
禽流感病毒特异性NP单克隆抗体的鉴定及禽流感特异性检测方法的建立
2014-04-02 08:44张旭辉柯少君沈晨光郭小怡康雅虹陈俊煜王明睿陈毅歆夏宁邵
中国人兽共患病学报 2014年8期
桂 勋,张旭辉,柯少君,李 睿,黄 彬,沈晨光,郭小怡,康雅虹,陈俊煜,王明睿,陈毅歆,,夏宁邵,
流感病毒属于正粘病毒科,单链负义的RNA病毒,根据其核蛋白(NP)及基质蛋白(M)抗原性的不同,分为甲(A)、乙(B)、丙(C)三型;根据其表面膜蛋白血凝素(HA)和神经氨酸酶(NA)的抗原性不同,分为16种H亚型和10种N亚型[1]。甲型流感病毒宿主广泛,能够感染禽、人、猪、马等动物,其中感染人类的主要有禽流感病毒和人流感病毒。人类甲型流感病毒主要有H1N1、H3N2和H2N2等3种,曾多次引起全球流感大流行[2-4]。禽流感病毒在自然界中广泛存在,目前已从禽类中分离到16种H亚型和10种N亚型的禽流感病毒[5]。禽流感病毒一般不能感染人类,但自1997年香港发生人感染禽流感H5N1病毒致死事件后,自然界中不断发生禽流感病毒突破种属屏障感染人类的事件,如禽流感病毒H5N1、H9N2、H7N7、H7N9、H10N8等[6-10],给人类健康和社会经济造成严重危害。更令人担忧的是,禽流感病毒与人流感病毒一旦发生基因重排,很容易形成能在人群中稳定传播的高致病性流感病毒株,造成全球流感大流行。因此,建立一种特异识别禽流感病毒的检测工具,有助于及时发现禽流感病毒,对禽流感病毒进行源头控制,防止禽流感疫情的扩散传播,防止发生禽流感病毒与人流感病毒的交叉感染事件,对降低禽流感的危害具有现实意义。目前国内外已有许多甲型流感病毒的抗原快速检测试剂得到应用[11],这些试剂多是特异识别甲型流感病毒NP蛋白,一般不能区分鉴别人流感病毒和禽流感病毒。鉴于禽流感病毒NP与人流感病毒NP存在抗原性差异,本研究拟通过筛选能够鉴别区分人流感NP和禽流感NP的单抗,进而建立一种能特异检测禽流感病毒但不识别人流感病毒的检测方法,为禽流感病毒的特异性诊断提供一种可行的检测工具。
1 材料与方法
1.1细胞、病毒与实验动物 小鼠骨髓瘤细胞株Sp2/0-Ag14(Sp2/0)以及MDCK细胞均为本中心保存。部分流感病毒株由香港大学微生物系陈鸿霖博士惠赠,部分流感病毒为本中心分离获得。BALB/c小鼠(6 ~ 8周龄)购自上海斯莱克实验动物有限公司。
1.2主要试剂及耗材 PEG1350、次黄嘌呤、胸腺嘧啶、氨基喋呤、DMSO、HRP、显色底物、FITC标记羊抗鼠二抗及DAPI均为Sigma公司产品。RPMI1640以及DMEM基础培养基为Gibco公司产品。胎牛血清为Hyclone公司产品。检测标HRP的兔多抗所用到的羊抗兔IgG由北京万泰生物药业有限公司提供。各种常规化学试剂为国产分析纯。硝酸纤维素膜为MDI公司的产品。渗滤装置由本中心完全自主设计开发[12]。
1.3NP基因进化树的制作 NP基因进化树采用软件MEGA5.0制作。
1.4NP重组蛋白的表达 首先,从NP进化树上选取禽源病毒分支上的代表株A/HK/212/03(H5N1,HK/212)及人源病毒分支上的代表株A/California/04/2009(H1N1,CA/04),利用RT-PCR的方法从病毒中调取NP基因全长序列,并进行测序鉴定。然后,采用常规分子克隆的方法分别构建克隆pET-28a/NP-HK/212及pET-28a/NP-CA/04,在E.coli菌株BL21中进行大量表达,取上清进行Ni2+柱亲和层析纯化,纯化后的NP重组蛋白的纯度采用10% SDS-PAGE做鉴定。
1.5NP单抗的制备 以50 μg rNP-HK/212稀释至250 μL,再与等量的福氏完全佐剂混匀,经皮下注射免疫BALB/c小鼠,剂量为500 μL/次。初免后第15 d和29 d,分别用同样剂量的重组蛋白加福氏不完全佐剂进行加强免疫。在融合前3 d,再以20 μg重组蛋白经脾脏注射做最后的加强免疫。细胞融合、细胞克隆化、腹水诱导及纯化均按常规方法进行,融合杂交瘤细胞株的筛选使用基于rNP-HK/212的ELISA进行检测。
1.6NP单抗的免疫荧光检测 利用4%的多聚甲醛固定禽流感病毒A/Teal/Hong Kong/W312/97(H6N1)和人流感病毒A/California/04/2009 (H1N1)感染的MDCK细胞,经0.03% TritonX-100通透后,加入山羊血清进行封闭1 h。加入相应的检测抗体反应1 h,PBS洗涤3次,再加入羊抗鼠二抗GAM-FITC反应40 min。经PBS洗涤3次后,加入DAPI染核5 min,用PBS洗涤5次后进行封片镜检。
1.7Dot-ELISA 首先将高纯度的NP单抗(如1F2及19C10)按5 μg/孔包被于硝酸纤维素膜的样品检测区,同时包被200 ng的羊抗兔IgG于对照区,25 ℃干燥后真空封装4 ℃保存备用。检测步骤为:①样品处理:200 μL样品与400 μL裂解液充分混匀;②加样:将上述处理后的样品全部加入样品检测区,待样品完全渗入后,摘掉流控装置;③加入酶标二抗:滴加200 μL酶标抗体(HRP标记的兔抗NP蛋白多克隆抗体),待样品完全渗入后,静置2 min;④洗涤:用洗涤液洗涤2次;⑤显色:待洗液渗入后,加入100 μL显色液,显色3 min;⑥终止读值:加入100 μL终止液,在5 min以内判定结果。
2 结 果
2.1甲型流感病毒NP基因的进化分析 首先,利用软件MEGA5.0对从GenBank上选取的不同分离时间、不同分离地点的甲型流感病毒NP基因进行了进化分析。结果显示,禽流感病毒与人流感病毒处在两个完全不同的分支上,禽流感病毒用红色标注,人流感病毒用蓝色标注(图1),说明禽流感NP和人流感病毒NP的抗原性可能存在不同。在此基础上,分别选取了禽流感病毒分支上的代表株A/HK/212/03 (H5N1)(黑点标注)及人流感病毒分支上的代表株A/California/04/2009 (H1N1)(红点标注)用于后续NP重组蛋白的表达、单抗制备及鉴定。
2.2NP重组蛋白的表达及纯化 采用RT-PCR的方法从A/HK/212/03及A/California/04/2009中调取了NP基因,经测序鉴定正确后,分别构建了pET-28a/NP-HK/212及pET-28a/NP-CA/04两种重组表达质粒。在E.coli菌株BL21(DE3)中进行大量表达后,利用亲和层析的方法纯化制备出高纯度的禽流感病毒NP抗原rNP-HK/212和人流感病毒NP抗原rNP-CA/04(图2)。
2.3NP单克隆抗体的制备 利用获得的rNP-HK/212重组蛋白免疫BALB/c小鼠,在免疫3针后选取血清滴度最高的两只小鼠进行了细胞融合实验及单抗筛选,共制备出抗rNP-HK/212单抗53株,并利用Protein A亲和层析的方式获得了高纯度的单抗。
2.4禽流感病毒NP蛋白特异性单抗的鉴定 利用间接ELISA的方法分别检测了上述53株单抗对rNP-HK/212及rNP-CA/04两种重组NP蛋白的反应性。根据单抗与两类NP抗原的反应性不同,可以将上述53株单抗分成三类(Class Ⅰ,Class Ⅱ和Class Ⅲ),其中Class Ⅰ类单抗有45株,该类单抗对两种NP重组蛋白均有很好的反应性;Class Ⅱ类单抗有3株,该类单抗对禽流感病毒NP重组蛋白rNP-HK/212有很好的反应性,但对人流感病毒NP重组蛋白rNP-CA/04不反应;Class Ⅲ类单抗对人流感病毒NP重组蛋白rNP-CA/04有很好的反应性,对禽流感病毒NP重组蛋白rNP-HK/212的反应性明显减弱(图3)。分别选取Class Ⅰ类单抗代表株19C10及Class Ⅱ 类单抗代表株1F2作为包被抗体检测其对禽流感病毒及人流感病毒的免疫荧光活性。结果显示,Class Ⅰ类单抗19C10对禽流感病毒A/Teal/Hong Kong/W312/97 (H6N1)及人流感病毒A/California/04/2009 (H1N1)均有很好的反应性;Class Ⅱ类单抗1F2对禽流感病毒有很好的反应性,但对人流感病毒不反应(图4)。说明Class Ⅱ类单抗能够特异识别禽流感病毒NP蛋白。
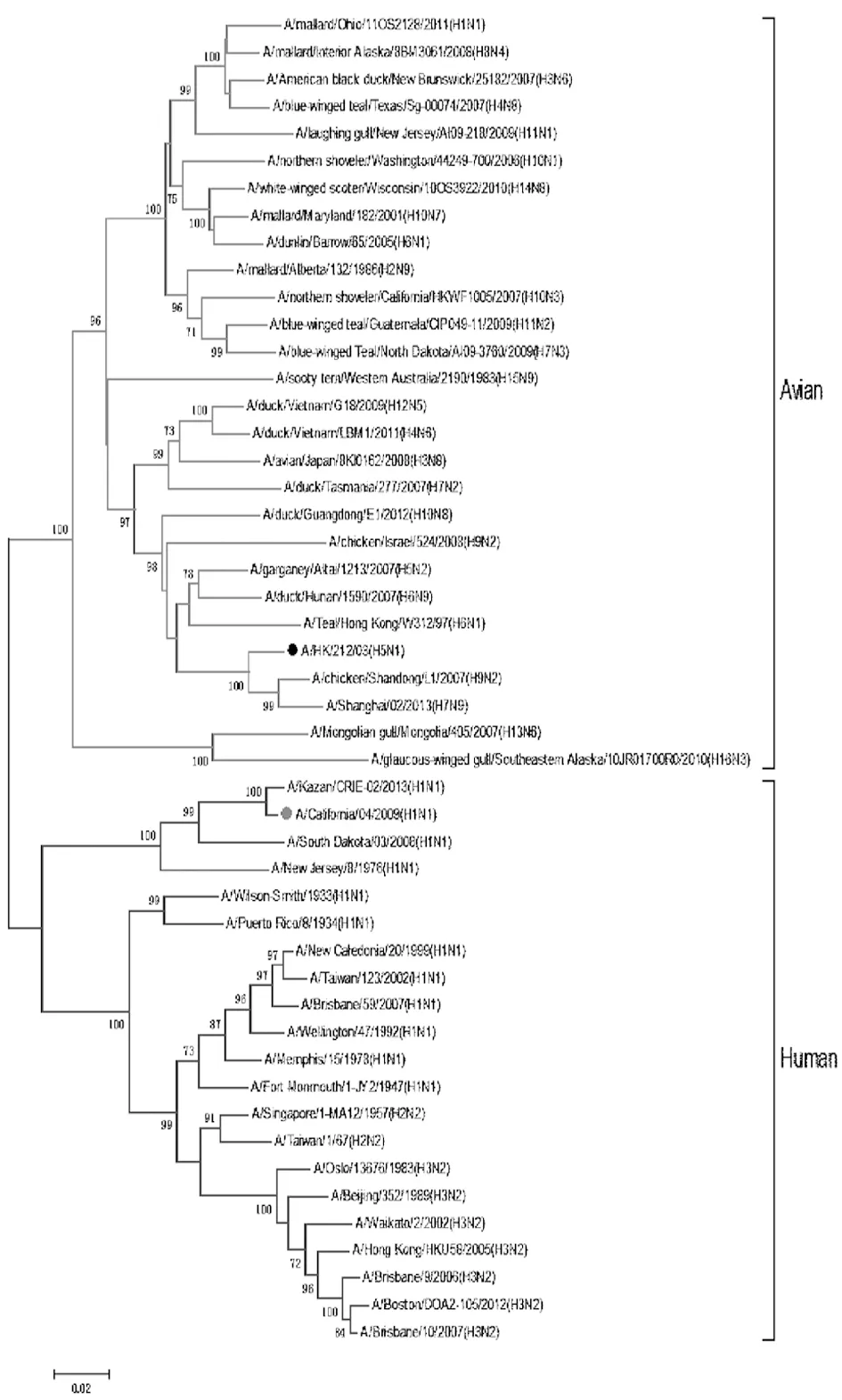
图1甲型流感病毒NP基因的进化分析
Fig.1PhylogenetictreebuiltwithNPsequencesofinfluenzaAvirus
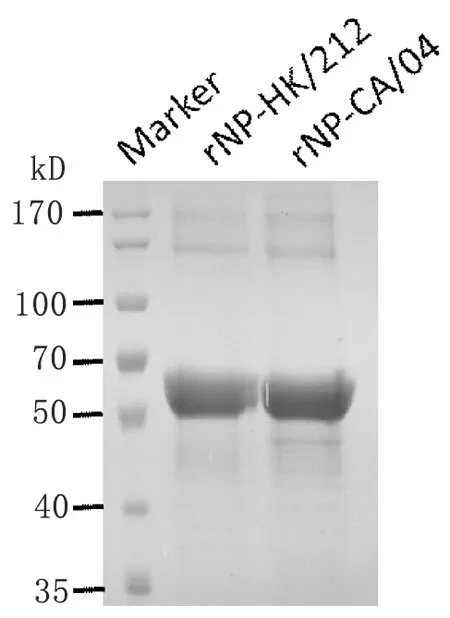
图2NP重组蛋白的制备
Fig.2PreparationofrecombinantNPantigensrNP-HK/212andrNP-CA/04

图3 NP单抗的分类鉴定结果
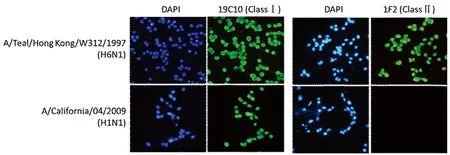
图4 两类NP单抗的免疫荧光检测结果
2.5禽流感病毒特异性Dot-ELISA检测方法的建立 本中心已研制出了多种酶联免疫渗滤(Dot-ELISA)检测试剂[12-13]。本研究在成熟的Dot-ELISA技术基础上,以禽流感病毒NP蛋白特异性单抗1F2作为包被抗体,以辣根过氧化物酶标记的NP兔多抗作为标记抗体,建立一种特异检测禽流感病毒的检测试剂AIV-Dot-ELISA。为评估该方法的检测效果,用19个不同亚型的禽流感病毒、8个不同亚型的人流感病毒和2个乙型流感病毒进行评估(表1),发现以Class Ⅱ 类单抗1F2为包被单抗的检测方法对19个禽流感病毒全部能够检出,其检测分析灵敏度约为0.025 ~ 0.1 HA titer,其对病毒浓度为5 HA titer 的8个人流感病毒和2个乙型流感病毒的检测均为阴性。而采用Class Ⅰ类NP广谱单抗19C10作为包被抗体的对照试剂对所有的甲型流感病毒均能检出,不能鉴别区分禽源流感病毒和人源流感病毒。上述结果说明本研究建立的禽流感特异性检测方法是有效可行的,下一步还需评估其对禽类拭子标本和人类拭子标本的检测效果。
3 讨 论
本研究利用分子进化分析选择了一个禽流感病毒NP基因和一个人流感病毒NP基因进行原核表达,获得两个高纯度的重组NP抗原,进而应用于NP单抗的筛选,获得3类NP特异性单抗,其中以1F2为代表的NP单抗只特异识别禽流感病毒而不识别人流感病毒,为构建特异识别禽流感病毒的检测方法奠定了基础。在此基础上,基于Dot-ELISA抗原快速检测技术平台,用禽流感病毒NP特异性单抗1F2作为包被抗体,成功建立起一个只特异识别禽流感病毒而不识别人流感病毒的适于禽类流感检测的抗原快速检测方法AIV-Dot-ELISA。鉴于近年来不同类型的禽流感病毒(H5N1、H9N2、H7N7、H7N9和H10N8)频频感染人类[6-8],而当前的甲型流感病毒快速检测试剂不能够鉴别区分禽流感病毒和人流感病毒的感染,因此,本研究建立的方法将有助于快速鉴别禽流感病毒的存在,将为禽流感疫情控制和人感染禽流感病毒鉴别诊断提供一种可行的分析检测工具。

表1 AIV-Dot-ELISA检测试剂对禽流感病毒及人流感病毒的检测结果
Note: +,Positive; -,Negative;
本研究获得的NP单抗可分成3类,Class Ⅰ类NP广谱单抗,对禽流感病毒和人流感病毒的识别能力相当;Class Ⅲ类NP单抗,只识别禽流感病毒而不识别人流感病毒;Class Ⅱ类单抗,对人流感病毒的识别活性显著高于对禽流感病毒的识别活性。这3类单抗存在的不同反应性特征说明流感病毒NP的抗原性存在宿主特异性,不同宿主来源的NP基因可能因为序列不同,存在不同的抗原性表位,这样的独特性表位可以被特异性单抗所鉴别区分,从而可被用来建立宿主特异性检测方法。下一步工作除了进一步利用临床采集标本完善评估该方法的检测效果外,还有必要利用表位分析方法研究上述3类单抗识别表位的特征区别,从而发现流感病毒中与宿主特异性相关的关键表位氨基酸及其在流感病毒生物学中的潜在功能。另外,本研究将为建立其他种属特异性流感病毒检测方法或亚型特异性流感病毒检测方法提供一种可供借鉴的研究经验。
参考文献:
[1]Peiris JS,de Jong MD,Guan Y. Avian influenza virus (H5N1): a threat to human health[J]. Clin Microbiol Rev,2007,20(2): 243-267. DOI: 10.1128/CMR.00037-06
[2]Patterson KD,Pyle GF. The geography and mortality of the 1918 influenza pandemic[J]. Bull Hist Med,1991,65(1): 4-21. DOI: 10.1353/bhm.2002.0022.
[3]Fraser C,Donnelly CA,Cauchemez S,et al. Pandemic potential of a strain of influenza A (H1N1): early findings[J]. Science,2009,324(5934): 1557-1561. DOI: 10.1126/science.1176062
[4]Muscatello DJ,Cretikos MA,Macintyre CR. All-cause mortality during first wave of pandemic (H1N1) 2009,New South Wales,Australia,2009[J]. Emerg Infect Dis,2010,16(9): 1396-1402. DOI: 10.3201/eid1609.091723
[5]Alexander DJ. A review of avian influenza in different bird species[J]. Vet Microbiol,2000,74(1-2): 3-13. DOI: 10.1016/S0378-1135(00)00160-7
[6]Li KS,Guan Y,Wang J,et al. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia[J]. Nature,2004,430(6996): 209-213. DOI: 10.1038/nature02746
[7]Gao R,Cao B,Hu Y,et al. Human infection with a novel avian-origin influenza A (H7N9) virus[J]. N Engl J Med,2013,368(20): 1888-1897. DOI: 10.1056/NEJMoa1304459
[8]Butt KM,Smith GJ,Chen H,et al. Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003[J]. J Clin Microbiol,2005,43(11): 5760-5767. DOI: 10.1128/JCM.43.11.5760-5767.2005
[9]Du Ry van Beest Holle M,Meijer A,Koopmans M,et al. Human-to-human transmission of avian influenza A/H7N7,The Netherlands,2003[J]. Euro Surveill,2005,10(12): 264-268.
[10]Parry J. H10N8 avian flu virus claims its first known human casualty[J]. BMJ,2014,348: g1360. DOI: 10.1136/bmj.g1360
[11]Reina J,Padilla E,Alonso F,et al. Evaluation of a new dot blot enzyme immunoassay (directigen flu A+B) for simultaneous and differential detection of influenza a and B virus antigens from respiratory samples[J]. J Clin Microbiol,2002,40(9): 3515-35157. DOI: 10.1128/JCM.40.9.3515-3517.2002
[12]Chen Y,Xu F,Fan X,et al. Evaluation of a rapid test for detection of H5N1 avian influenza virus[J]. J Virol Methods,2008,154(1-2): 213-215. DOI: 10.1016/j.jviromet.2008.08.013
[13]Chen Y,Xu F,Gui X,et al. A rapid test for the detection of influenza A virus including pandemic influenza A/H1N1 2009[J]. J Virol Methods,2010,167(1): 100-102. DOI: 10.1016/j.jviromet.2010.02.001
猜你喜欢
广东农业科学(2022年11期)2023-01-13
实用肿瘤学杂志(2022年3期)2022-11-30
医院管理论坛(2022年7期)2022-10-14
世界科学技术-中医药现代化(2021年12期)2021-04-19
皮肤性病诊疗学杂志(2020年4期)2020-09-02
科学24小时(2019年5期)2019-06-11
环球时报(2018-03-19)2018-03-19
猪业科学(2018年6期)2018-01-23
北方牧业(2016年15期)2016-12-17
小星星·阅读100分(高年级)(2014年6期)2014-06-27
