
不同生育时期的牡丹叶片和花瓣光合相关生理指标变化分析
2014-04-08 01:05李永华杨秋生
河南农业大学学报 2014年1期
崔 洋,张 凌,李永华,杨秋生
(河南农业大学林学院,河南 郑州 450002)
牡丹是中国重要的园林观赏植物,其开花过程需要消耗大量能量以满足各种物质的合成及新陈代谢的需求,叶片和花瓣会随着开花及衰败发生一系列的变化.在成花过程中,植物体内淀粉、蔗糖等碳水化合物含量及组成比例对开花具有关键作用[1].植物通过光合作用合成碳水化合物,为开花提供能量物质,这是影响牡丹开花质量的重要因素之一[2,3].史国安等[4]研究表明,牡丹花瓣中总可溶性糖呈迅速增加的趋势,特别是己糖(葡萄糖和果糖)含量显著增加,盛开后己糖水平达到最高,而蔗糖含量呈现逐渐下降的变化,牡丹花瓣中己糖的积累在花开放和衰老过程中起着重要作用.翟敏等[5]、刘志敏等[6]探讨了盆栽和地栽牡丹年周期光合作用变化和根系碳氮代谢动态,张开明等[7]对根、茎中的糖组分含量及变化情况进行了分析,然而对开花过程中叶片和花瓣中糖代谢的组分及含量研究较少.因此,本试验以牡丹品种‘胡红’为试材,对开花过程中叶片和花瓣碳水化合物组分、含量以及光合特性指标进行测定,分析糖组分的变化规律,为牡丹花期调控技术提供理论基础.
1 材料与方法
1.1材料
试验材料为6年生牡丹(Paeoniasuffruticosa)品种‘胡红’,河南农业大学科教园区内露地栽培.选择生长健壮、长势一致的植株,于风铃期、圆桃期、破绽期、开花期、谢花期5个时期,取叶片和花瓣用于生理指标测定.花瓣取自外缘向内的第3~5层小花,叶片取茎中段第3~5片复叶.所取样品迅速洗净擦干,塑料袋封装,放入液氮罐带回实验室,置于-70 ℃冰箱内保存备用.
1.2方法
1.2.1 糖组分测定 蔗糖、葡萄糖、果糖的测定参考谢复炜等[8]的方法,采用离子色谱仪(Dionex ICS-3000,Thermo,USA)测定.采用CarboPac TMPA10(4×250 mm)糖分析柱和CarboPac TMPA10(4×50 mm)糖保护柱,流动相A:蒸馏水;流动相B:250 mmol·L-1NaOH溶液,2个流动相体积比V(A)∶V(B)=55∶45,积分脉冲安培检测法检测,流速为0.7 mL·min-1,柱温40 ℃,进样量25 μL,所有样品进样前过0.22 μm滤头,3次重复.
1.2.2 淀粉、叶绿素含量测定 淀粉、叶绿素含量测定参考李合生[9]的方法,3次重复.
1.2.3 光合参数测定 以牡丹品种‘胡红’为试材,分别在圆桃期、破绽期、开花期、谢花期4个时期各选晴好天气进行光合参数的测定.测定时选取长势一致、无病虫害的植株,使用便携式光合作用测定仪(Li-6400,LI-COR,USA)进行测定,叶片选取完全展开的第2复叶的顶小叶.上午10∶00~11∶30进行光合作用参数测定,包括净光合速率(Net photosynthetic rate, Pn)、蒸腾速率(Transpiration rate,Tr)、气孔导度(Stomatal conductance ,Gs)、胞间CO2浓度(Intercellular CO2concentration,Ci),3次重复.
2 结果与分析
2.1不同生育时期糖组分在不同牡丹器官中的变化
在不同生育时期,叶片和花瓣中葡萄糖、果糖含量变化相似,前期呈现上升趋势,破绽期达到最高,与风铃期相比分别升高44.63%、49.06%;花瓣中葡萄糖、果糖含量开花期达到最高点,分别升高66.49%、72.53%,花瓣中果糖含量在开花期比叶片增加27.64%;随着花瓣的衰老,葡萄糖和果糖的含量逐渐降低(表1).由表1可知,破绽期叶片中蔗糖含量比风铃期升高42.9%,开花期达到最低点(下降25.31%),而谢花期则升高170.5%;花瓣中蔗糖含量逐渐上升,但一直低于叶片.在不同生育时期,叶片和花瓣的葡萄糖、果糖、蔗糖含量差异显著.
2.2不同生育时期牡丹同一器官中糖组分的变化
叶片中葡萄糖和果糖含量变化趋势均为先增加后下降(图1-A),从风铃期到破绽期,葡萄糖和果糖含量分别升高了0.8倍与0.96倍,而蔗糖含量下降.从图1-B中看出,试验期间花瓣中的葡萄糖、果糖含量显著增加,开花期葡萄糖、果糖含量分别为圆桃期的2.98倍和3.64倍,蔗糖含量降低.牡丹花发育过程中,蔗糖水解为葡萄糖和果糖,为细胞生长提供能量.
2.3不同生育时期牡丹叶片和花瓣中淀粉含量变化
淀粉是植物体内的主要储藏营养物质.牡丹叶片和花瓣淀粉含量变化的总体趋势一致,均为先上升后下降(图2).叶片中淀粉含量高于花瓣,在破绽期升到最高,为风铃期的1.8倍;而花瓣则在开花期达到顶峰,为圆桃期的2.98倍.淀粉含量在开花期达到最高,能够为牡丹开花提供充足的能量供应,这是决定牡丹能否正常开花的关键因素之一.
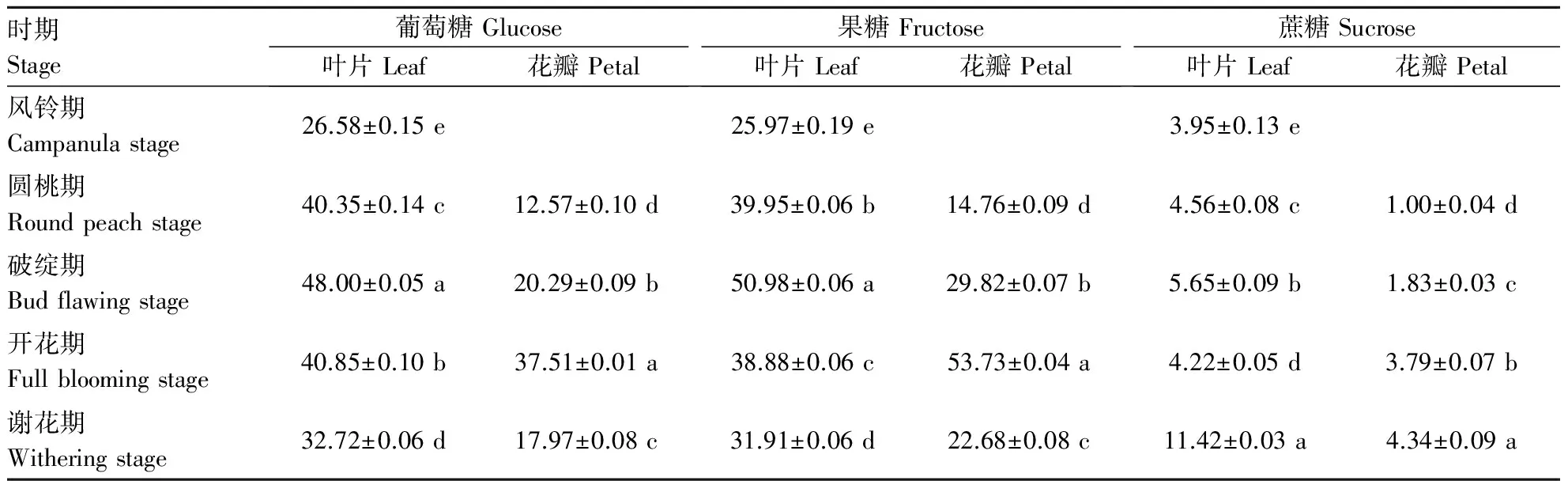
表1 不同生育时期葡萄糖、果糖和蔗糖在不同牡丹器官中的含量变化

图1 不同生育时期叶片(A)和花瓣(B)中糖组分含量变化

图2 不同生育时期叶片和花瓣中淀粉含量变化
2.4不同生育时期牡丹叶绿素含量的变化
由表2中可知,整个开花前后不同生育时期牡丹叶绿素a、叶绿素b、叶绿素总含量及叶绿素a/b值的变化一致,均是先升高后下降.展叶初期,三者含量较低,随着幼叶的成长,破绽期含量最高,从风铃期到破绽期,叶绿素a、叶绿素b和叶绿素总含量分别增加了69.15%、68.29%和65.71%.在开花前后,叶绿素b含量一直低于叶绿素a的含量,牡丹叶片中叶绿素a/b值均在2.0以上.
2.5不同生育时期牡丹光合作用的变化
由表3可知,Pn呈现先升高后降低的趋势,牡丹开始展新叶的时候Pn较低,随着叶片的发育在破绽期Pn迅速达到峰值,比圆桃期增加47.55%,随着花瓣的衰老Pn逐渐下降,谢花期降低30.94%.叶片Gs变化与Pn变化相似,在破绽期最大,之后呈下降趋势,谢花期时下降32.28%.Tr变化一直处于上升状态.破绽期后Gs下降,但Ci逐渐增加,Pn下降,说明牡丹叶片光合能力逐渐减弱,这可能与牡丹叶片发育进程及牡丹花发育对碳水化合物的需求量减少有关.
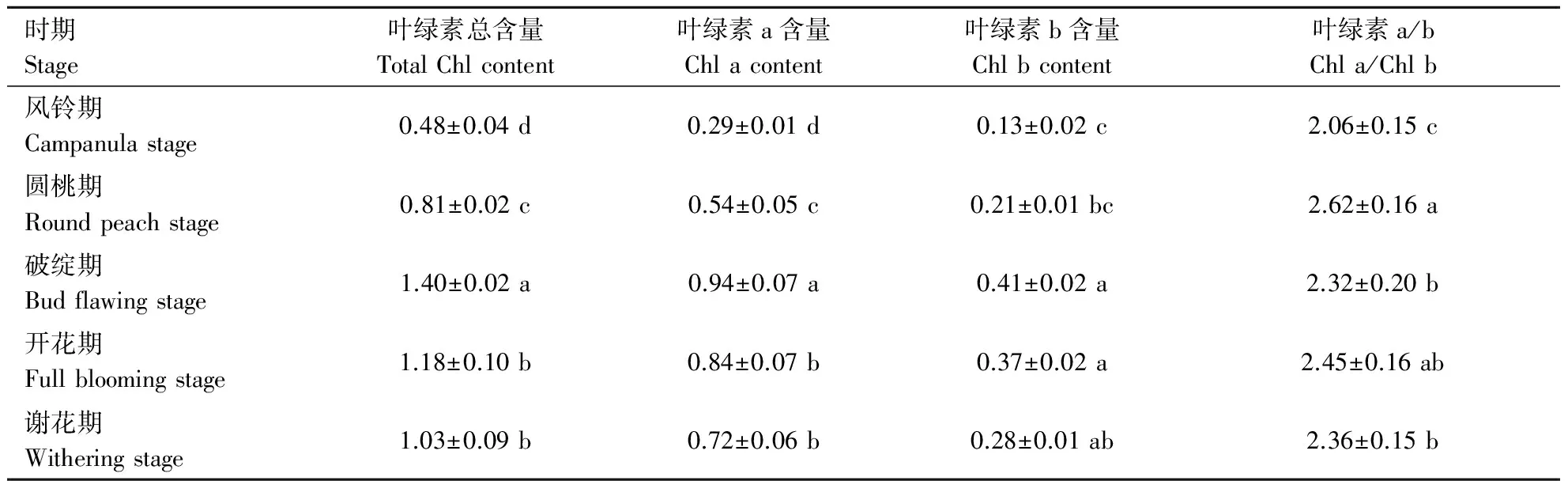
表2 不同生育时期牡丹叶片叶绿素含量变化

表3 不同生育时期牡丹叶片净光合速率、气孔导度、蒸腾速率和胞间CO2浓度的变化
3 讨论
牡丹开花过程需要足够的能量来促进开花,可溶性糖与淀粉含量的高低影响牡丹开花的品质与数量.柴叶茂等[10]分析草莓果实发育过程中可溶性糖含量的变化,研究表明,蔗糖、葡萄糖和果糖含量随着草莓果实发育均呈逐渐增加的趋势,尤其蔗糖积累与果实成熟的关系较密切.分析牡丹开花前后根、枝条中的各种营养成分的变化,发现牡丹开花是明显的能量消耗过程,根是植株的主要养分贮存器官,为开花提供所需的营养物质[7,11].蔗糖是研究最多的与成花过程有关的可溶性糖,蔗糖代谢是糖积累的关键环节[12,13].本试验研究表明,在牡丹不同生育时期,叶片和花瓣中碳水化合物含量呈先上升后下降的趋势.叶片中葡萄糖、果糖含量在破绽期达到最大值,蔗糖含量也逐渐上升,可能是因为牡丹开花前叶片逐渐变大和数量增多产生大量的同化物;破绽期之后叶片中葡萄糖、果糖含量开始下降,可能是因为叶片的衰老促使可溶性糖急剧外运,植株本身可溶性糖合成能力下降;另一方面可能是随着花蕾的逐渐形成,生殖生长逐渐增强,开花时需要大量的营养成分和较多的能量,从而导致叶片中葡萄糖、果糖含量下降.牡丹花期较短,延长花期、提高观赏品质是满足牡丹市场供应需要解决的关键问题.适当遮阴可以有效延长露地牡丹花期,但由于光照不足导致还原性糖含量过低,进而造成花朵较小、花色较浅.盆栽牡丹冬季催花过程中,所需能量主要来源于根系贮藏的有限的碳水化合物,适当补充还原性糖能够改善牡丹的催花品质.本试验探讨了牡丹开花前后糖组分的变化规律,为完善牡丹的花期调控技术提供了理论基础.
叶绿素含量的多少直接影响到光合作用的强弱.随着土壤干旱胁迫程度的增加,叶绿素含量显著减少,牡丹叶片的光合潜力受到显著抑制[14,15].翟敏等[5]研究表明,盆栽和地栽牡丹较强的光合能力主要集中在适宜生长的春季4、5月份,以后逐渐下降,9月份降至最低,大风铃期和叶片放大期,叶绿素a、叶绿素b和叶绿素总量盆栽牡丹均低于地栽牡丹.本研究结果表明,破绽期叶片叶绿素a、叶绿素b和叶绿素总量含量达到最高,谢花期叶绿素含量迅速下降,可能是因为花衰败后不再有充足的碳水化合物等营养物质向上运输,另外可能是因为高温强光使其破坏或降解速度超过合成速度.随着气温升高,牡丹花瓣衰败,开花期过后牡丹Pn下降,谢花期叶绿素含量下降.牡丹叶片叶绿素含量高,Pn也高,叶绿素含量与Pn之间呈一定的正相关,这与之前的研究结果一致[16].
牡丹不同生育时期叶片和花瓣中葡萄糖、果糖和蔗糖含量发生明显变化,这种变化与品种类型、源-库关系、肉质根贮藏物质以及花瓣的程序性死亡、乙烯等激素的调控等相关,并且与牡丹花的衰老过程相一致.牡丹叶片和花瓣中葡萄糖、果糖和蔗糖除了作为能源物质和渗透调节物质以外,在不同生育时期叶片和花瓣中的信号转导机制以及调控途径仍需要进一步研究.
参考文献:
[1]LIU C, THONG Z, YU H. Coming into bloom: the specification of floral meristems [J]. Development, 2009, 136: 3379-3391.
[2]侯小改,段春燕,刘改秀,等.土壤含水量对牡丹光合特性的影响[J].华北农学报,2006,21(2):91-94.
[3]丰亚南,郑国生,王宗正,等.牡丹开花前后碳水化合物的分配与光合速率的关系[J].园艺学报,2007,34(1):153-156.
[4]史国安,郭香凤,张国海,等.牡丹开花和衰老期间花瓣糖代谢的研究[J].园艺学报,2009,36(8):1184-1190.
[5]翟 敏,李永华,杨秋生.盆栽和地栽牡丹光合特性的比较[J].园艺学报,2008,35(2):251-256.
[6]刘志敏,孔德政,李永华,等.盆栽和地栽‘洛阳红’牡丹根系的碳氮代谢动态[J].林业科学,2008,9(44):162-164.
[7]张开明,王翠丽,王 丹,等.牡丹不同生育期茎与根系糖组分的动态变化[J].河南农业大学学报,2013,47(3):284-288.
[8]谢复炜,赵晓兰,宋志超,等.烟草中果胶的离子色谱分析方法研究[J].西北大学学报,2010,40:215-216.
[9]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[10]柴叶茂,贾海锋,李春丽,等.草莓果实发育过程中糖代谢相关基因的表达分析[J].园艺学报,2011,38(4):637-643.
[11]高志民,王 雁,李振坚,等.牡丹开花前后营养变化分析研究[J].林业科学研究,2007,20(3):390-393.
[12]DAVIES C, ROBINSON S P. Sugar accumulation in grape berries: Cloning of two putative vacuolar invertase cDNAs and the expressioning grape vine tissues[J]. Plant Physiol, 1996, 111:275-283.
[13]SUNG S S, SHEIH W J, GEIGER D R,Growth, sucrose synthase and invertase activities of developingPhaseolusvulgarisL. Fruit[J].Plant Cell and Environment, 1994, 17: 419-426.
[14]侯小改,段春燕,刘素云,等.不同土壤水分条件下牡丹的生理特性研究[J].华北农学报,2007,22(3):80-83.
[15]张 锋,孔祥生,范志业,等.逐渐干旱对牡丹光合和荧光特性的影响[J].中国农学通报,2008,24(12):251-255.
[16]张其德,卢从明,张启峰,等.几组杂交组合的杂交稻及其亲本光合特性的比较研究[J].生物物理学报,1996,12(3):511-516.
猜你喜欢
中成药(2021年5期)2021-07-21
广西糖业(2020年3期)2020-09-25
学生天地(2020年2期)2020-08-25
食品界(2019年8期)2019-10-07
浙江工业大学学报(2017年5期)2018-01-22
现代园艺(2017年13期)2018-01-19
连环画报(2017年1期)2017-07-13
中国糖料(2016年1期)2016-12-01
北方音乐(2016年12期)2016-08-23
中成药(2016年4期)2016-05-17
