
纹状体神经元可塑性与帕金森病的运动防治研究进展
2014-04-15 02:43:52陈巍乔德才刘晓莉
中国运动医学杂志 2014年7期
陈巍 乔德才 刘晓莉
1 北京师范大学体育与运动学院(北京100875)
2 河北科技师范学院体育系
帕金森病(Parkinson’s disease,PD)是继阿尔茨海默病之后的第二大神经退行性病变。 我国55岁以上人群中PD发病人数多达170余万人, 且仍以每年10万的速度递增, 该病对健康的危害已引起社会的高度关注[1]。 PD主要病理改变是黑质致密部多巴胺(dopamine,DA) 神经元渐进性变性,DA神经递质缺失,从而引起基底神经节功能失调[2]。 DA替代治疗及手术对PD患者有一定效果,但随着病情加重,这些方法的有效性逐渐降低,副作用也逐渐显现[2,3],其中最为突出的是出现运动障碍,严重影响了PD患者的日常生活。 有研究证实,PD的发生与纹状体内中等多棘神经元(medium spiny neurons,MSNs)功能改变有密切关系[4]。 近年来的流行病学调查结果表明,身体活动水平与PD的发病之间存在一定关联[5,6]。 临床资料显示,适当的身体活动能显著改善PD患者的身体姿势、步态、平衡等行为功能,其机制可能与运动增加了DA的合成与分泌、促进了纹状体神经元结构与功能可塑性,从而改善PD患者基底神经节的功能紊乱有关[7]。
1 基底神经节在运动调控中的作用
躯体运动是在大脑皮层和基底神经节的调控下实现的。 从功能上可将基底神经节划分为纹状体(Str)、苍白球外侧部(GPe)、黑质致密部(SNc)、丘脑底核(STN)、黑质网状部/苍白球内侧部复合体(SNr/GPi)等5个核团[8]。 基底神经节接受来自大脑皮层和丘脑的神经投射,经直接通路(direct pathway)和间接通路(indirect pathway)调节与整合后,再经丘脑腹前核(VA)和腹外侧核(VL)返回皮层,构成了皮质-基底节-丘脑-皮质神经环路[2,8]。其中,直接通路是指从大脑皮层的广泛区域到纹状体, 再由纹状体发出纤维, 经苍白球内侧部接替后到达丘脑腹前核和腹外侧核, 最后返回大脑皮层运动前区和前额叶的通路; 而间接通路是指在上述直接通路中的纹状体与苍白球内侧部之间插入苍白球外侧部和丘脑底核两个中间接替过程的通路[9]。研究人员早已明确基底神经节发出下行纤维到达脊髓,参与锥体外系,调控骨骼肌活动。 其中,黑质致密部DA能神经元发出黑质-纹状体投射纤维调控纹状体的两个主要传出通路,一条作用于纹状体γ-氨基丁酸 (γ-amino butyric acid,GABA)/P物质能神经元上的多巴胺Ⅰ型受体(D1DR),促进直接通路的传递;另一条是DA释放后作用于GABA/ENK能神经元上的多巴胺Ⅱ型受体(D2DR),抑制间接通路的传递。 黑质-纹状体通路的DA能神经元在纹状体内释放DA总的效应是减少基底神经节对丘脑的抑制,促进丘脑-皮质投射神经元的活动[8,9]。 因此,黑质被认为是调节躯体运动的“开关”,而纹状体被认为是基底神经节神经回路的中央加工区, 它们均直接或间接参与随意运动的程序编制与执行, 在实现躯体运动参数调控方面发挥重要作用。
2 PD与基底神经节功能紊乱
PD是一种中枢神经系统进行性退变引发的运动障碍疾病, 主要临床表现为静止性震颤、 运动徐缓、肌强直和姿势步态异常等[2]。 PD的发病机制尚未明确,大多学者认为可能与环境、免疫学异常、线粒体功能障碍、氧化应激、年龄老化、细胞凋亡等因素有关[10]。 目前已经明确,黑质致密部DA能神经元变性缺失及纹状体DA水平降低是诱导PD的起始事件[9,10]。研究显示,PD动物模型表现出纹状体神经元自发放电频率增高[11]以及丘脑底核神经元高频放电与爆发式放电增多等异常现象[12]。这可能与黑质-纹状体DA能系统活性减弱, 而皮层-纹状体谷氨酸(glutamate,Glu)能系统活性增强有关[10]。 由此提出了基底神经节内神经递质比例失衡引发的直接通路兴奋性降低,而间接通路兴奋性增强,最终抑制丘脑-皮质投射神经元的活动可能是引发PD患者行为功能障碍的假设(图1)。
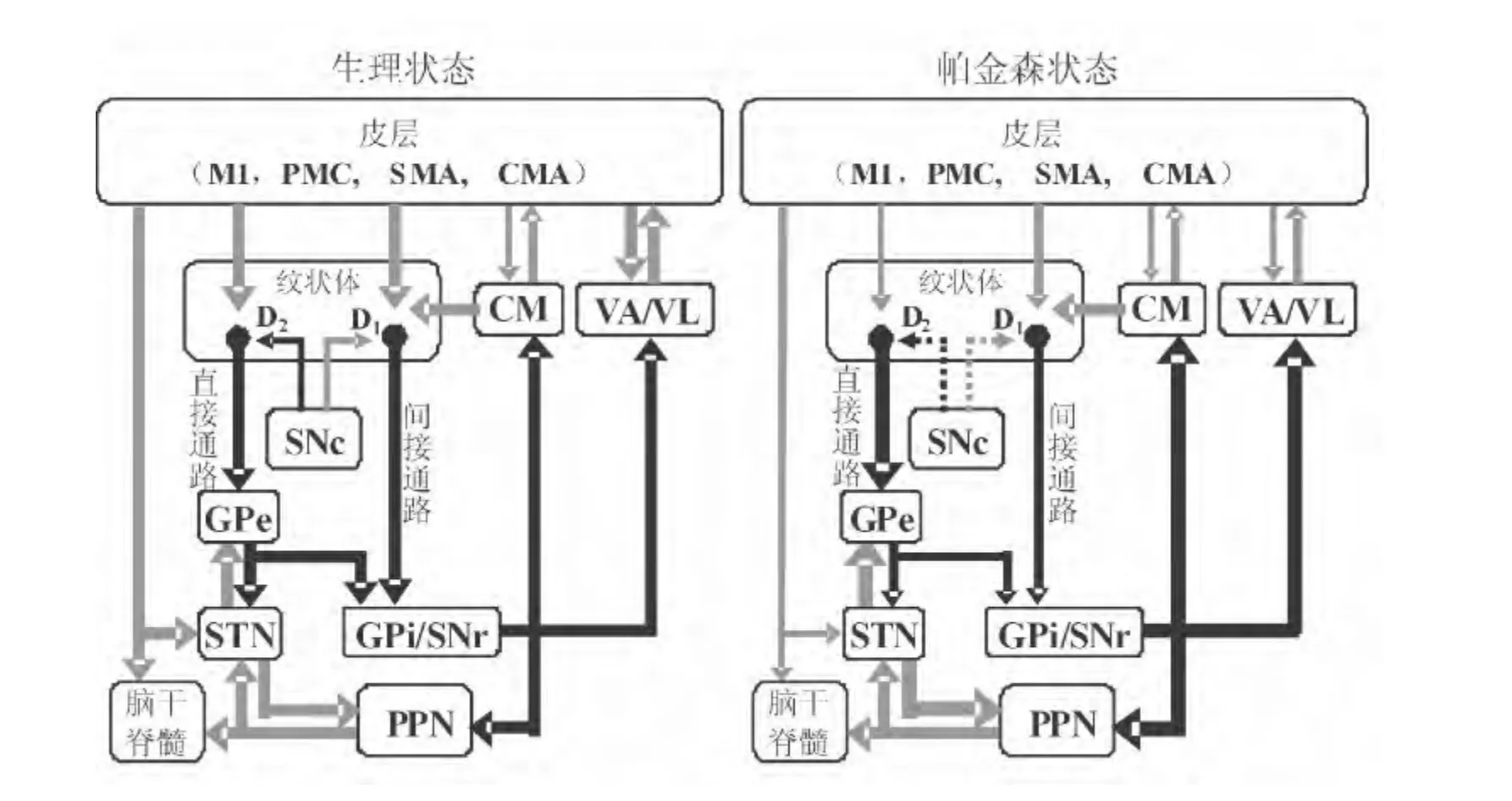
图1 PD基底神经节-丘脑-皮层运动环路的活性变化模式图[10]
3 PD与纹状体中等多棘神经元可塑性
3.1 纹状体MSNs形态与突触分布
纹状体内的神经元约95%属于MSNs, 其树突及其分支上存在大量棘状突起,即树突棘,它是神经元之间形成突触的重要部位[13]。 电子显微镜观察显示,MSNs可同时接受黑质-纹状体和皮层-纹状体两条通路的神经纤维投射,源自皮层的Glu能突触位于树突棘头部,而源自黑质致密部的DA能突触则位于树突棘颈部[14]。所以在解剖学上,纹状体MSNs是调节直接通路和间接通路活动的重要位点,MSNs的形态与结构可塑性在运动行为功能调节方面具有重要作用。动物实验显示,去DA能神经支配后,纹状体MSNs兴奋性显著增强[15]。 最近研究还发现,纹状体内活化程度较高的Glu能突触的MSNs树突棘可优先接收DA能神经元轴突的投射, 进一步证实了DA对Glu能突触传递的调节作用[16]。
3.2 PD与MSNs形态及突触功能
对动物模型的研究均发现,纹状体MSNs树突棘密度与树突长度明显降低[17,18]。 在PD发病早期,通过增加纹状体D2DR表达可代偿DA水平的下降,但随着疾病发展,MSNs树突棘数量显著减少会限制这种代偿机制,这在一定程度上解释了DA替代治疗在PD发展晚期的局限性[14]。也有研究发现,黑质致密部DA能神经元丢失后,纹状体MSNs内不对称突触总体数量降低, 但不对称突触中穿通型突触的比例却成倍增加[19]。这提示失去DA能神经支配后,来自大脑皮层或丘脑的Glu能纤维活性增强;而实施皮质剥除术又可抑制由于DA缺失诱导的纹状体MSNs树突棘数量丢失,表明纹状体MSNs树突棘脱落与皮层-纹状体Glu活性增强有直接联系。 目前认为,PD的发生与纹状体DA受体波动性刺激引起的下游突触后神经元功能改变导致的直接通路和间接通路失衡有关。 MSNs突触结构与功能可塑性的变化与PD的发展存在密切联系[20]。
3.3 MSNs树突棘形态与功能重塑
目前已有证据表明, 中枢神经系统内树突棘形态与数量的改变与Glu介导的胞内Ca2+水平增加有关。 胞浆内Ca2+水平增加主要通过3条途径实现,即Glu受体门控通道、电压门控Ca2+通道(VSCCs)及内质网的Ca2+释放[21]。 其中,N-甲基-D-门冬氨酸(NMDA)受体是位于突触后膜上的配体门控Ca2+通道,其包含两个亚单位家族,即NMDAR1与NMDAR2,它们的数量与组成可直接影响NMDA受体的功能。 研究发现,纹状体内NMDAR1分布与数量与D2DR受体分布的密集区相一致[22];NMDAR1与D2DR之间的平衡受到破坏将引起皮层-纹状体MSNs突触结构和功能重塑[21,22]。 VSCCs拮抗剂可以抑制PD动物由于DA缺失诱导的树突棘密度下降;而L-型Ca2+通道(CaV1.3)基因缺失的PD动物纹状体MSNs未出现树突棘脱落现象[20],表明CaV1.3可能也参与PD纹状体MSNs突触可塑性的调节。 当胞内Ca2+水平升高时,即可引发钙结合蛋白(CaM)构象改变,进而促进与Ca2+依赖型蛋白激酶II(CaMKII)结合,增加其286位苏氨酸磷酸化水平。 CaMKⅡ激活后与NMDA受体亲和力增加,却与富含突触后致密物(postsynaptic densitied,PSD)的亚细胞结构的结合水平下降[20]。 由于CaMKⅡ占PSD组分蛋白总量的20%~30%,同时PSD又是树突棘的重要结构基础, 因此,CaMKⅡ过度激活可能会影响到树突棘的稳定性。 这提示胞内Ca2+水平升高诱导MSNs树突棘形态与数量变化的机制与CaV1.3 及CaMKII的活化程度密切相关(图2)。
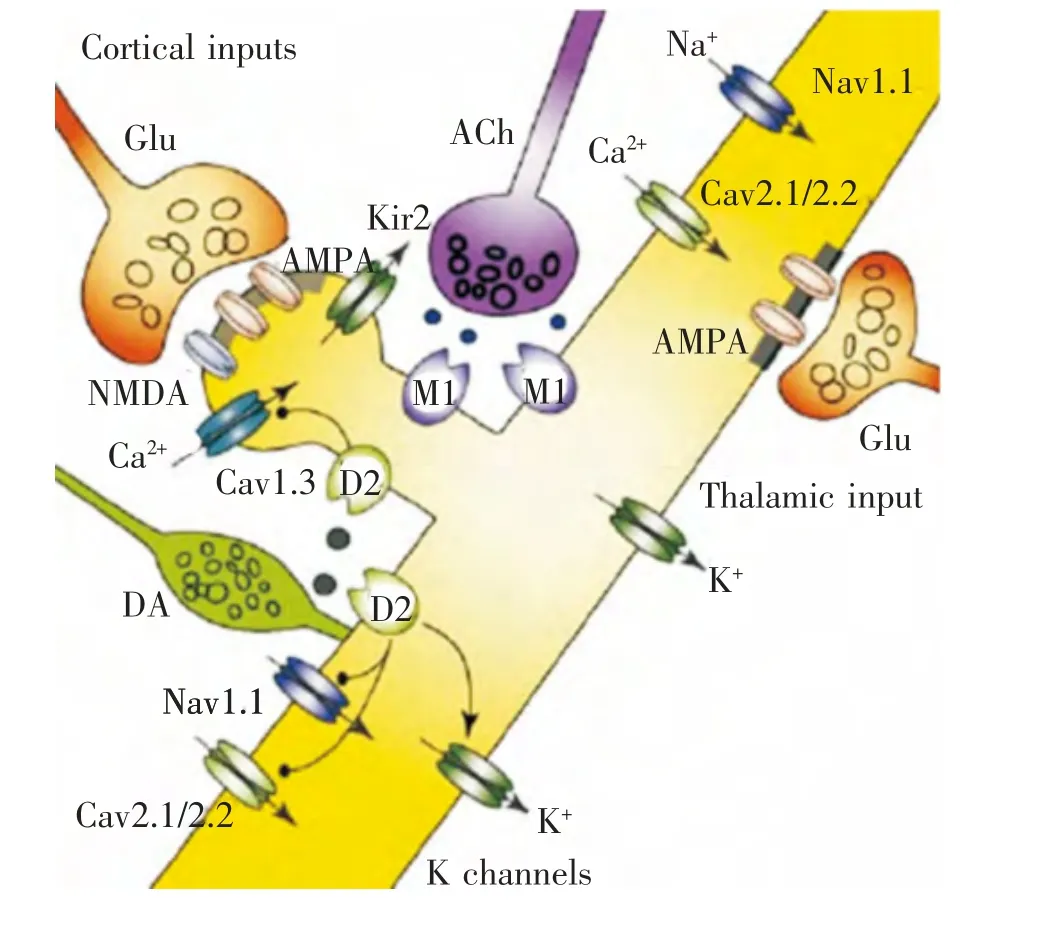
图2 纹状体MSNs树突棘DA突触传递对Ca2+信号调节的影响[20]
4 PD的运动干预及相关机制研究
4.1 PD临床研究与运动干预
在相当长的一段时间内, 由于人们并不认为运动可以对PD产生预期的效果,甚至还认为可能会使PD病情恶化,因此最初运动疗法并没有被推崇为PD患者的康复策略。 之后的流行病学研究发现,PD发病风险与身体活动水平之间存在显著负相关, 由此引发了研究者们对运动影响PD患者脑功能及行为能力等的关注[1,6]。 PD患者进行跑台训练后,与接受标准物理治疗的患者比较,疾病严重程度评分降低,步态与平衡功能均有较为明显的改善[7]。 Evan等[23,24]的研究也发现,接受运动疗法的PD患者,身体活动水平较未接受运动治疗的患者显著提高。 Paul等[25,26]的研究提出,PD患者应将运动干预重点放在有效增强神经-骨骼肌系统的功能上, 以缓解患者肌力衰退, 并建议将力量训练作为防治PD的有效手段之一。Kleim等[27]认为,较小的运动负荷不足以对中枢神经系统产生有效刺激, 运动强度与持续时间对神经系统可塑性的影响至关重要。目前,临床上已将运动干预作为治疗PD患者的一种新的治疗手段,通过力量训练、平衡训练、拉伸训练等方法改善患者的身体姿势、 步态、 日常活动能力以及神经肌肉系统的症状,以全面提高PD患者生活质量[6,7,25,26,28]。
4.2 运动干预防治PD的相关生物学机制
1-甲基-4-苯基-1,2,3,6-四氢吡啶(MPTP)和6羟基多巴(6-OHDA)诱导的PD模型能够以与环境毒素相似的作用机制选择性地破坏黑质DA能神经元[3,5,6],使动物在病理及行为等方面出现与PD患者相似的特征,且具有建模成功率较高、可操作性强、利于控制损伤程度和评价治疗效果等优点, 被广泛运用于PD发病机制的研究中[29]。 目前,有关运动对PD防治作用的生物学机制研究主要关注以下几个方面:
4.2.1 运动调节黑质-纹状体DA能神经传递
有研究显示预运动可显著提高PD动物黑质DA能神经元存活率,并明显改善PD动物的行为障碍,表明运动对DA能神经元产生保护效应[30,31]。 Yoon等[32]发现大鼠注射6-OHDA 24小时后,进行30 min/d、连续2周的跑台运动,结果显示,黑质-纹状体通路DA能神经元的损伤程度明显降低。 Tajiri等[33]的研究也得到同样的结果。 Gerecke等[30]的研究显示,小鼠在注射MPTP前预先接受3个月的自主跑轮训练,纹状体DA水平显著升高,但黑质DA能神经元数量未见显著变化,提示预运动可促进DA的合成。 但也有研究发现,在注射MPTP 4~5天后进行数周跑台训练的小鼠行为功能明显改善, 纹状体DA水平却未见显著变化;用快速扫描循环伏安法检测显示DA突触间隙的衰减程度降低[34]。 由此可见,PD状态下,运动除增加DA合成外, 还可能通过促进DA能突触传递来调节黑质-纹状体神经通路的功能。利用反义寡核苷酸阻断DAT后,小鼠对MPTP毒素反应性减弱;DAT基因完全敲除的小鼠,MPTP对其黑质DA能神经元的毒作用消失[35]。 这提示DAT参与DA能神经元对MPTP摄取的调节。 Feisher等[36,37]的研究发现,MPTP小鼠进行运动训练后,其黑质DA能神经元数量及纹状体DA水平均未改变, 但其行为功能得到改善; 同时发现纹状体DAT表达下降而D2DR表达增加。 Marta等[38]的研究也发现,跑台训练可以显著上调MPTP小鼠纹状体D2DR表达; 采用18F-fallypride作为D2DR受体显像剂进行观察发现, 运动可增加MPTP小鼠D2DR结合潜能,进一步证明了运动具有促进黑质-纹状体DA能神经传递的作用,这也在一定程度上解释了PD动物进行运动训练后行为功能改善与DA总量变化并非一致的现象。
4.2.2 运动调节皮层-纹状体Glu能神经传递
生理状况下,纹状体神经元活动受黑质DA 能神经元与大脑皮质Glu能神经元共同调节。 PD状态下,伴随着黑质DA能神经元的丢失,皮层-纹状体Glu能神经传递明显增强,纹状体Glu水平显著升高,这在PD发展进程中产生重要影响[8,10]。 VanLeeuwen等[39]的研究显示,小鼠在注射MPTP 5天后,接受为期4周的高强度跑台训练, 纹状体GluR2亚基表达显著增加,MSNs兴奋性突触后电位波幅下降,同时,其运动功能得到改善。此外,还有研究发现,经MPTP处理的小鼠纹状体AMPA/NMDA比率显著升高, 而高强度运动后此比率明显下降[40]。 这提示运动改善PD动物行为功能的机制可能与皮质-纹状体Glu能神经传递活性降低有关。
4.2.3 运动可能促进PD动物纹状体MSNs形态与功能重塑
已有研究证实,PD模型动物行为功能改善与纹状体MSNs形态与功能重塑并存[41]。 虽然目前尚缺乏运动对PD患者及动物纹状体MSNs可塑性影响的直接证据,但Takamatsu等[42]的研究发现,脑缺血可诱导大鼠纹状体MSNs树突棘密度下降,胶原酶损毁纹状体后的4~14天进行跑台训练,纹状体MSNs树突长度及树突棘形态均有改变。 本实验室的前期研究[43]中,大鼠注射6-OHDA 24小时后,进行11 m/min、30 min/d、5 day/wk、连续4周的跑台运动,结果显示,PD大鼠纹状体MSNs高频放电活动降低,爆发式放电活动减少,且大鼠行为功能明显改善。 上述研究表明,MSNs功能可塑性可能参与了运动对PD大鼠行为功能改善的过程。
5 小结
PD的发生与黑质致密部DA能神经元渐进性缺失及纹状体DA水平显著下降导致的基底神经节调节功能紊乱密切相关。纹状体MSNs可接受黑质DA能神经元与皮层Glu能神经元的双重投射,MSNs形态与功能的改变与PD的发病存在密切联系。 运动通过调节黑质-纹状体DA能神经传递与皮层-纹状体Glu能神经传递改善PD症状的机制可能与纹状体MSNs形态与功能重塑有关。 探索运动对PD患者纹状体MSNs可塑性的影响及机制可能是今后PD防治研究的新关注点。
[1] Shin YC,Sheng TT. The epidemiology of Parkinson’s disease. Tzu Chi Med J,2010,22(2):73-81.
[2] Claire LT,Rebecca S,Smitaa PM,et al. Systematic review of levodopa dose equivalency reporting in Parkinson's disease. Mov Disord,2010,25(15):2649-2653.
[3] Follett KA,Weaver FM,Stern M,et al. Pallidal versus subthalamic deep-brain stimulation for Parkinson’s disease. New Engl J Med,2010,362(22):2077-2091.
[4] Blanchet PJ,Calon F,Morissette M,et al. Relevance of the MPTP primate model in the study of dyskinesia priming mechanisms. Parkinsonism Relat Disord,2004,10 (5):297-304.
[5] Xu Q,Park Y,Huang X,et al. Physical activities and future risk of Parkinson disease. Neurology,2010,75(4):341-348.
[6] Dibble LE,Hale TF,Marcus RL,et al. High intensity eccentric resistance training decreases bradykinesia and improves quality of life in persons with Parkinson’s disease:a preliminary study. Parkinsonism Relat Disord,2009,15(10):752-757.
[7] Herman T,Giladi N,Gruendlinger L,et al. Six weeks of intensive treadmill training improves gait and quality of life in patients with Parkinson’s disease:a pilot study.Arch Phys Med Rehabil,2007,88(9):1154-1158.
[8] Fabio B,Giuseppe N,Cristina T,et al. Functional changes of the basal ganglia circuitry in Parkinson's disease. Prog Neurobiol,2000,62(1):63-68.
[9] Robert ST,Michel D. Basal ganglia contributions to motor control:a vigorous tutor. Curr Opi Neurobiol,2010,20(6):704-716.
[10] Galvan A,Wichmann T. Pathophysiology of parkinsonism.Clin Neurophysiol,2008,119:1459-1474.
[11] 管强,曹学兵,孙圣刚,等. 帕金森病及异动症大鼠模型纹状体神经元的电生理变化的研究. 卒中与神经疾病,2005,12(6):323-326.
[12] 李文娟,席悦,张巧俊,等. 大鼠丘脑背内侧核神经元在帕金森病状态下电活动的变化. 西安交通大学学报 (医学版),2013,34(5):614-619.
[13] Zaja-Milatovic S,Milatovic D,Schantz AM,et al. Dendritic degeneration in neostriatal medium spiny neurons in Parkinson disease. Neurology,2005,64:545-547.
[14] Ariel YD,Roger JC,Danny JW. Striatal plasticity and medium spiny neuron dendritic remodeling in parkinsonism. PParkinsonism Relat Disord,2007,13:S251-S258.
[15] Bamford NS,Zhang H,Schmitz Y,et al. Heterosynaptic dopamine neurotransmission selects sets of corticostriatal terminals. Neuron,2004,42:653-663.
[16] Changlei X,Junpeng Z,Yujun L,et al. Dopaminergic axons preferentially innervate dendritic spines with hyperactive glutamatergic synapses in the rat striatum. Brain Res,2012,1486:92-102.
[17] Neely MD,Schmidt DE,Deutch AY. Cortical regulation of dopamine depletion-induced dendritic spine loss in striatal medium spiny neurons. Neuroscience,2007,149:457-464.
[18] Stephens B,Mueller AJ,Shering AF,et al. Evidence of a breakdown of corticostriatal connections in Parkinson’s disease. Neuroscience,2005,132:741-754.
[19] 曹学兵,孙圣刚,王岚,等. 左旋多巴诱发异动症大鼠皮质纹状体突触超微结构与功能的变化. 中华神经科杂志,2004,27(2):126-132.
[20] Day M,Wang Z,Ding J,et al. Selective elimination of glutamatergic synapses on striatopallidal neurons in Parkinson disease models. Nat Neurosci,2006,9:251-259.
[21] Thomas GO,Andrew M. Calcium regulation of actin dynamics in dendritic spines. Cell Calcium,2005,37:477-482.
[22] Ng J,Rashid AJ,So CH,et al. Activation of calcium/calmodulin-dependent protein kinase IIα in the striatum by the heteromeric D1-D2 dopamine receptor complex.Neuroscience,2010,165(2):535-541.
[23] Thacker EL,Chen H,Patel AV,et al. Recreational physical activity and risk of Parkinson's disease. Mov Disord,2008,23(1):69-74.
[24] Fuzhong L,Peter H,Kathleen F,et al. Tai Chi and postural stability in patients with Parkinson's Disease. New Engl J Med,2012,366(6):511-519.
[25] Paul SS,Canning CG,Sherrington C,et al. Reduced muscle strength is the major determinant of reduced leg muscle power in Parkinson’s disease. Parkinsonism Relat Disord,2012,18(8):974-977.
[26] Chris JH,Thomas AB,Chris P,et al. Progressive resistance training improves gait initiation in individuals with Parkinson’s disease. Gait Posture,2012,35(4):669-673.
[27] Kleim JA,Jones TA. Principles of experience dependent neuralplasticity:implications for rehabilitation after brain damage. J Speech Lang Hear Res,2008,51 (1):S225-S239.
[28] Schenkman M,Hall D,Kumar R,et al. Endurance exercise training to improve economy of movement of people with Parkinson disease:three case reports. Phys Ther,2008,88(1):63-76.
[29] Vernice JL,Javier B,Serge P. Animal models of Parkinson’s disease. Parkinsonism Relat Disord,2012,18(1):S183-S185.
[30] Gerecke KM,Jiao Y,Pani A,et al. Exercise protects against MPTP induced neurotoxicity in mice. Brain Res,2010,1341:72-83.
[31] Zigmond MJ,Cameron JL,Leak RK,et al. Triggering endogenous neuroprotective processes through exercise in models of dopamine deficiency. Parkinsonism Relat Disord,2009,15(Suppl 3):S42-S45.
[32] Yoon MC,Shin MS,Kim TS,et al. Treadmill exercise suppresses nigrostriatal dopaminergic neuronal loss in 6-hydroxydopamine-induced Parkinson’s rats. Neurosci Lett,2007,423:12-17.
[33] Tajiri N,Yasuhara T,Shingo T,et al. Exercise exerts neuroprotective effects on Parkinson's disease model of rats.Brain Res,2010,1310(15):200-207.
[34] Petzinger GM,Walsh JP,Akopian G,et al. Effects of treadmill exercise on dopaminergic transmission in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse model of basal ganglia injury. J Neurosci,2007,27:5291-5300.
[35] 徐严明,陶恩祥,丁小军,等. 反义寡核苷酸阻断多巴胺转运体表达的大鼠对MPIP反应性的改变. 中华医学遗传杂志,2008,25(2):236-241.
[36] Fisher BE,Petzinger GM,Nixon K,et al. Exercise-induced behavioral recovery and neuroplasticity in the 1-methyl-4 -phenyl -1,2,3,6 -tetrahydropyridine -lesioned mouse basal ganglia. J Neurosci Res,2004,77,378-390.
[37] Tillerson JL,Caudle WM,Reveron ME,et al. Exercise induces behavioral recovery and attenuates neurochemical deficits in rodent models of Parkinson’s disease. Neuroscience,2003,119(3):899-911.
[38] Marta GV,Quanzheng L,Beth F,et al. Exercise elevates dopamine D2 receptor in a mouse moudel of parkinson’s disease:in vivo imaging with [18F] Fallypride. Mov Disord,2010,25(16):2777-2784.
[39] VanLeeuwen JE,Petzinger GM,Walsh JP,et al. Altered AMPA receptor expression with treadmill exercise in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse model of basal ganglia injury. J Neurosci Res,2010,88(3):650-668.
[40] Wolf JA,Moyer JT,Lazarewicz MT,et al. NMDA/AMPA ratio impacts state transitions and entrainment to oscillations in a computational model of the nucleus accumbens medium spiny projection neuron. J Neurosci,2005,25:9080-9095.
[41] Wonju K,Mi JI,Cheol HP,et al. Remodeling of the dendritic structure of the striatal medium spiny neurons accompanies behavioral recovery in a mouse model of Parkinson’s disease. Neurosci Lett,2013,557:95-100.
[42] Takamatsu Y,Ishida A,Hamakawa M,et al. Treadmill running improves motor function and alters dendritic morphology in the striatum after collagenase-induced intracerebral hemorrhage in rats. Brain Res,2010,1355 (8):165-173.
[43] 刘晓莉,时凯旋,乔德才. 运动对帕金森病模型大鼠纹状体神经元电活动的影响. 北京体育大学学报,2014,37(5):57-61.
猜你喜欢
海外星云(2021年6期)2021-10-14 07:20:40
浙江医学(2020年9期)2020-07-01 10:17:42
卒中与神经疾病(2017年1期)2017-01-11 10:12:50
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国组织化学与细胞化学杂志(2016年4期)2016-02-27 11:15:55
磁共振成像(2015年7期)2015-12-23 08:53:04
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:21
医学研究杂志(2015年3期)2015-06-10 06:41:52
环球中医药(2015年4期)2015-02-27 15:01:51
中华皮肤科杂志(2014年3期)2014-12-19 12:54:41
