
施氮量对枣棉间作棉花干物质积累、 产量与品质的影响
2014-04-08 11:16:14阿丽艳肉孜郭仁松张巨松
植物营养与肥料学报 2014年3期
阿丽艳·肉孜, 郭仁松, 杜 强, 武 辉, 张巨松*
(1 新疆农业大学,教育部棉花工程研究中心,乌鲁木齐 830052; 2 新疆农科院经济作物研究所,乌鲁木齐 830052)
枣棉间作是目前南疆棉花主产区普遍推广的果棉立体种植模式之一,其改变了以往单一枣树或棉花的平面种植,能有效提高土、 水、 热、 光资源的利用率,增加经济产值,有利于农田生态系统的良性循环和农业的可持续发展。但枣棉间作种植模式中枣树会对棉花产生一定的荫蔽效应,影响棉花生长发育、 产量以及棉纤维品质。光照和氮素是影响棉花生长最为重要的两个因素,对作物生物量的累积、 产量和品质有决定性的作用。在生产中氮素营养是调控作物生长及光合生产的重要手段之一[1-2]。作物的高产是以较高的生物量为前提,而生物量累积又是以氮素吸收为基础[3]。前人研究认为棉花生物量的累积特征符合Logistic模型,氮素对增长模型的基本形态影响较小,但对增长速率影响较大[4-6],因此可以通过不同的施氮量来调节棉花快速生长期的生长特征值,达到提高产量和品质的目的。施氮量过高时棉花生物量增加幅度变缓,会降低其品质[7]。在产量方面,前人研究指出,作物过多吸收的养分不仅没有转换成产量,反而影响作物生长,使产量下降,同时也造成了养分的浪费[8]。施氮量对棉纤维伸长率、 绒长、 马克隆值影响比较显著,对比强度、 整齐度影响较小[9]。弱光会导致棉纤维马克隆值及比强度下降,纤维细度和成熟度降低,棉纤维加长[10-11]。前人对单作系统中氮肥的影响效应研究较为多,而对间作复合系统中棉花干物质积累和不同部位产量构成因素以及棉纤维品质的研究较少。本文研究了枣棉间作条件下不同氮素水平对棉花生物量积累和产量以及棉花植株上、 中、 下不同部位棉纤维品质的影响,以期为枣棉间作模式的合理施氮提供依据。
1 材料与方法
1.1 试验设计

表1 供试土壤基本肥力状况
1.2 测定项目与方法
干物重的测定 分别于棉花现蕾、 开花、 盛花、 盛铃和吐絮期采用挖根法取植株样,挖根深度现蕾期为0—60 cm、 开花期、 盛花为0—140 cm,盛铃和吐絮期为0—160 cm。在每个试验小区的近冠和远冠区分别选取有代表性的棉株3株,样品洗净后在105℃下杀青30 min,80℃烘至恒重,测定其干物质重。
产量和产量构成因素的测定于9月10日调查各处理小区第2膜所有株数及总铃数; 10月20日调查霜前花率,计算单株有效结铃数; 在棉株吐絮时,选取60个吐絮铃,从下到上即在第3果枝(下)、 第4果枝(中)、 和第7果枝(上)3个部位各取20个吐絮铃,称重得到单铃重。轧花后将皮棉重量与铃重相除可得衣分,皮棉产量为收获株数、 单株结铃数、 单铃重和衣分的乘积。
棉纤维品质的测定 取各处理的棉花样20 g,由农业部棉花质量监督检验测试中心(乌鲁木齐)测定棉纤维长度、 比强度、 整齐度以及马克隆值。
数据处理采用Excel 2007、 DPS 7.05软件,LSD法进行方差分析。
2 结果与分析
2.1 施氮量对枣棉间作棉花根系和地上部干物质积累特点的影响
由表2可知,不同施氮处理棉花的干物质积累量在近冠区与远冠区均表现为有为N2>N1>N3>N0,差异显著,干物质快速积累持续天数近冠区与远冠区表现为N2>N1>N0>N3,N3处理的干物质积累最大速率出现时间较早,说明较高施氮量可促进棉株提早进入旺盛的营养生长期,但由于持续时间短,不利于光合产物的积累。
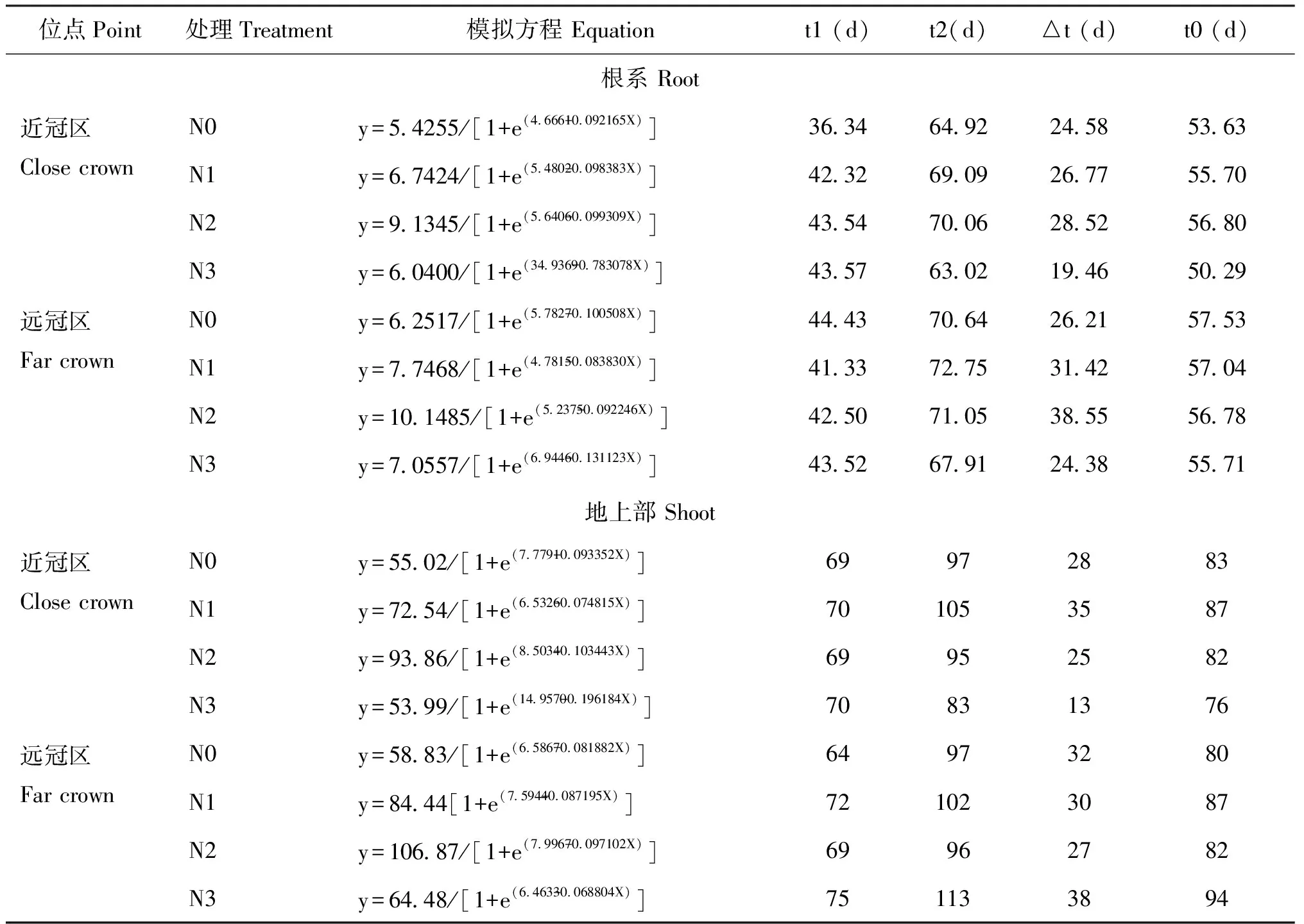
表2 棉花根系和地上部干物质积累模型参数特征值
2.2 施氮量对枣棉间作棉花产量及其构成因素的影响
由表3可以看出,各处理间棉花产量与棉花生长量的趋势一致。N2处理的单株结铃数显著大于其他处理(P>0.05)。单铃重各处理间均表现为N2>N1>N3>N0,施氮量较低的处理衣分值较高,各处理间皮棉产量差异达极显著水平。N0、 N1、 N2、 N3处理皮棉产量近冠区比远冠区分别少367.0、 256.5、 95.7和132.1 kg/hm2,说明光照对棉花产量有较大影响。在同一冠区,N1处理的皮棉产量均低于N2,而N2处理在近冠区的产量为2076.2 kg/hm2,远冠区为2171.9 kg/hm2,也为N2高于N1,说明氮素和光照均对棉花产量有显著影响。

表3 施氮量对棉花产量和产量构成因素的影响
2.3 施氮量对枣棉间作棉花不同部位单铃重和衣分的影响
中氮(N2)与不施氮(N0)处理棉株上部衣分均表现为远冠区大于近冠区,且近冠区最大衣分值出现在N1处理,远冠区最大衣分值出现在N3处理; 不同施氮量对棉株中部衣分无太大影响,远冠区略高于近冠区; 棉株下部近冠区以N0处理、 远冠区N3处理有较高的衣分,说明遮荫降低棉花的衣分值,并且上部棉铃需要较多的氮素用于纤维的形成。
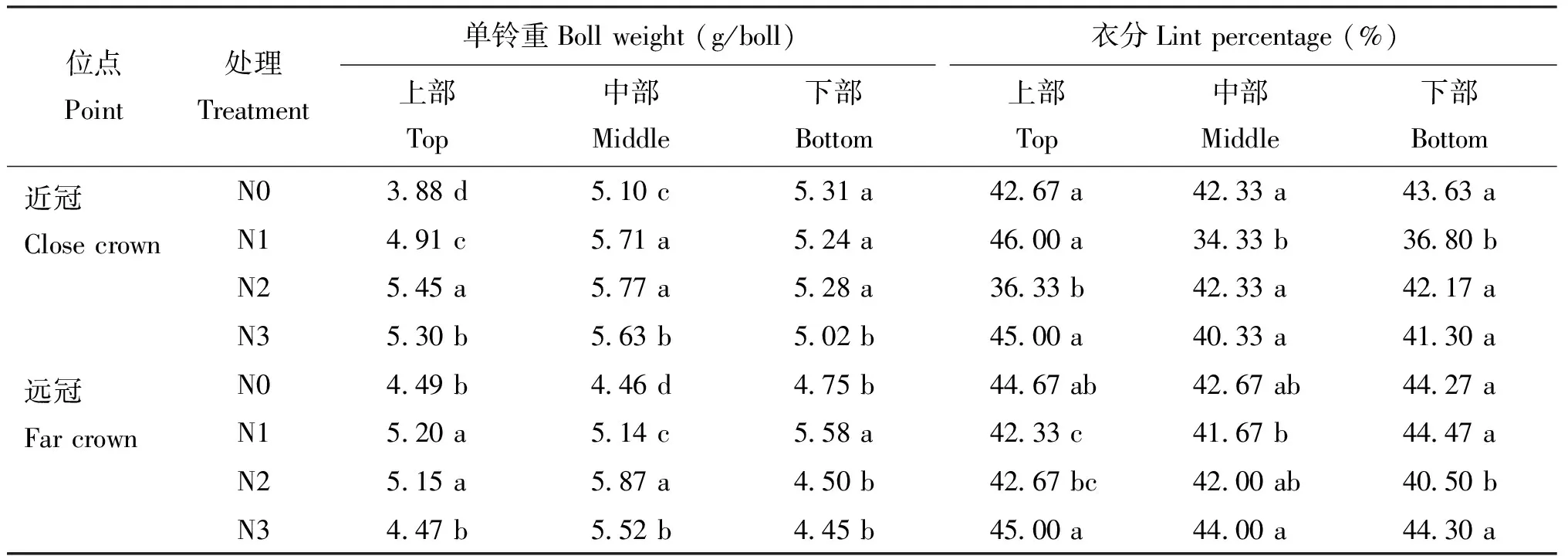
表4 施氮量对棉花不同部位单铃重和衣分的影响
2.4 施氮量对枣棉间作棉花纤维品质的影响
2.4.1施氮量对枣棉间作棉花植株上部棉纤维品质的影响 棉纤维整齐度、 比强度、 马克隆值随施氮量的提高呈先增后降的趋势(表5)。棉纤维长度和比强度在N0、 N1处理表现为近冠区低于远冠区,N2、 N3则表现为近冠区高于远冠区,而整齐度与施氮量的关系与棉纤维长度和比强度的趋势相反。各处理近冠区棉纤维伸长率和马克隆值小于远冠区。N1、 N2处理的棉纤维长度、 整齐度、 比强度显著高于N3,说明施氮过多使棉纤维品质变劣。马克隆值是细度和成熟度的综合指标,马克隆值大表明成熟度高,纤维粗。因此,栽培上不能单纯强调细度或成熟度。随施氮量增加,近冠区马克隆值为N2>N1>N0>N3,远冠区为N2>N1>N3>N0,与陈亮等[12]得出的施氮使马克隆值减小的结论不一致。

表5 不同氮素处理棉花不同部位的纤维品质
2.4.2 施氮量对枣棉间作棉花植株中部纤维品质的影响 如表5所示, 棉纤维整齐度和马克隆值随施氮量增加呈先升后降趋势,纤维长度和马克隆值在N0、 N1处理中表现为近冠区小于远冠区,N2、 N3则相反; 整齐度与施氮量的关系与纤维长度和马克隆值相反,并且近冠区以N1、 远冠以N2整齐度最高。纤维比强度近冠区均大于远冠区,近冠区纤维比强度随施氮量增加先增后减,远冠区则呈持续上升的趋势。对于植株中部的棉纤维来说,施氮量和间作对其马克隆值和比强度影响较大。间作遮阴棉花在较低施氮量下有较好的整齐度,较高施氮量下有较好的纤维长度和较高的马克隆值,同时遮阴增加了中部植株棉纤维的比强度。
2.4.3 施氮量对枣棉间作棉花植株下部纤维品质的影响 棉纤维长度各处理近冠区均小于远冠区,N1、 N2与N3差异显著; 伸长率则是近冠区大于远冠区,并且表现为N3>N0>N2>N1。比强度随施氮量的增加呈先升后降的趋势,N1、 N2与N0差异显著。马克隆值在近冠区为N1>N2>N3>N0,远冠区则随施氮量增加而增大,整体呈下部>中部>上部。说明氮肥不足或过多会增大棉纤维伸长率和短绒指数,降低其长度和比强度。
3 讨论与结论
作物的高产是以较高的生物量为前提的,说明作物生物量与产量有密切关系。薛晓萍、 张旺峰等研究表明,棉花生物量动态积累曲线均符合Logistic模型[5, 13],肥料因子并不改变其生长模型,但可以影响特征参数,本文与其结论一致,并且认为棉花单株生物量积累对氮肥反应比较敏感。前人的研究结果显示,施氮能增加作物产量,但达一定限度后产量不再增加[14]。农林间作下农作物产量的试验结果也不尽相同[15-16]。优化氮肥管理措施能够显著提高棉花的纤维长度和比强度,并确保马克隆值处于最优的品质范围内。在棉花生长的任何时期遮阴都会降低纤维比强度、 成熟度和马克隆值。间作棉花在较低的施氮量条件下有较好的整齐度,在略高施氮量时有较好的棉纤维长度和比强度以及马克隆值,同时遮阴减小了棉纤维伸长率。与棉花植株上部比较,中部棉铃对氮的需求较高,并且施氮不足影响的主要指标是比强度和马克隆值,而氮肥过量影响的则是纤维长度和整齐度。对于间作和氮肥对棉纤维品质的影响程度方面,对上部和下部的影响大于中部。而中部棉铃是产量形成的关键部位,因此合理的氮肥调控不但能保证棉株中层的产量输出,而且可促进上部和下部棉产量和品质的最优化。
本研究结果表明,枣棉间作棉花生物量动态积累曲线符合Logistic模型,并且棉花单株生物量积累与根系生物量积累对氮肥的反应更加敏感。近冠区N2处理可以获得较高的棉花生物累积量,从而获得较大的干物质积累速率和较早的最大速率出现时间,利于最终产量的形成。
枣棉间作下施氮能提高棉花单株结铃数和单铃重,不同施氮量对衣分的影响不大,N0和N2处理皮棉产量表现为近冠区大于远冠区,N1和N3则反之,且不论近冠区还是远冠区N2产量最高。相比较而言,较高的施氮量能增大上部铃重和衣分值,但不利于下部棉铃的生长发育,因此可根据品种和株型来确定适宜的氮肥用量。本试验认为,在枣棉间作条件下,过量施氮更易加重棉花受互作效应中不利因素的胁迫程度,因此总施氮量在450 kg/hm2为宜。
参考文献:
[1] Read S M, Northcote D H. Minimization of variation in the response to different protein of the coomassic blue gdye dinding: Assay for protein[J]. Analytical Biochemistry, 1981, 116: 53-64.
[2] 邹琦. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2000. 72-75.
Zou Q. The guidance of plant physiology experiments[M]. Beijing: China Agriculture Press, 2000. 72-75.
[3] 孟亚利,曹卫星,柳新伟,等.水稻地上部干物质分配动态模拟的初步研究[J]. 作物学报,2004,30(4): 376-381.
Meng Y L, Cao W X, Liu X Wetal. A preliminary study of simulation on shoot dry matter partitioning in rice[J]. Acta Agronomica Sinica, 2004, 30(4): 376-381.
[4] 伍维模, 郑德明, 董合林, 等. 南疆棉花干物质和氮磷钾养分积累的模拟分析[J]. 西北农业学报, 2002, 11(1): 92-96.
Wu W M, Zheng D M, Dong H Letal. Simulating the dynamics of dry matter and N, P2O5, K2O accumulation of cotton in South Xinjiang[J]. Journal of Northwest Agriculture, 2002, 11(1): 92-96.
[5] 薛晓萍, 王建国, 郭文琦, 等. 氮素水平对初花后棉株生物量、 氮素积累特征及氮素利用动态变化的影响[J]. 生态学报, 2006, 26(11): 3631-3640.
Xue X P, Wang J G, Guo W Qetal. Effect of nitrogen application levels on the dynamics of biomass, nitrogen accumulation and nitrogen recovery rate of cotton after initial flowering[J]. Acta Ecology Sinica, 2006, 26(11): 3631-3640.
[6] 何萍, 金继运, 林葆, 等. 不同氮磷钾用量下春玉米生物产量及其组分动态与养分吸收模式研究[J].作物学报, 1998, 4(2): 123-130.
He P, Jin J Y, Lin Betal. Dynamics of biomass and its components and models of nutrients absorption by spring maize under different nitrogen, phosphorous and potassium application rates[J]. Acta Agronomica Sinica, 1998, 4(2): 123-130.
[7] Scheer C, Wassmann R, Kienzler Ketal. Nitrous oxide emissions from fertilized, irrigated cotton (GossipyumhirsutumL.) in the Aral Sea Basin, Uzbekistan: Influence of nitrogen applications and irrigation practices[J]. Soil Biology and Biochemistry, 2008, 40: 290-301.
[8] 陆扣萍, 谢寅峰, 闵炬, 等. 不同施氮量对大棚莴苣根系形态及产量和品质的影响[J]. 土壤, 2011, 43(4): 542-547.
Lu K P, Xie Y F, Min Jetal. Effects of different nitrogen rates on root morphology, yield and quality of lettuce cultivated in plastic greenhouse[J]. Soil, 2011, 43(4): 542-547.
[9] 马溶慧, 许乃银, 张传喜, 等. 氮素调控棉花纤维蔗糖代谢及纤维比强度的生理机制[J]. 作物学报, 2008, 34(12): 2143-2151.
Ma R H, Xu N Y, Zhang C Xetal. Physiological mechanism of sucrose metabolism in cotton fiber and fiber strength regulated by nitrogen[J]. Acta Agronomica Sinica, 2008, 34(12): 2143-2151.
[10] 薛晓萍, 周治国, 张丽娟, 等. 棉花花后临界氮浓度稀释模型的建立及在施氮量调控中的应用[J]. 生态学报, 2006, 26(6): 1781-1791.
Xue X P, Zhou Z G, Zhang L Jetal. Development and application of critical nitrogen concentration dilution model for cotton after flowering[J]. Acta Ecology Sinica, 2006, 26(6): 1781-1791.
[11] 张炎, 侯秀玲, 王晓静, 等. 优化氮肥管理对膜下滴灌海岛棉产量和品质的影响[J]. 西北农业学报, 2006, 15(4): 122-125.
Zhang Y, Hou X L, Wang X Jetal. Effect of optimized nitrogen management on Sea Island cotton yield and fiber quality under plastic mulch drip irrigation[J]. Journal of Northwest Agriculture, 2006, 15(4): 122-125.
[12] 陈亮, 杨国正, 祝珍珍, 等. 氮素用量对棉花产量和品质的影响[J]. 中国棉花, 2011, 38(4): 15-18.
Chen L L, Yang G Z, Zhu Z Zetal. Effects of nitrogen fertilizer dosage on cotton yield and quality[J]. China Cotton, 2011, 38(4): 15-18.
[13] Gayler S, Wang E, Priesack Eetal. Modeling biomass growth, N-uptake and physiological development of potato crop[J]. Geoderma, 2002, 105: 367-383.
[14] 李春燕, 封超年, 张容, 等. 密度、 氮素对优质弱筋小麦宁麦 9 号旗叶早衰的调控效应[J]. 麦类作物学报, 2005, 25(5): 60-64.
Li C Y, Feng C N, Zhang Retal. Effect of plant density and nitrogen on flag leaf senescence of weak gluten wheat Ningmai 9 with good grain quality[J]. Journal of Triticeae Crops, 2005, 25(5): 60-64.
[15] 王之杰, 郭天财, 王化岑, 等. 种植密度对超高产小麦生育后期光合特性及产量的影响[J]. 麦类作物学报, 2001, 21(3): 64-67.
Wang Z J, Guo T C, Wang H Cetal. Effect of planting density on photosynthetic characteristics and grain yield of super high yield winter wheat at late growth stages[J]. Journal of Triticeae Crops, 2001, 21(3): 64-67.
[16] 吕新, 张伟, 曹连莆. 不同密度对新疆高产棉花冠层结构光合特性和产量形成的影响[J]. 西北农业学报, 2005, 14(1): 142-148.
Lü X, Zhang W, Cao L Petal. Effect of different density on canopy structure, photosynthesis and yield formation in high-yield cotton of Xinjiang[J]. Journal of Northwest Agriculture, 2005, 14(1): 142-148.
猜你喜欢
经济林研究(2022年4期)2023-01-12 09:14:46
中国纤检(2022年8期)2022-09-22 07:28:02
纺织标准与质量(2022年4期)2022-09-05 03:28:12
河北果树(2020年4期)2020-11-26 06:05:18
国际纺织导报(2016年6期)2016-09-18 02:17:43
新疆农垦科技(2016年2期)2016-08-21 13:50:18
中国棉花加工(2015年4期)2015-12-19 06:40:58
植物营养与肥料学报(2014年1期)2014-03-11 20:24:27
新疆农垦科技(2014年10期)2014-02-28 19:21:12
植物营养与肥料学报(2012年2期)2012-10-26 02:52:24
