
日本沼虾幼虾对饲料中铜的需求量
2014-03-28 10:50:50郭建林陈建明沈斌乾孙丽慧姜建湖
动物营养学报 2014年6期
郭建林 陈建明 潘 茜 沈斌乾 孙丽慧 姜建湖
铜是血液中血蓝蛋白的重要组分[1],还是机体中细胞色素C氧化酶、赖氨酸氧化酶、超氧化物歧化酶等酶的组分,其参与机体一系列重要功能,如细胞呼吸、组织色素沉积、角化作用等,对机体营养物质代谢和生长发育具有重要的作用[2]。铜对虾蟹类等甲壳动物来说是非常重要的矿物元素,如果虾类在生长过程中铜摄入量不足,将导致生长缓慢,肝胰脏、甲壳、血液中铜含量降低以及心脏增大等症状,并影响其免疫机能[3-4],而且其在生长蜕壳过程中机体也会损失一部分的铜,如不及时补充就会影响生长和成活。但虾类摄入铜过多会对其生长和免疫功能造成负面影响[4],并因肝胰脏积累过量的铜而导致中毒[5],且过多的铜随机体排出体外会造成水体污染。因此,确定配合饲料中铜的适宜含量对于提高虾类饲料质量和改善养殖水域环境都有重要的意义。
日本沼虾(Macrobrachium nipponense)是我国传统的经济虾种,其人工养殖已成为我国淡水养殖中的重要产业之一,至2012年,其养殖产量达23.74 万 t[6]。但是,与养殖产业规模相比,对日本沼虾饲料营养的研究还相对滞后,现有的研究主要集中在蛋白质[7-9]、脂肪[9]、赖氨酸[10]、锌[11]等方面,尚无有关该虾饲料中铜需求量的研究。因此,本试验拟以日本沼虾幼虾为研究对象,开展其铜需求量的梯度试验,旨在为该虾的营养学研究提供基础性资料,并为生产实践提供理论依据和指导。
1 材料与方法
1.1 试验饲料
试验以酪蛋白(C5890,Sigma)、玉米淀粉、糊精、α-纤维素、鱼油、豆油等为原料制成基础饲料,其组成及营养水平见表1。以无水硫酸铜(分析纯,纯度为98.0%,购自国药集团化学试剂有限公司)为铜源,通过在基础饲料中添加 0、7、14、21、28、35、70 mg/kg 的铜,配制成铜含量分别为 0.63、6.70、12.64、19.15、25.64、31.99、65.27 mg/kg 的 7种试验饲料。试验饲料制作时,先将原料粉碎,使其全部过60目筛,再按配方比例充分混匀后,适量加水搅拌,通过绞肉机挤压成直径1.5 mm的长条,风干后破碎成粒径为1~2 mm的破碎料,置于4℃冰箱内密封保存备用。
1.2 试验动物与饲养管理
试验用日本沼虾幼虾由浙江省淡水水产研究所特种水产苗种基地提供,平均体重为(0.10±0.01)g,选择体质健壮、规格整齐的个体随机分组进行养殖试验。试验共分7组,每组3个重复,每个重复40尾虾。试验养殖系统为室内反复循环流水式玻璃缸养殖系统,以重复为单位放养于水族箱内,水族箱养殖水体为 0.9 m×0.5 m×0.4 m,水族箱内放置生物过滤装置和气石,并放置足够数量的网片作为试验虾的栖息隐蔽物。试验用水为经过曝气的自来水,日换水量约为总水体的1/3,连续充气。采用饱食投喂,投喂时间为08:00、16:00,在投喂1 h后吸取残饵并收集烘干,以计算摄食量。试验期间,养殖水体参数:温度22~30 ℃;pH 7.4~8.1;溶氧浓度 6.3~8.0 mg/L;氨氮浓度 0.01~0.05 mg/L;亚硝酸盐浓度 0.01~0.05 mg/L;硫化物浓度 0.02~0.05 mg/L;铜含量(0.5±0.2)μg/L。试验期为 49 d。

表1 基础饲料组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet(air-dry basis) %
1.3 生长性能的测定
饲养试验结束时,统计每个重复的饲料总摄食量,停食24 h后称虾体总重并记录虾尾数,计算增重率(WGR)、特定生长率(SGR)、成活率(SR)和饲料系数(FCR)。相关计算公式如下:
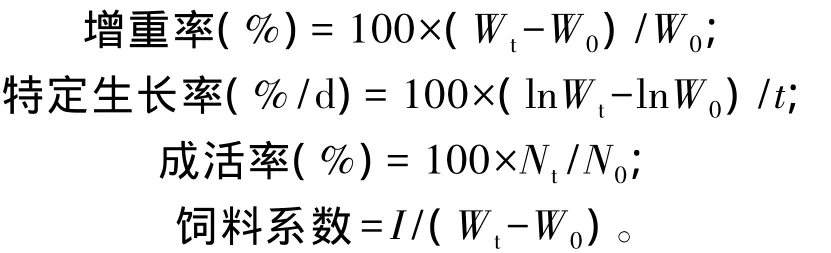
式中:W0为初始体重(g);Wt为终末体重(g);N0为初始尾数(尾);Nt为终末尾数(尾);t为养殖天数(d);I为饲料摄入量(g)。
1.4 样品采集与分析
从每箱里随机取22尾虾,其中10尾作为全虾样品,其余12尾采集血清后分离肝胰脏、甲壳和肌肉。水分含量采用105℃常压干燥法测定,粗蛋白质含量采用凯氏定氮法测定,粗脂肪含量采用索氏抽提法测定,粗灰分含量采用马福炉550℃灼烧法测定。上述指标测定步骤均参照AOAC(1995)[12]的方法进行。
用无菌注射器从虾围心腔取血,由于虾体血液较少,4尾虾体血液混合置于1个无菌的离心管中,于4℃过夜后,5 000 r/min(4℃)离心20 min后移出上清液即得血清。血清酚氧化酶(PO)活力的测定参照王雷等[13]的方法,铜锌超氧化物歧化酶(CuZn-SOD)活力采用黄嘌呤氧化酶-羟胺法测定,上述指标测定所用试剂盒由南京建成生物工程研究所生产。
肌肉、肝胰脏和甲壳中铜及全虾中铜、锌、铁含量采用原子吸收分光光度法[11]测定。将样品在105℃常压下干燥,粉碎后对样品进行预处理:称取0.2 g样品置于80 mL的特氟龙微波消解罐中,加入10 mL浓硝酸与1 mL浓盐酸,置于微波消解系统(ETHOS1,Milestone)中 150℃工作80 min,冷却至室温后取出微波消解罐,在150℃加热板上赶酸后,冷却定容至50 mL容量瓶中。用原子吸收分光光度计(AA240,Varian)测定溶液中铜、锌、铁含量后换算成组织中的含量。
1.5 数据处理
试验数据以平均值±标准差(mean±SD)表示,采用SPSS 11.5分析软件的one-way ANOVA程序对所得试验数据进行单因素方差分析,若有显著差异,再用Duncan氏法进行组间差异显著性的多重比较,P<0.05为差异显著。分别以增重率和甲壳中铜含量为评价指标,采用折线模型(brokenline model)分析日本沼虾幼虾对饲料中铜的需求量。
2 结果
2.1 饲料中铜含量对日本沼虾幼虾生长性能的影响
由表2可以看出,随着饲料中铜含量的升高,日本沼虾幼虾的增重率和特定生长率均呈先升高后降低的趋势,二者均在25.64 mg/kg组达到最高,除与 31.99 mg/kg 组无显著差异(P>0.05)外,显著高于其他各组(P<0.05)。各组日本沼虾幼虾的成活率在 68.19%~73.33%之间,通过方差分析,各组间无显著差异(P>0.05)。随着饲料中铜含量的升高,日本沼虾幼虾的饲料系数呈先降低后升高的趋势,以25.64 mg/kg组的饲料系数最低,显著低于 0.63、6.70、12.64 和 65.27 mg/kg 组(P<0.05),与这些组相比,25.64 mg/kg 组的饲料系数 分 别 降 低 了 22.86%、12.90%、8.78% 和10.00%。以增重率为评价指标,采用折线模型分析得出日本沼虾幼虾对饲料中铜的需求量为24.69 mg/kg(图 1)。
2.2 饲料铜含量对日本沼虾幼虾体成分的影响
由表3可以看出,日本沼虾全虾的水分、粗蛋白质、粗脂肪与粗灰分含量在各组之间均无显著差异(P>0.05)。
2.3 饲料中铜含量对日本沼虾幼虾血清PO和CuZn-SOD活力的影响
由表4可以看出,随着饲料中铜含量的升高,日本沼虾幼虾血清PO活力呈先上升后下降的趋势,以 31.99 mg/kg 组最高,显著高于 0.63、6.70、12.64 mg/kg 组(P <0.05)。饲 料 中 铜 含 量 在0.63~12.64 mg/kg 时,血清 CuZn-SOD 活力随着饲料中铜含量的升高而显著上升(P<0.05);饲料中 铜 含 量 在 12.64~31.99 mg/kg时,血 清CuZn-SOD活力出现了近似的平台期,其活力趋于稳定,12.64、19.15、26.54、31.99 mg/kg 组间无显著差异(P>0.05);而当饲料中铜含量继续增加至65.27 mg/kg时,血清CuZn-SOD活力则显著下降(P<0.05)。
2.4 饲料中铜含量对日本沼虾幼虾组织中铜、锌、铁含量的影响
由表5可以看出,饲料中铜含量达到19.15 mg/kg后再继续升高,全虾中铜含量显著上升(P<0.05),0.63、6.70 和 12.64 mg/kg 组全虾中铜含量较低,显著低于 31.99和 65.27 mg/kg组(P<0.05)。肌肉中铜含量随着饲料中铜含量的升高而 持 续 上 升,12.64、19.15、26.54、31.99 和63.70 mg/kg组肌肉中铜含量显著高于 0.63和6.70 mg/kg组(P<0.05),并在 63.70 mg/kg 组达到最大,此组肌肉中铜含量显著高于其他各组(P<0.05)。随着饲料中铜含量的升高,肝胰脏中铜含量也持续上升,尤其是在饲料中铜含量达到19.15 mg/kg后,肝胰脏中铜含量成倍上升,65.27 mg/kg组肝胰脏中铜含量分别是 0.63、6.70、12.64、19.15、25.64 和 31.99 mg/kg 组的 63.70、50.10、40.08、11.10、3.09 和 1.37 倍,差异显著(P<0.05);各 组 间 肝 胰 脏 中 铜 含 量 除 6.70和12.64 mg/kg组无显著差异(P>0.05)外,其他各组间差异显著(P<0.05)。饲料中铜含量在 0.63~12.64 mg/kg时,甲壳中铜含量随着饲料中铜含量的升高而上升,在饲料中铜含量达到19.15 mg/kg后,甲壳中铜含量趋于稳定。以日本沼虾幼虾甲壳中铜含量为评价指标,采用折线模型分析得出日本沼虾幼虾对饲料中铜的需求量为21.48 mg/kg(图 2)。
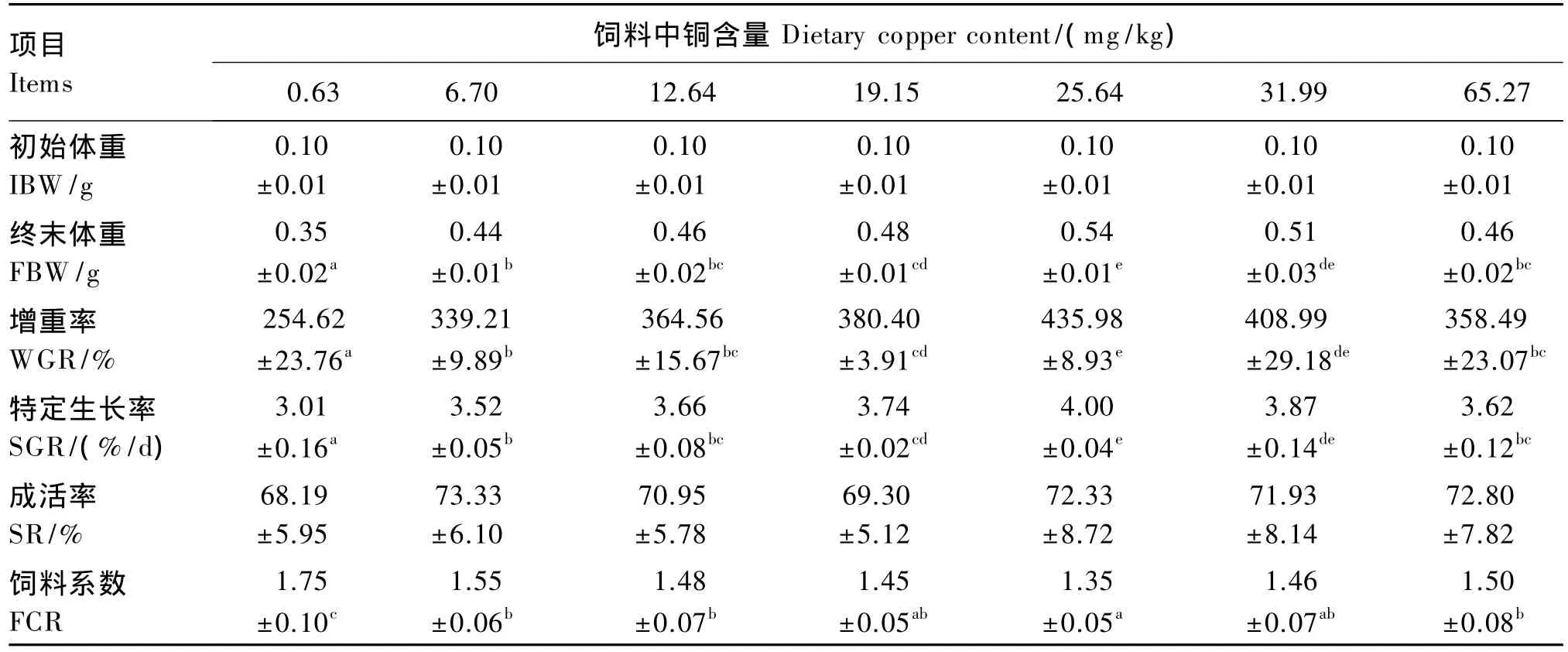
表2 饲料中铜含量对日本沼虾幼虾生长性能的影响Table 2 Effects of dietary copper content on growth performance of juvenile oriental river prawn
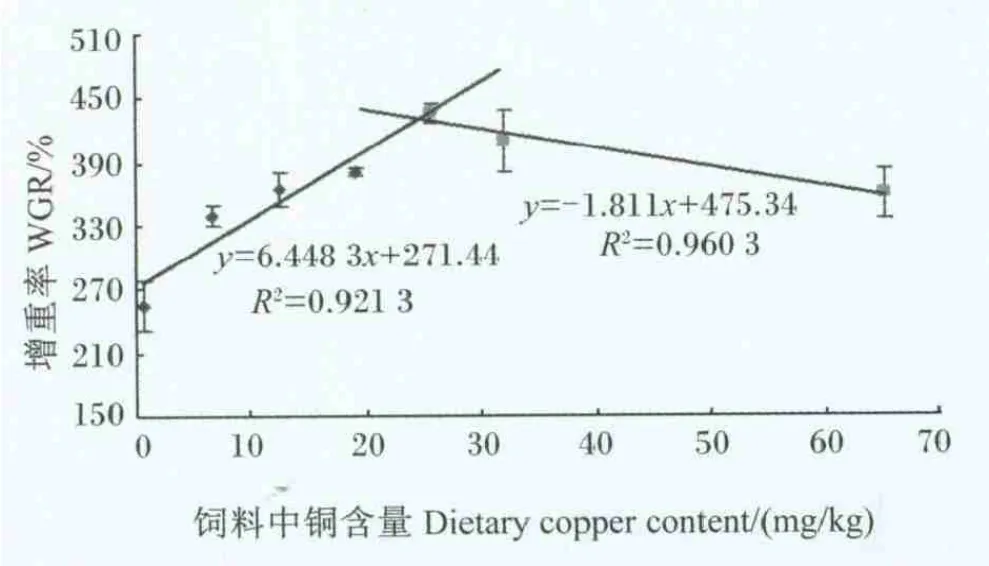
图1 饲料中铜含量与日本沼虾幼虾增重率的关系Fig.1 Relationship between dietary copper content and WGR of juvenile oriental river prawn
由表5可以看出,随着饲料中铜含量的升高,全虾中锌含量呈先平稳后下降的趋势,以0.63、6.70和12.64 mg/kg组全虾中锌含量较高,上述3组间差异不显著(P>0.05),但均显著高于其他各组(P<0.05),而 19.15 mg/kg 组全虾中锌含量则显著高于 26.54、31.99 和 63.70 mg/kg 组(P<0.05),后 3组全虾中锌含量无显著差异(P>0.05)。饲料中铜含量也影响着全虾中铁含量,随着饲料中铜含量的升高,全虾中铁含量呈阶段式下降的趋势。0.63 mg/kg组全虾中铁含量显著高于其他各组(P<0.05),其铁含量是其他组的1.62~2.38 倍,6.70、12.64 和 19.15 mg/kg 组之间全虾中铁含量无显著差异(P>0.05),但均显著高于 26.54、31.99 和 63.70 mg/kg 组(P<0.05)。
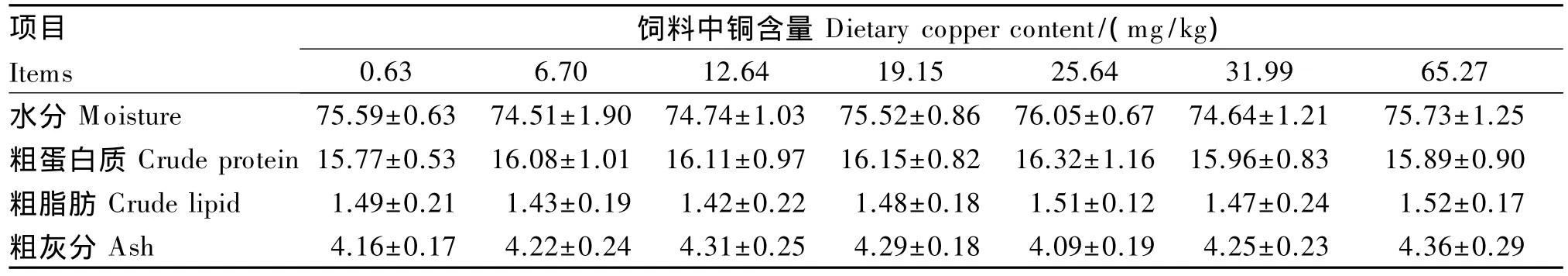
表3 饲料中铜含量对日本沼虾幼虾体成分的影响(鲜重基础)Table 3 Effects of dietary copper content on body composition of juvenile oriental river prawn(fresh weight basis)%
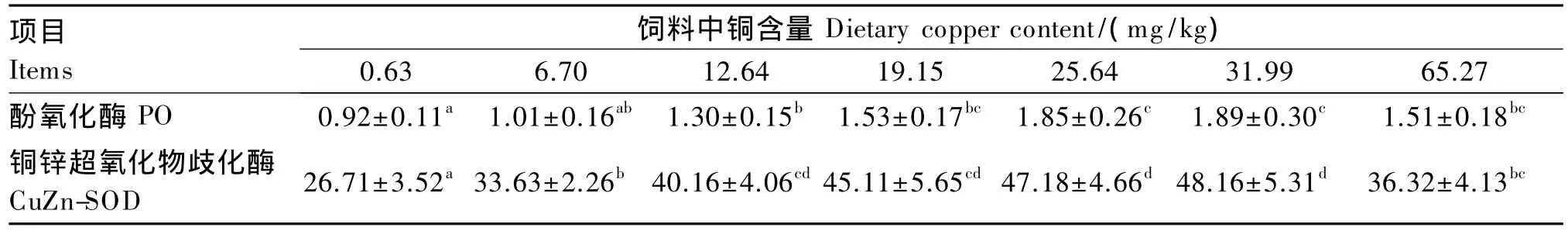
表4 饲料中铜含量对日本沼虾幼虾血清PO和CuZn-SOD活力的影响Table 4 Effects of dietary copper content on PO and CuZn-SOD activities in serum of juvenile oriental river prawn U/mL
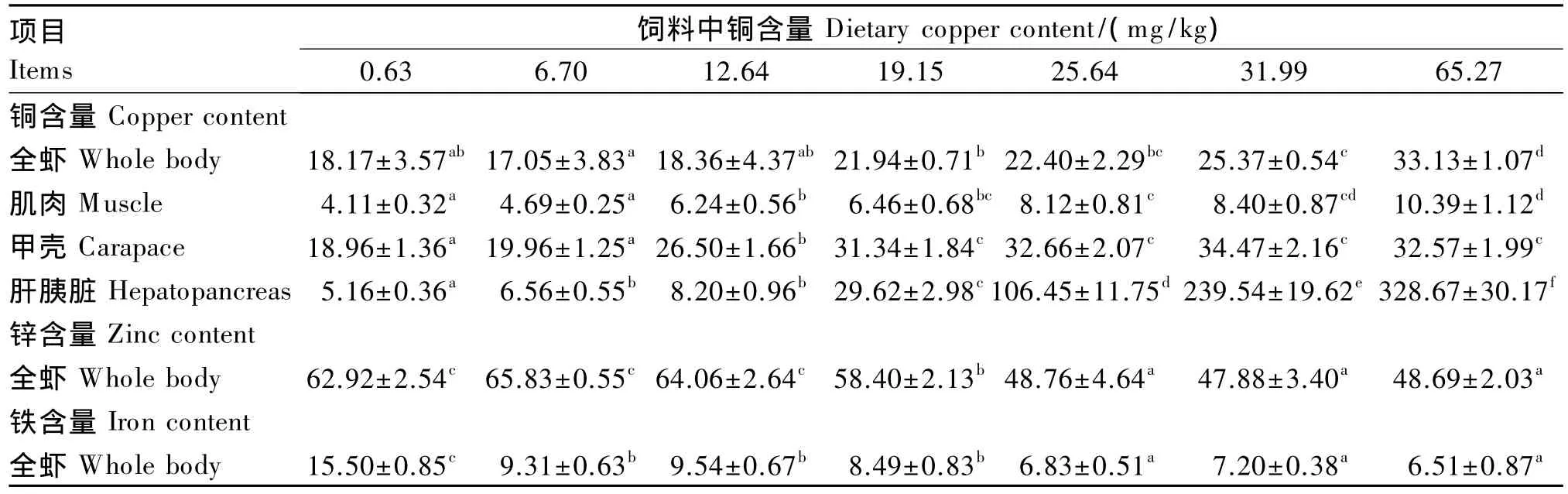
表5 饲料中铜含量对日本沼虾幼虾组织中铜、锌、铁含量的影响Table 5 Effects of dietary copper content on copper,zinc and iron contents in tissues of juvenile oriental river prawn mg/kg
3 讨论
3.1 饲料中铜含量对日本沼虾幼虾生长性能的影响
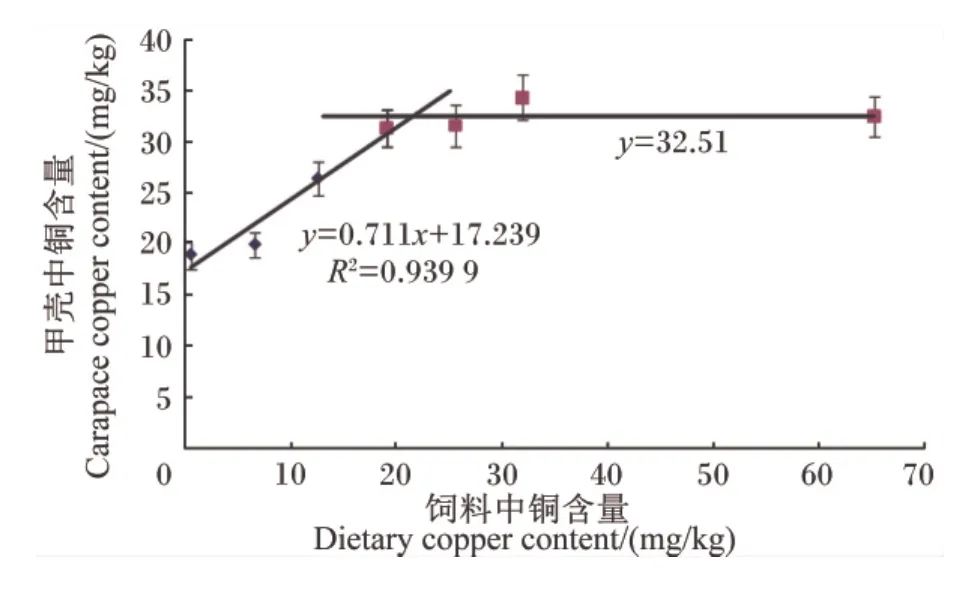
图2 饲料中铜含量与日本沼虾幼虾甲壳中铜含量的关系Fig.2 Relationship of dietary copper content and carapace copper content of juvenile oriental river prawn
在本试验中,不同的铜添加量可以不同程度地提高日本沼虾幼虾的生长性能,生长最慢的添加组(添加7 mg/kg铜的组)的增重率也比对照组(未添加铜的组)高出了33.22%,证明了铜对日本沼虾来说是一种重要的矿物元素。研究普遍认为水产动物对铜的需求有一个适宜的剂量范围,饲料中铜含量过低或过高均可能会引起生长抑制,使成活率降低[4,14]。本试验的结果也证实了这一点,当饲料中铜含量低于最佳生长的需求量时,日本沼虾幼虾的生长均受到了不同程度地抑制,这在对凡纳滨对虾(Litopenaeus vannamei)[3]、斑节对虾(Penaeus monodon)[4]、中华绒螯蟹(Eriocheir sinensis)[15]等的研究中也出现了相似的结果;而当饲料中铜含量高于最佳生长的需求量时,对日本沼虾的生长产生了负面影响,生长速度明显下降,这可能是因为高铜抑制了细胞色素氧化酶的活性,进而影响了虾体的生长[5]。Lee 等[4]对斑节对虾的研究也表明,饲料中铜含量高于40 mg/kg时,斑节对虾出现生长受阻,增重率下降;同样类似的结果在凡纳滨对虾[3]和中华绒螯蟹[15]的相关研究中也得到了证实。饲料系数是评定饲料利用率的重要指标。本试验中,随着饲料中铜含量的升高,饲料系数呈先下降后上升的趋势,以26.54 mg/kg组的饲料系数最低,这与增重率的结果相一致,与周萌等[16]、宋维彦等[17]的研究结果相似,说明饲料中适宜含量的铜能提高虾体对饲料的利用率,降低饲料系数。
增重率在评价水产动物矿物元素需求中具有重要的作用[3-4,11,15],因此,根据本试验结果,对日本沼虾幼虾的增重率和饲料中铜含量作了回归分析,结果表明,日本沼虾幼虾获得最佳增重率时对饲料中铜的需求量为 24.69 mg/kg。王维娜等[18]的研究表明,当饲料中铜含量为25.27 mg/kg时,中国对虾(Fenneropenaeus chinensis)具有最大的生长量;Lee等[4]对斑节对虾铜需求量的研究表明,在饲料中铜含量为21.3 mg/kg时,斑节对虾具有最大的增重;Davis等[3]在对凡纳滨对虾的研究中发现,饲料中添加32 mg/kg铜(铜总含量为34 mg/kg)时,凡纳滨对虾的增重达到最大;郭志勋等[19]的研究表明,在饲料中添加 30 mg/kg铜时,南美白对虾(亦称凡纳滨对虾)的生长速度最快;Sun 等[15]的研究表明,饲料中铜含量为20.78~40.24 mg/kg时,中华绒螯蟹生长速度较快。与上述研究结果相比,本研究得出的日本沼虾幼虾对饲料中铜的需求量与中国对虾接近,略高于斑节对虾,但低于凡纳滨对虾。这可能与不同种类虾对铜的需求量不同相关,也与试验研究使用的配方有差异相关,若配方中的植物原料(植酸)含量较高,植酸会与铜等矿物元素螯合,从而影响机体对矿物元素的利用率。
3.2 饲料中铜含量对日本沼虾幼虾免疫力的影响
PO是虾蟹类等甲壳动物酚氧化酶原激活(proPO)系统的产物,在甲壳动物免疫过程中发挥着重要作用,是甲壳动物最重要的免疫酶之一[20-22]。本研究发现,饲料中适宜的铜含量(19.15~31.99 mg/kg)能显著提高血清 PO 活力,但饲料中铜含量超过31.99 mg/kg时,血清PO活力下降。郭志勋等[19]对凡纳滨对虾的研究也表明,在饲料中添加30 mg/kg铜时血清PO活力最高,当铜添加量超过30 mg/kg时,血清PO活力则开始下降;董晓慧等[23]的研究也发现,凡纳滨对虾的血清PO活力随着饲料中铜添加量的增加而上升,在添加量为30 mg/kg时达到最大;Sun等[15]对中华绒螯蟹的研究也表明,血清PO活力随着饲料中铜含量的升高呈先上升后下降的趋势,在20.78~40.34 mg/kg时活力较大。这表明饲料中适宜的铜含量能促进甲壳动物血清PO活力的提高,从而增强免疫力。铜是抗氧化酶CuZn-SOD活性中心的重要组成成分[24],因此CuZn-SOD活力被认为是机体内铜含量的一个敏感标志[25-28]。Sun等[15]在对中华绒螯蟹的研究中发现,饲喂对照组饲料时中华绒螯蟹的血清CuZn-SOD活力较低,而在饲料中补充铜后显著提高了血清CuZn-SOD活力,但当饲料中铜含量超过40.34 mg/kg时,血清CuZn-SOD活力开始下降。本试验结果与上述研究结果相似,日本沼虾幼虾血清CuZn-SOD活力随着饲料中铜含量的增加呈先上升再平稳后下降的趋势,在 19.15~31.99 mg/kg组中其活力较强,这表明饲料中适宜的铜含量有利于日本沼虾幼虾血清CuZn-SOD活力的提升,进而提高虾体的抗氧化力和免疫力。本试验中,血清PO和CuZn-SOD活力与增重率的结果相一致,说明日本沼虾摄食适宜铜含量的饲料不但可获得最佳的生长,而且可增强机体的免疫力,这与在凡纳滨对虾[19,23]和中华绒螯蟹[15]上的研究结果相似。
3.3 饲料中铜含量对日本沼虾幼虾组织中铜、锌、铁含量的影响
在对矿物元素的研究中,机体组织中矿物元素的含量是评定其需求量的重要指标[11,29-30]。本试验中,全虾中铜含量随着饲料中铜含量的升高而上升,甲壳中铜含量也随着饲料中铜含量的升高而上升,但趋势与全虾不同,当饲料中铜含量低于19.15 mg/kg时,甲壳中铜含量显著上升,当饲料中铜含量达到和超过19.15 mg/kg后,甲壳中铜含量趋于平稳,出现一个近似平台期。这可能是因为日本沼虾的代谢机制在满足甲壳铜沉积量后,饲料中过剩的铜被肝胰脏等其他组织储存或者代谢掉,在短时间内饲料中铜含量的升高还不足以引起日本沼虾甲壳中铜含量的上升,也许只有当饲料中铜含量非常高时(破坏虾体的代谢机能)才会打破这个平台期。铜是参与纤维化的赖氨酰氧化酶等酶的辅助因子,对维持正常骨骼强度具有重要的意义[2]。甲壳作为虾蟹类的外骨骼,其作用与脊椎动物的骨骼有相似,因此甲壳中铜含量可以作为评价日本沼虾幼虾对饲料中铜的需求量的有效指标。通过折线模型分析得出,饲料中铜含量为21.48 mg/kg时,日本沼虾幼虾甲壳中铜含量达到饱和。虾类对铜的积累主要在肝胰脏中,仅有一小部分铜被传递到肌肉中[31],这一点在本试验中得到了证实,虽然饲料中铜含量影响着日本沼虾幼虾肌肉中铜含量,但是其含量远远小于肝胰腺中铜含量,尤其是在铜含量达到日本沼虾幼虾营养需求量后,肝胰脏中铜含量是肌肉的 13.11 ~ 31.63 倍,这在凡纳滨对虾[3,14,19,23]、中国对虾[18]的相关研究中也得到类似的结果。这可能是肌肉和肝胰脏在机体代谢活动中所发挥的生理功能不同造成的,肝胰脏主要是解毒及代谢外来异物,而肌肉的主要功能是运动;也可能是肌肉的代谢机制使其铜含量可自我平衡调节,使肌肉中铜的积累受到控制[31],保持相对稳定性。魏万权等[32]的研究也指出,肝胰脏对铜的高度积累能力是某种预防机制,可能具有如下生理功能:当铜摄入量不足时,肝胰脏中库存的铜释放出来,满足动物机体代谢需要;当铜摄入过量时,铜可以在肝胰脏中积累而降低其对机体的毒性。
矿物元素间的关系是研究水产动物矿物元素营养需求的重要内容[11,29-30,33]。矿物元素之间相互影响,存在着拮抗和协同的关系,如铜和锌存在着拮抗关系[11,34-37],饲料中铜含量过高会影响锌的吸收和利用,其机制在于铜和锌在机体内会共同竞争结合金属硫蛋白,任何一方过多都会抑制另一方与金属硫蛋白的结合,影响机体对其的吸收利用[38]。本试验结果也表明,随着饲料中铜含量的升高,日本沼虾幼虾全虾中锌含量均呈先平稳后下降的趋势。王维娜等[18]对中国对虾的研究也证实了这一点,即中国对虾的头胸部锌含量随着饲料中铜含量的升高而下降。郭建林等[11]的研究发现,高含量锌抑制了日本沼虾对饲料中铜的吸收,全虾、甲壳和肝胰脏中铜含量显著降低。因此,在设计虾饲料配方时,要考虑铜、锌的交互作用,尽量避免两者产生拮抗。饲料中适量的铜可促进动物机体对铁的吸收利用,但过量的铜会抑制铁的吸收利用[39]。王维娜等[18]研究认为,随着饲料中铜含量的升高,中国对虾头胸部铁含量呈先保持稳定后显著下降的趋势。本试验结果表明,全虾中铁含量随着饲料中铜含量的增加呈阶段式下降的趋势。现有的对水产动物铜与其他矿物元素间相互作用的报道较少,因此,有必要进一步开展对水产动物铜与锌、铜与铁以及铜与其他主要矿物元素交互作用的研究,如一种矿物质的正常代谢需要另一种矿物质的参与、运输或储存及分子结合位点的竞争、酶活性位点的替代等。
4 结论
①在本试验条件下,饲料中适宜的铜含量能提高日本沼虾幼虾的生长性能和免疫力,且饲料中铜含量影响着组织中铜、锌、铁含量。
②以增重率和甲壳中铜含量为评价指标,通过折线模型分析得出日本沼虾幼虾对饲料中铜的需求量分别为 24.69 和 21.48 mg/kg。
[1] OWEN C A,Jr.Biochemical Aspects of copper:copper proteins,ceruloplasmin,and copper protein binding[M].Park Ridge:Noyes Publications,1982:41-44.
[2] O’DELL B L.Biochemistry of copper[J].Medical Clinics of North America,1976,60(4):687-703.
[3] DAVIS D A,LAWRENCE A L,GATLIN D M.Dietary copper requirement of Penaeus vannamei[J].Nippon Suisan Gakkaishi,1993,59(1):117-122.
[4] LEE M H,SHIAU S Y.Dietary copper requirement of juvenile grass shrimp,Penaeus monodon,and effects on non-specific immune response[J].Fish and Shellfish Immunol,2002,13(4):259-270.
[5] 刘发义,梁德海,孙凤,等.饵料中的铜对中国对虾的影响[J].海洋与湖沼,1990,21(5):404-410.
[6] 农业部渔业局.2013中国渔业统计年鉴[M].北京:中国农业出版社,2013:30.
[7] 张凌燕,叶金云,王友慧,等.配合饲料中不同蛋白质水平对日本沼虾生长的影响[J].上海水产大学学报,2008,17(6):668-673.
[8] 谢国驷,蔡永祥,徐维娜,等.饲料蛋白水平对日本沼虾生长、消化酶和免疫酶的影响[J].江苏农业学报,2007,23(6):612-617.
[9] 虞冰如,沈竑.日本沼虾饲料最适蛋白质、脂肪含量及能量蛋白比的研究[J].水产学报,1990,14(4):821-827.
[10] 孙丽慧,沈斌乾,陈建明,等.日本沼虾对饲料赖氨酸的需要量研究[J].上海海洋大学学报,2013,22(1):100-104.
[11] 郭建林,陈建明,孙丽慧,等.日本沼虾幼虾对饲料中锌的需求量[J].动物营养学报,2013,25(3):661-668.
[12] AOAC.Official methods of association of official chemists[S].16th ed.Arlington,VA:AOAC International,1995.
[13] 王雷,李光友,毛远兴.口服免疫药物后中国对虾某些血淋巴因子的测定及方法研究[J].海洋与湖沼,1995,26(1):34-41.
[14] 刘伟,文华,吴建开,等.高铜饲料对凡纳滨对虾生长、肌肉和肝胰脏铜积累的影响[J].淡水渔业,2008,38(2):12-16.
[15] SUN SM,QIN JG,YU N,et al.Effect of dietary copper on the growth performance,non-specific immunity and resistance to Aeromonas hydrophila of juvenile Chinese mitten crab,Eriocheir sinensis[J].Fish &Shellfish Immunology,2013,34(5):1195-1201.
[16] 周萌,王安利,曹俊明.饲料中不同形式的铜及添加量对凡纳滨对虾(Litopenaeus vannamei)生长性能、血清铜蓝蛋白和生长激素水平的影响[J].海洋与湖沼,2010,41(4):577-562.
[17] 宋维彦,王秀敏,靳桂双.铁铜锌对凡纳滨对虾生长和非特异免疫的影响[J].江苏农业科学,2011,39(6):376-399.
[18] 王维娜,王安利,刘存岐,等.饵料中的铜含量对中国对虾生长及体内铜、锌和铁含量的影响[J].水产学报,1997,21(3):258-262.
[19] 郭志勋,陈毕生,徐力文,等.饲料中铜的添加量对南美白对虾生长、血液免疫因子及组织铜的影响[J].中国水产科学,2003,10(6):526-528.
[20] CERENIUS L,SODERHALL K.The prophenoloxidase activating system in invertebrates[J].Immunological Reviews,2004,198(1):116-126.
[21] 刘凯.南美白对虾和罗氏沼虾血细胞中酚氧化酶活力的比较[J].水生态学杂志,2009,2(4):103-107.
[22] 刘媛,王维娜,王安利,等.牛磺酸对日本沼虾生长及酚氧化酶活性的影响[J].淡水渔业,2005,35(2):28-30.
[23] 董晓慧,杨原志,郑石轩,等.饲料中不同铜源和水平对凡纳滨对虾生长、免疫和组织铜含量的影响[J].大连水产学院学报,2007,22(5):377-383.
[24] 方允中,郑荣梁.自由基生物学的理论与应用[M].北京:科学出版社,2002:162-212.
[25] GATLIN D M,WILON R P.Dietary copper requirement of fingerling channel catfish[J].Aquaculture,1986,54(4):277-285.
[26] HARRIS E D.Copper as a cofactor and regulator of copper,zinc superoxide dismutase[J].Nutrition,1992,122(Suppl.3):636-640.
[27] HARI R,BURDE V,ARKING R.Immunological confirmation of elevated levels of CuZn superoxide dismutase protein in an artificially selected long-lived strain of Drosophila melanogaster[J].Experimental Gerontol,1998,33(3):227-237.
[28] GRALLA E B,KOSMAN D J.Moleculargenetics of superoxide dismutases in yeasts and related fungi[J].Advances in Genetics,1992,30:251-319.
[29] 崔立娇,张利民,王际英,等.饲料锌水平对星斑川鲽幼鱼生长、组织积累和抗氧化功能的影响[J].水产学报,2010,34(9):1420-1428.
[30] 叶超霞,刘永坚,田丽霞,等.斜带石斑鱼(Epinephelus coioides)饲料中铜的适宜添加量研究[J].海洋与湖沼,2013,44(2):461-466.
[31] WHITE S L,RAINBOW P S.On the metabolic requirements for copper and zinc in molluses and crustaceans[J].Marine Environmental Research,1986,16(3):215-229.
[32] 魏万权,李爱杰,李德尚.牙鲆幼鱼饲料中铜的适宜添加量研究[J].海洋湖沼通报,2001(2):54-59.
[33] KIM S G,KANG J C.Effect of dietary copper exposure on accumulation,growth and hematological parameters of the juvenile rockfish,Sebaste schlegeli[J].Marine Environmental Research,2004,58(1):65-82.
[34] HILTON J W.The interaction of vitamins,minerals and diet composition in the diet of fish[J].Aquaculture,1989,79(1/2/3/4):223-244.
[35] 钟山,任波,邓铭,等.几种微量元素的协同作用与拮抗作用的研究[J].湖北医科大学学报,1999,20(2):107-108.
[36] 宁红梅,葛亚明,李敬玺,等.硒锌铜交互效应对肉鸡胸肌铜、锌、铁、锰、钙含量的影响[J].郑州轻工业学院学报:自然科学版,2011,26(4):6-12.
[37] 南旭阳.Cu2+对鲫鱼体内Zn2+和血红蛋白含量影响[J].中国公共卫生,2006,22(1):62-63.
[38] 梁艺怀,金泰廙.锌或铜诱导肝脏金属硫蛋白结合锌镉比值与铜镉比值的研究[J].环境与职业医学,2010,27(1):31-33.
[39] 张养梅.微量元素在人体中的生物利用率及元素间关联的研究[D].硕士学位论文.北京:中国原子能科学研究院,2002.
猜你喜欢
当代水产(2022年7期)2022-09-20 06:23:00
当代水产(2022年6期)2022-06-29 01:12:02
江苏农业科学(2022年4期)2022-03-11 21:30:10
当代水产(2021年8期)2021-11-04 08:49:10
猪业科学(2021年3期)2021-05-21 02:05:42
科学养鱼(2021年3期)2021-04-16 01:23:16
科学养鱼(2020年5期)2020-06-18 08:38:26
海洋与渔业(2017年11期)2018-01-26 01:44:58
淡水渔业(2016年3期)2016-06-14 09:42:51
当代化工研究(2016年9期)2016-03-20 16:22:13
