
不同瓣型茉莉水培生根能力及根尖解剖结构比较
2014-04-09 10:34邓衍明叶晓青梁丽建贾新平
植物资源与环境学报 2014年1期
邓衍明, 叶晓青, 梁丽建, 贾新平
(江苏省农业科学院农业生物技术研究所, 江苏 南京 210014)
茉莉(JasminumsambacAiton)又名茉莉花、茶叶花或叶子花,为木犀科(Oleaceae)素馨属(JasminumLinn.)常绿灌木,原产波斯湾附近,在地中海周边及亚洲多国均有分布。茉莉引入中国后,因其具有枝叶繁茂、株型玲珑、香气浓郁而不浊且清香持久等特点,在广西、福建、江苏、广东和浙江等地广为栽培。据不完全统计,中国的茉莉栽培面积约1×104hm2,占全球茉莉栽培总面积的2/3左右;而中国的茉莉鲜花产量则占全球总产量的50%以上[1-2]。茉莉花的茶用、药用和观赏价值均较高,素有“天下第一香”的美誉。
茉莉虽然品种较多,但依其花形结构可主要划分为单瓣、双瓣和重瓣(或多瓣)3种类型[1-2]。不同瓣型茉莉不仅在花朵形态和大小、产花量和香气类型等观赏性状和经济性状上存在差异,对温度、水分和光照等环境因子的适应性也不同。郭素枝等[3]研究了低温对单瓣和双瓣茉莉的影响,认为单瓣茉莉属于不耐寒植物,而双瓣茉莉属轻度耐寒植物;Deng等[4-5]比较了不同程度遮光处理对双瓣和重瓣茉莉生长发育和生理生化指标的影响,发现双瓣茉莉对弱光的适应能力强于重瓣茉莉;而郭素枝等[6]的研究结果显示:3种瓣型茉莉对干旱胁迫的耐性有差异,其中,双瓣茉莉对干旱胁迫的耐性强于单瓣和重瓣茉莉。至于不同瓣型茉莉存在生态适应性差异的原因,有研究者从器官或细胞结构方面进行了研究。如郭素枝等[7]比较了单瓣和双瓣茉莉营养器官的解剖结构特征,认为双瓣茉莉的叶片结构特征与单瓣茉莉不同,可能是其更适应低温和干旱生境的原因之一;Deng等[4-5]认为:双瓣茉莉叶片薄壁细胞中含有更多的叶绿体,使其在弱光条件下依然能保持较高的光合速率,从而导致其耐阴性强于重瓣茉莉;而郭素枝等[6]的研究结果表明:干旱胁迫对双瓣茉莉叶肉细胞超微结构的影响最小。然而,虽然根系对植物的正常生长以及耐寒和耐旱等抗逆性具有十分重要的作用[8],但目前对不同瓣型茉莉生根能力及根器官解剖结构差异的研究尚不多见。
花卉水培是以水(或溶液)为基质的一种新型无土栽培方法,可集赏花、赏叶、观根及观鱼等特点于一体,且具有便于管理、可规模化生产、易保持环境整洁等诸多优点,越来越受到人们的欢迎,应用前景广阔[9]。因茉莉生殖障碍严重(几乎不结实),只能以扦插和分株等无性方式进行繁殖[10-11],但目前有关茉莉离体再生和快繁技术体系的研究尚处于初级阶段[12-13],因而,开展茉莉水培生根研究具有重要的理论意义和现实意义,既有助于了解不同瓣型茉莉生态适应性差异的原因,又可拓宽种苗繁殖渠道、提升产品市场价值。有鉴于此,作者对水培条件下3种瓣型茉莉生根能力及根尖的解剖结构特征进行了观察和比较,以期为相关研究提供参考依据。
1 材料和方法
1.1 材料
实验于2012年5月至6月进行,供试3种瓣型茉莉均采自江苏省农业科学院茉莉种质资源圃。选取生长健壮、无病虫害的1年生半木质化枝条,剪取中、下段为插穗;插穗长8~12 cm,保留3个节和上部的4片叶,备用。
1.2 方法
1.2.1 插穗处理方法 为利于插穗生根,参照文献[14]的方法,在水培前先将插穗置于1 000 mg·L-1多菌灵溶液中消毒10 min,然后将插穗下端约1/3浸泡在200 mg·L-1α-NAA溶液中处理2 h。
1.2.2 水培生根及生根指标测量方法 参照文献[14]的方法进行水培,每种瓣型茉莉20支插穗,各设3次重复。培养至第30天,调查每种瓣型茉莉的生根率、单株生根数和根长等指标(仅统计长度大于或等于3 mm的根)。
1.2.3 根尖解剖结构观察 在每支插穗上随机剪取长度约5~10 mm的健壮根尖2条,分别用于根尖横切面和纵切面解剖结构的观察,每种瓣型插穗分别观察根尖20条。根尖取样后立即用FAA溶液固定,并于24 h后转入体积分数70%乙醇中保存。
参照文献[15-16]制作石蜡切片。已固定的根尖经乙醇逐级梯度脱水、渗蜡和包埋后,用KD2258型轮式切片机(金华市科迪仪器设备有限公司)切片,切片厚度6~8 μm;切片经二甲苯和乙醇逐级梯度透明、复水后用固绿染色,再经脱水和透明后用树脂封片。置于Olympus BX-43型光学显微镜(日本奥林巴斯公司)下观察,并用Olympus DP21型CCD成像系统(日本奥林巴斯公司)拍照。
拍照后每种瓣型茉莉选取位于根尖纵切面中间部位的图片10张,用标尺测量根冠宽度(根冠与分生区分界处的直径)和长度(分界处至根尖末端的距离)并计算根冠长/宽比[14]。为便于统计和比较,以根尖横切面成熟区气腔的个数代表每一根尖的气腔数量,以根尖纵切面伸长区最大气腔的长度和宽度代表根尖内气腔的长度和宽度。
1.3 数据分析
应用SPSS 7.0统计分析软件对测量数据进行差异显著性分析及相关性分析。
2 结果和分析
2.1 不同瓣型茉莉的水培生根能力分析
在水培条件下3种瓣型茉莉插穗的生根能力见表1。在水培条件下培养30 d后,单瓣、双瓣和重瓣茉莉插穗的生根率均达到100%,但不同瓣型茉莉的生根能力不同,单株生根数和平均根长均有显著差异(P<0.05)。总体而言,以双瓣茉莉的水培生根能力最强,单株生根数18.3条、平均根长23.9 mm,均显著高于单瓣和重瓣茉莉;单瓣茉莉的水培生根能力次之,单株生根数15.9条、平均根长18.6 mm;而重瓣茉莉的水培生根能力最差,单株生根数仅为7.8条,平均根长仅16.5 mm。此外,双瓣茉莉的单株生根数和根长的变异系数最小(分别为0.21和0.39),说明双瓣茉莉插穗生根一致;重瓣茉莉单株生根数变异系数最大(0.28)、单瓣茉莉根长变异系数最大(0.44),说明单瓣和重瓣茉莉插穗生根不够整齐,差异较大。
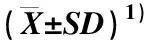
表1 水培条件下3种瓣型茉莉插穗的生根能力比较
2.2 不同瓣型茉莉根尖解剖结构比较
外观上看,3种瓣型茉莉插穗的水生根均较粗壮且色白。从3种瓣型茉莉根尖横切面和纵切面的解剖结构可见(图1):水生根根尖细胞的细胞质染色较浅,根冠细胞壁较薄,细胞内积累的内含物较少;根尖成熟区和伸长区都出现了大小不一、形状各异的细胞间隙或空腔(统称为气腔),形成了比较发达的类似水生植物根系通气组织状的结构(图1)。但不同瓣型茉莉根尖的解剖结构特征有一定差异。
3种瓣型茉莉根冠性状见表2。比较结果表明:单瓣、双瓣和重瓣茉莉根冠长度分别为550、503和480 μm,三者差异显著(P<0.05)。重瓣茉莉的根冠宽度最大,为395 μm,显著大于双瓣茉莉和单瓣茉莉(P<0.05);双瓣茉莉和单瓣茉莉的根冠宽度分别为375 和362 μm,二者差异不显著(P>0.05)。根冠长/宽比以单瓣茉莉最大(1.52)、双瓣茉莉次之(1.34)、重瓣茉莉最小(1.21),三者间的差异达显著水平(P<0.05)。3种瓣型茉莉的根冠长度和宽度的变异系数均较小且差异不大,说明不同瓣型茉莉的根冠形态性状相对稳定。
不同瓣型茉莉水生根的根尖内气腔大小和数量也有一定差异,水培条件下单瓣、双瓣和重瓣茉莉根尖内气腔性状见表3。比较结果表明:单瓣茉莉每根尖气腔数最少(平均每个根尖成熟区横切面可观察到3.1个气腔),与双瓣和重瓣茉莉差异显著(P<0.05);双瓣和重瓣茉莉每根尖气腔数分别为4.1和4.2个,两者差异不显著(P>0.05)。气腔宽度最大的是重瓣茉莉,气腔长度最大的是双瓣茉莉,单瓣茉莉的气腔长度和宽度均最小,且与双瓣和重瓣茉莉差异显著(P<0.05)。

A. 单瓣茉莉根尖Root tip of single-petal type of J. sambac: A1. 成熟区横切面Transverse section of maturation zone; A2. 伸长区纵切面Longitudinal section of elongation zone; A3. 根冠和分生区纵切面Longitudinal section of root cap and meristematic zone. B. 双瓣茉莉根尖Root tip of double-petal type of J. sambac: B1. 成熟区横切面Transverse section of maturation zone; B2. 伸长区纵切面Longitudinal section of elongation zone; B3. 根冠和分生区纵切面Longitudinal section of root cap and meristematic zone. C. 重瓣茉莉根尖Root tip of multi-petal type of J. sambac: C1. 成熟区横切面Transverse section of maturation zone; C2. 伸长区纵切面Longitudinal section of elongation zone; C3. 根冠和分生区纵切面Longitudinal section of root cap and meristematic zone. 箭头示气腔Arrows show air chambers.
表2 水培条件下3种瓣型茉莉根冠性状的比较
表3 水培条件下3种瓣型茉莉根尖内气腔性状的比较
总体上看,单瓣茉莉根尖细长,重瓣茉莉根尖短粗;双瓣茉莉根尖内气腔不仅数量多而且较大,形成的通气组织状结构也最为发达。
2.3 不同瓣型茉莉生根性状及根尖解剖结构特征间的相关性分析
水培条件下单瓣、双瓣和重瓣茉莉生根性状及根尖内根冠与气腔性状的相关系数见表4。由表4可以看出:水培条件下3种瓣型茉莉的生根指标与根冠性状及气腔性状指标间均有一定的相关性,但不同指标间的相关程度不同。其中,在单株生根数和平均根长这2个反映生根能力的指标中,单株生根数与根冠长度、根冠长/宽比和气腔长度呈正相关,与根冠宽度、每根尖气腔数和气腔宽度呈负相关;平均根长与每根尖气腔数呈负相关,与其他指标均呈正相关;但这2个生根能力指标与根尖各解剖性状间的相关性均未达到显著(P>0.05)水平。
根冠性状与气腔性状间也具有一定的相关性。其中,根冠长度与每根尖气腔数、气腔长度和气腔宽度均呈负相关,且与每根尖气腔数和气腔宽度间的相关性分别达到显著(P<0.05)和极显著(P<0.01)水平;根冠长/宽比与3个气腔性状间均呈极显著(P<0.01)负相关;根冠宽度与3个气腔性状间均呈正相关,但相关性不显著(P>0.05)。

表4 水培条件下3种瓣型茉莉根系各指标间的相关系数1)
3 讨 论
不同瓣型茉莉不仅花朵形态、经济性状和化学成分不同,其枝条和叶片形态以及生态习性等也存在差异[2]。植物的生态适应性不仅与其外部形态特征相关,与其组织器官的内在结构也密不可分。对相同生境下栽培的单瓣和双瓣茉莉营养器官解剖结构特征的研究结果[7]显示:与单瓣茉莉相比,双瓣茉莉的茎皮层细胞层更多、叶片角质层更厚、细胞排列更紧密,且这些特性是双瓣茉莉对低温和干旱环境适应性更强的原因之一;Deng等[4-5]比较了不同程度光照对双瓣和重瓣茉莉生长发育的影响,认为双瓣茉莉较强的耐阴性与其叶片薄壁细胞内叶绿体的分布更有利于进行光合作用等超微结构特征密切相关;邓传远等[17]比较了不同瓣型茉莉叶片超微结构的季节变化,也发现双瓣茉莉较强的抗高温(夏季)和抗低温(冬季)能力与其稳定的细胞超微结构有关。本研究中,不同瓣型茉莉不仅水培生根能力不同,水生根的根尖解剖结构特征也存在差异。根系是植物地下部分的主体,在植物体的正常生长发育过程中有着至关重要的作用;生根能力的强弱和根系的发达与否往往直接决定了植物的长势及其对环境的适应能力和对逆境胁迫的抗性等。双瓣茉莉的耐寒、耐旱、耐阴和抗病虫等抗逆性均强于其他2种瓣型茉莉[3-7],可能与双瓣茉莉较强的生根能力有关。
相关性分析结果表明:水培条件下茉莉的生根能力与根尖解剖结构特征的多个指标相关,尤其是根冠长度、根冠宽度、根冠长/宽比和气腔长度等指标贡献较大,说明不同瓣型茉莉水培生根能力受根尖解剖结构的影响较大,而且与根尖内部能否形成较为发达的气腔有一定关系。在上述相关指标中,根冠长度、根冠长/宽比和气腔长度等是最主要的正相关指标,每根尖气腔数和根冠宽度则是最主要的负相关指标。在3种瓣型茉莉中,单瓣茉莉虽然根冠长度和根冠长/宽比均最大,但其根尖内通气组织状结构最不发达,气腔长度和气腔宽度等指标均最小,故其生根能力弱于双瓣茉莉;而重瓣茉莉的根尖内虽发育出较为发达的通气组织状结构,但其根冠宽度最大,且根冠长/宽比最小,这是其生根能力最差的主要原因。然而,由于各指标所提供的信息发生重叠,说明它们与茉莉生根能力的相关性也不尽相同,故直接利用这些指标中的单一指标进行生根能力评价可能会具有一定的片面性。
为适应环境,水生植物和陆生植物的根系结构有明显区别[18]。前者的代表性植物水稻(OryzasativaLinn.),其皮层细胞会裂开并发育形成非常有利于根系通气的组织;而陆生植物一般不会分化出这种结构[19]。虽然通气组织形式多样、结构复杂,但其作用主要是作为O2进入根和植物体内的通道,故通气组织具有重要的生理意义[20];此外,通气组织还可用于直接排泄一些对植物有害的代谢废物、调节根际氧化势以缓解有害物质对植物体的毒害等[21-22]。因此,水培后陆生植物的根系形态和结构往往会随水生生境而发生相应改变。孔妤等[23]比较观察了土培和水培吊兰〔Chlorophytumcomosum(Thunb.) Baker〕的根系结构特征,结果显示水培吊兰根冠部位几乎不含晶细胞和淀粉体、髓部占中柱面积的比例变小,而质体会转变为叶绿体;赵兰枝等[24]比较了绿萝〔Epipremnumaureum(Linden et Andre) Bunting〕水培苗与土培苗根系外观形态、生长特性和组织结构等方面的差异,发现水生苗呈现出根系变细,表皮厚度、中柱直径以及中柱在根系横截面上所占的比例相应变小等现象。在水培条件下,茉莉水培苗根系同样存在根部细胞壁变薄、内含物变少等现象,与郭素枝等[7]报道的“单瓣和双瓣茉莉土生根的皮层薄壁细胞中有较多淀粉粒积累”等现象有明显差异;不仅如此,与此前报道[14]的双瓣茉莉类似,单瓣和重瓣茉莉在水培时根尖内均出现一定数量的细胞间隙或空腔等通气组织状结构,而这一结构在3种瓣型茉莉的土生根根尖内并不存在[7]。导致这一差异的原因可能在于:土壤或固体基质栽培环境下根的营养吸收、支持固定等功能非常重要,而水生环境下缺氧是影响植株正常生长的首要限制因子,根系运输O2功能得到强化,故分化出通气组织。在水培条件下3种瓣型茉莉根尖均分化出类似于气腔的通气组织状结构,说明它们的根系对水生环境都具有一定的适应性。但不同瓣型茉莉水培生根能力不同,表明其根系对水生环境的适应性不同,而这一差异与根尖解剖结构特征有一定关系。
张福平等[11]的研究结果表明:在固体基质上进行茉莉扦插实验,激素种类和浓度对其生根率、生根数、根长和根质量等指标均有显著影响;而双瓣茉莉的水培生根实验结果[14]也显示:激素处理不仅对其生根能力有显著影响,而且与根尖内部通气组织的形成也有密切关系;李小方等[25]认为:大多数情况下不定根的发生是由于植物器官受伤或受到激素、病原微生物的刺激而表现出的再生反应。本研究中,在进行水培前3种瓣型茉莉插穗基部均进行了激素处理,因此,不同瓣型茉莉插穗水培生根能力及根尖内通气组织状结构的差异,究竟是对激素处理的响应不同还是各瓣型茉莉自身遗传机制不同所致?尚需进一步研究。
参考文献:
[1] 董利娟, 张曙光. 茉莉花的生产现状与科研方向[J]. 茶叶通讯, 2001(2): 11-13.
[2] 谢玲玲, 黄法就, 蔡 汉. 茉莉花的栽培品种及其比较[J]. 现代农业科技, 2007(3): 26-27.
[3] 郭素枝, 邓传远, 张国军, 等. 低温对单、双瓣茉莉叶片细胞膜透性的影响[J]. 中国生态农业学报, 2006, 14(1): 42-44.
[4] DENG Y M, LI C C, SHAO Q S, et al. Differential responses of double petal and multi petal jasmine to shading: Ⅰ. Photosynthetic characteristics and chloroplast ultrastructure[J]. Plant Physiology and Biochemistry, 2012, 55: 93-102.
[5] DENG Y M, SHAO Q S, LI C C, et al. Differential responses of double petal and multi petal jasmine to shading: Ⅱ. Morphology, anatomy and physiology[J].Scientia Horticulturae,2012, 144: 19-28.
[6] 郭素枝, 张明辉, 邓传远, 等. 干旱胁迫对茉莉3个品种叶片光合特性和超微结构的影响[J]. 热带亚热带植物学报, 2013, 21(2): 123-129.
[7] 郭素枝, 邓传远, 张育松, 等. 单、双瓣茉莉营养器官解剖结构特征及其生态适应性研究[J]. 中国生态农业学报, 2004, 12(3): 40-43.
[8] DENG Y M, CHEN S M, CHANG Q S, et al. The chrysanthemum×Artemisiavulgarisintergeneric hybrid has better rooting ability and higher resistance to alternaria leaf spot than its chrysanthemum parent[J]. Scientia Horticulturae, 2012, 134: 185-190.
[9] 郭世荣. 无土栽培学[M]. 北京: 中国农业出版社, 2003: 1-2.
[10] 赖明志, 连长伟, 杨如兴. 茉莉育性的研究[J]. 福建农业大学学报, 1996, 25(4): 442-445.
[11] 张福平, 杨少珍. 茉莉花扦插试验研究[J]. 北方园艺, 2007 (11): 144-146.
[12] 孙艳妮, 汤访评, 房伟民, 等. 茉莉离体快繁体系的建立[J]. 浙江农业学报, 2009, 21(4): 390-394.
[13] 李聪聪, 陆长梅, 佘建明, 等. 双瓣茉莉离体微繁技术[J]. 江苏农业科学, 2012, 40(4): 65-68.
[14] 邓衍明, 叶晓青, 李聪聪.α-萘乙酸对茉莉水培生根及根尖解剖结构的影响[J]. 江西农业学报, 2012, 24(2): 35-37.
[15] DENG Y M, CHEN S M, TENG N J, et al. Flower morphologic anatomy and embryological characteristics inChrysanthemummulticaule(Asteraceae)[J]. Scientia Horticulturae, 2010, 124: 500-505.
[16] DENG Y M, JIANG J F, CHEN S M, et al. Combination of multiple resistance traits from wild relative species inChrysanthemumvia trigeneric hybridization[J]. PLoS One, 2012, 7(8): e44337.
[17] 邓传远, 郭素枝, 王湘平, 等. 茉莉3品种叶肉细胞超微结构的季节变化研究[J]. 热带作物学报, 2012, 33(2): 267-273.
[18] 强 胜. 植物学[M]. 北京: 高等教育出版社, 2006: 82-91.
[19] 汪 强, 苏 菊, 孙合金, 等. 水培花卉水生根系诱导研究初报[J]. 中国农学通报, 2008, 24(1): 60-63.
[20] KLUDZE H K, DELAUNE R D, PATRICK W H Jr. Aerenchyma formation and methane and oxygen exchange in rice[J]. Soil Science Society of America Journal, 1993, 57(2): 386-391.
[21] JACKSON M B, ARMSTRONG W. Formation of aerenchyma and the processes of plant ventilation in relation to soil flooding and submergence[J]. Plant Biology, 1999, 1(3): 274-287.
[22] GUNAWARDENA A H L A N, PEARCE M D, JACKSON M B, et al. Characterisation of programmed cell death during aerenchyma formation induced by ethylene or hypoxia in roots of maize (ZeamaysL.)[J]. Planta, 2001, 212: 205-214.
[23] 孔 妤, 王 忠, 顾蕴洁, 等. 土培和水培吊兰根系结构的观察[J]. 园艺学报, 2009, 36(4): 533-538.
[24] 赵兰枝, 郑树景, 郭瑞珍, 等. 水培与土培绿萝的根系形态及结构比较[J]. 广东农业科学, 2010(11): 121-122.
[25] 李小方, 汤章城, 何玉科. 不定根的形态发生与调节机制[J]. 细胞生物学杂志, 2001, 23(3): 130-136.
猜你喜欢
少儿科学周刊·儿童版(2022年12期)2022-07-09
少儿科学周刊·少年版(2022年12期)2022-07-08
学校教育研究(2021年21期)2021-05-24
食品与生物技术学报(2021年8期)2021-01-17
学校教育研究(2021年23期)2021-01-02
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
现代农业科技(2017年16期)2017-09-22
现代园艺(2017年1期)2017-07-14
农村百事通(2016年7期)2016-05-14
