进作用;96h时低浓度组的相对增长率为12%,对照组的相对增长率为12.6%。高浓度组(Zn2+=38mg/L)的Zn2+胁迫对微藻生长具有抑制作用,96h时与对照组相比达到极显著水平(Paired t-test,P<0.01),此时微藻种群的相对增长率只有2.5%。Zn2+对等鞭金藻8701的胁迫表现出类似的作用趋势,与对照组相比,在24h时低、中浓度组(3.5~5.0mg/L)的细胞密度比0h略有增加,但相对增长率与对照组相比有所降低,且呈现出明显的浓度依稳性,即浓度越高抑制作用越明显。至96h时,高浓度组(5,6.5,8mg/L)的抑制作用与对照组相比达到极显著程度(P<0.01)(见图2)。

图2 Zn2+胁迫对2种微藻种群增长及相对增长率的影响Fig.2 Acute toxic effect of Zn2+ on population growth and relative growth rates of two species of microalgae
Cu2+对2种微藻种群增长的抑制作用与Cd2+和Zn2+基本一致,但Cu2+对2种微藻的毒性作用更大。如图3所示,各处理组及对照组中的细胞密度均随胁迫时间的延长而不断增加,但处理组中的细胞密度始终小于处理组,并呈现出一定的剂量-效应关系,即胁迫浓度越高,种群密度越低;相对增长率的变化与种群密度变化趋势一致,至96h时,低浓度组(Cu2+=0.2mg/L)的相对增长率为31%,与对照组相比差异性不显著(P>0.05),但高浓度组的相对增长率为7%左右,与对照组相比差异极显著(P<0.01)。
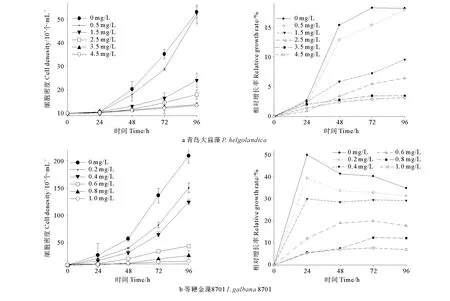
图3 Cu2+胁迫对2种微藻种群增长及相对增长率的影响Fig.3 Acute toxic effect of Cu2+ on population growth and relative growth rates of two species of microalgae
根据上述实验结果,计算了3种重金属对2种微藻胁迫的48h-EC50和96h-EC50见(见表2)。
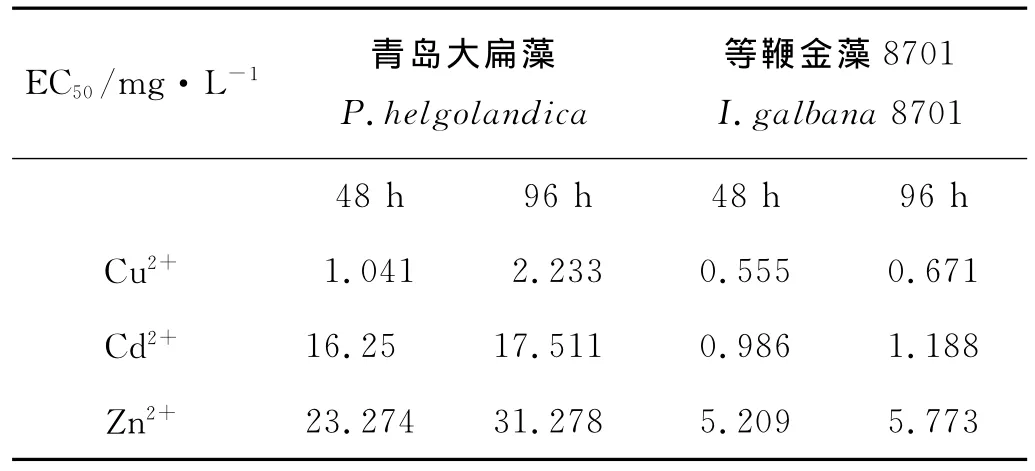
表2 3种重金属对2种微藻胁迫的48h-EC50和96h-EC50Table 2 48h-EC50and 96h-EC50values when exposed to different concentrations of heavy metals’stress
3种重金属离子胁迫下等鞭金藻8701的48h-EC50和96h-EC50都低于青岛大扁藻,而3种金属对于2种藻的48h-EC50和96h-EC50的大小比较为 Cu2+<Cd2+<Zn2+(见表2)。等鞭金藻8701对于3种金属胁迫的敏感性大于青岛大扁藻,同时3种重金属离子的毒性大小表现为Cu2+>Cd2+>Zn2+。2种微藻的48h-EC50都低于其96h-EC50,这说明随着胁迫时间的延长,3种重金属对2种藻生长的抑制作用是减弱的。
2.2 Cu2+胁迫条件下微藻的超微结构的变化
2.2.1 等鞭金藻8701的超微结构的变化 以毒性最强的Cu2+为目标,研究了重金属胁迫对青岛大扁藻亚细胞结构的影响,结果发现:对照组的金藻细胞形态完整,呈椭圆形,长约5μm,宽约4μm,最外层有鳞片,没有细胞壁,内里2个大的片状色素体(见图4A)紧贴细胞膜,并环藻体细胞分布,色素体中间有一个大的白糖体(见图4B)。细胞核完整,呈椭圆形于细胞中间,有核膜包被,核仁清晰可见,有多个线粒体呈椭圆形或长条状,没有大液泡(见图4B)。0.2mg/L的低浓度处理组中,藻细胞鳞片部分脱落,细胞质内空泡数量增多,液泡增大,细胞核与高尔基体出现轻微水肿,叶绿体片层结构少量断裂,有细胞分泌物产生,依旧可见多个线粒体,且形态完整,各细胞器膜结构未发生大的变化(见图4C)。0.6mg/L的高浓度组中,藻细胞鳞片脱落现象严重,内部细胞质空泡化更严重,大液泡大量产生,细胞核轻度变形,核膜水肿程度增加,但结构仍然完整,细胞分泌物增多甚至连成一片,叶绿体几乎完全断裂,且排布混乱,但其中的淀粉粒结构完整,并未太多受损,且线粒体结构仍然完整(见图4D)。TEM检测结果显示,Cu2+对等鞭金藻8701的光合作用供能系统损伤较大,但对线粒体和细胞核影响较小。中上部有一个大的淀粉(蛋白质)核,未观察到脂肪粒(见图5A)。藻细胞中央有一个细胞核,有核膜包被,中央部位靠近细胞膜的位置生有眼点(见图5B)。当受到Cu2+时(见图5C-F),藻细胞外部形态变化不大,并未出现变形或破损的现象,但细胞壁增厚(见图5D),且随着胁迫浓度的增大,细胞内部淀粉粒的数量明显减少,虽然大的淀粉核依旧可见但周围的淀粉粒明显变小且排列紊乱(见图5E),脂肪粒的数量明显增多,个体增大,说明其储能机制出现问题,脂肪代谢产生阻碍。另外,随着胁迫浓度的增加其细胞核虽有水肿但形态依然完整(见图5D)。
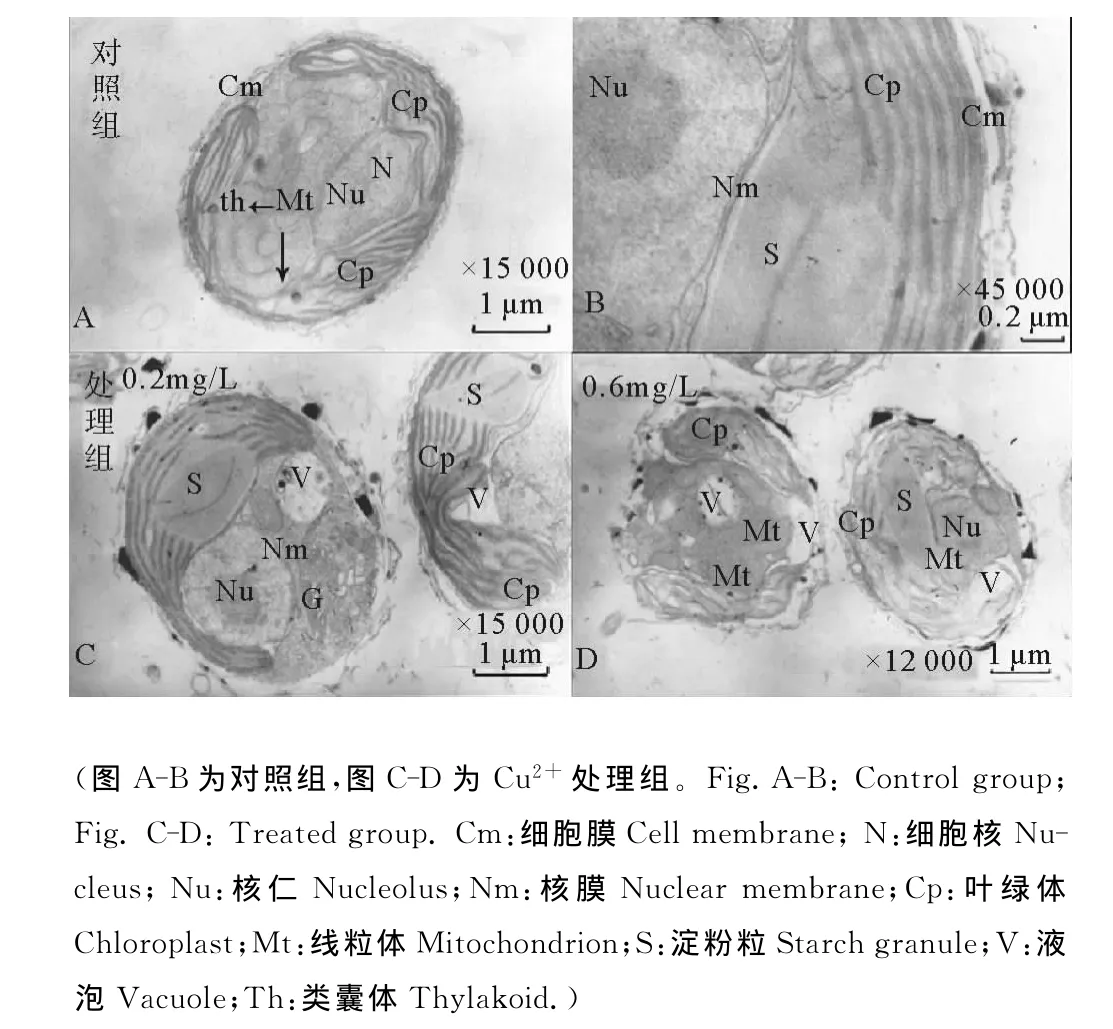
图4 Cu2+胁迫前后等鞭金藻8701超微结构变化Fig.4 Changes of ultrastructure of I.galbana8701when exposed to Cu2+ stress under TEM
2.2.2 青岛大扁藻的超微结构的变化 对照组中的青岛大扁藻藻体细胞呈椭圆形,且细胞外壁染色较深,这是它含有纤维素的细胞壁结构,细胞中可以看到有多个棒状的淀粉粒分布在藻细胞的叶绿体中,在细胞

图5 Cu2+胁迫前后青岛大扁藻超微结构变化Fig.5 Changes of ultrastructure of P.helgolandica when exposed to Cu2+ stress under TEM
3 讨论
当生物体受到污染物刺激时,会出现一个先抑制后补偿的过程,但是随着生物体对污染物的适应,补偿行为会逐渐超过抑制行为,最终导致一个净刺激效应,这个效应被称为hormesis效应,即兴奋效应[10]。在实验中,Zn2+胁迫对青岛大扁藻的生长表现出低浓度刺激、高浓度抑制的现象。高浓度的3种重金属则对微藻均表现出显著的抑制作用,其中Cu2+的毒性效应最强,Cd2+次之,Zn2+最弱。同时,通过2种藻48h-EC50和96h-EC50的比较发现,等鞭金藻8701对3种重金属的敏感性比青岛大扁藻高。从以往的研究可知,藻类细胞表面所包含的与重金属有强亲和性的官能团的数量和密度是决定微藻对重金属的敏感性差异的主要原因[11]。因此很可能是2种微藻与Cu2+的亲和性最大,Cd2+和Zn2+次之,或是它们所具有的亲和Cu2+的官能团最多。
另一方面,微藻细胞有两大供能系统:光合系统和呼吸系统。其中线粒体是有氧呼吸产能的主要场所,色素体如叶绿体等是主要的光合作用场所。重金属对海洋微藻发生胁迫时,微藻首先会通过自身防御系统抵制重金属的损伤,主要通过自身细胞壁所含官能团或分泌胞外物质与金属离子发生螯合,已进入胞内的会与液泡释放物质或细胞质中所含物质络合形成沉淀,再排出胞外[12]。因此,重金属对海洋微藻的损伤首先主要体现在对藻细胞及细胞器内膜系统的损伤,但当两大供能系统的主要发生场所受损后,藻细胞代谢必然会产生异常。另外,这也说明,不同藻种在新陈代谢过程中,主动排除细胞内和表面重金属的能力也会影响到自身受重金属胁迫损伤的程度[13]。
Cu2+是生物体内多种酶的组成成分,并且可以作为辅助因子参与植物的代谢过程,包括呼吸作用、光合作用和藻类的增殖等[14]。因此,选取Cu2+作为胁迫因子对2种藻的亚显微结构进行观察研究是很有必要的,从本文的实验结果来看,2种藻对Cu2+胁迫都最为敏感,并且TEM检测结果显示,当Cu2+浓度增加时,2种藻的光合系统或代谢过程都有不同程度的损伤。当藻类与金属离子开始接触时,藻首先表现为对金属的表面吸附,低浓度时这种吸附不足以威胁到藻细胞,当金属离子集聚到一定浓度时就会触发藻细胞的自身防御机制。有些藻类可以通过细胞壁上带负电荷的配位基团的结构将其重金属离子沉淀于表面,阻止其进入,实验中青岛大扁藻(见图5)便含有细胞壁结构[15],这可能是青岛大扁藻对3种重金属的敏感性小于等鞭金藻8701的主要原因[16]。没有细胞壁的藻体,可以通过分泌胞外物质将重金属排出体外并阻止其进入,在等鞭金藻8701(见图4)的TEM检测结果中可以看到其藻体周围有染色较深的胞外物质,随着胁迫浓度的增大,这些胞外物质有数量增多、体积增大的趋势。另外,藻细胞内的液泡含有多种物质,常能与重金属发生络合,降低毒性,当藻细胞受胁迫时,内部液泡会释放这些物质,在本实验TEM监测结果中也可以明显看到,内部空泡化增强,液泡数量增多、体积增大的现象。当胁迫浓度不断增大至超过藻细胞自身承受能力时,就会破坏这些防御机制,使膜结构渗透压改变,重金属大量进入,内部膜结构遭到破坏,两大供能系统受损,代谢紊乱,各细胞贮能结构减少,细胞生长受到抑制,甚至停止生长[17-18]。
[1]国家海洋局.2009年中国海洋环境质量公报[J].海洋开发与管理,2010(4):24-57.
[2]Haritonidis S,Malea P.Bioaccumulation of metals by the green algaUlvarigidafrom Thermaikos Gulf,Greece[J].Environmental Pollution,1999,89(3):319-327.
[3]Edding M,Tala F.Copper Transfer and Influence on a Marine Food Chain[J].Bulletin of Environmental Contamination and Toxicology,1996,57(4):617-624.
[4]赵艳芳,俞志明,宋秀贤,等.营养盐对长江口2种主要赤潮原因藻光合色素和光合作用影响的比较研究[J].环境科学,2009,30(3):700-706.
[5]施国新,杜开和,解凯彬,等.汞、镉污染对黑藻叶细胞伤害的超微结构研究[J].植物学报,2000,42(4):373-378.
[6]周启星,孔繁翔,朱琳.生态毒理学[M].北京:科学出版社,2004:306-348.
[7]周汝伦,孙爱淑.金藻8701的分离、培养和应用初报[J].海洋湖沼通报,1990,1:34-40.
[8]周宏,项斯端.重金属铜、锌、铅、镉对小形月牙藻生长及亚显微结构的影响 [J].杭州大学学报(自然科学版),1998,25(2):85-92.
[9]Guillard R R L.Culture of phytoplankton for feeding marine invertebrates[M]//Smith W L,Chanley M H.Culture of Marine Animals.New York:Plenum Press,1975:26-60.
[10]Mark P M.Hormesis Defined[J].Ageing Res Rev,2008,7(1):1-7.
[11]Melchor G D.The role of phytoplankton cells on the control of heavy metal concentration in seawater[J].Mar Chem,1995,48:215-236.
[12]Baker A J M.Metal Tolerance [J].New Phytologist,1987,106:93-111.
[13]李英敏,杨海波,吕福荣,等.叉鞭金藻对微量锌镉的吸附效应研究[J].环境污染与防治,2004,26(5):396-398.
[14]Satoha A,Vudikariab L Q.Evaluation of the sensitivity of marine microalgal strainsto the heavy metals,Cu,As,Sb,Pb and Cd[J].Environment International,2005,31(5):713-722.
[15]刘春凤,吴雪,高悦.两种等鞭金藻外部形态与超微结构的观察比较[J].大连水产学院学报,2008,23(4):273-277.
[16]Garrido J,Gaspar A,Garrido E M.Alkyl esters of hydroxycinnamic acids with improved antioxidant activity and lipophilicity protect PC12cells against oxidative stress[J].Biochimie,2012,94:961-967.
[17]苏秀榕,刘照彬,迟庆宏.从超微结构变化研究Cu2+,Zn2+,Se4+,Cd2+对三角褐指藻的毒性效应[J].辽宁师范大学学报,2000,23(1):68-72.
[18]梁英,王帅.重金属对微藻胁迫的研究现状及前景[J].海洋湖沼通报,2009,11(4):72-82.
猜你喜欢
中国岩溶(2022年3期)2022-11-30
今日农业(2022年15期)2022-09-20
古今农业(2022年1期)2022-05-05
河北金融年鉴(2021年0期)2021-08-25
知识就是力量(2021年6期)2021-07-09
湖南电力(2021年1期)2021-04-13
河北金融年鉴(2020年0期)2021-01-21
环球时报(2017-01-26)2017-01-26
海外星云 (2014年21期)2015-01-14
食品工业科技(2014年23期)2014-03-11
