
企业孵化过程知识传递机制研究
2014-03-22 08:13徐菁鸿郭燕青
沈阳师范大学学报(社会科学版) 2014年6期
徐菁鸿,郭燕青
(1.辽宁大学 商学院,辽宁沈阳 110136;2.沈阳工程学院 管理学院,辽宁 沈阳 110136)
企业孵化过程知识传递机制研究
徐菁鸿1,2,郭燕青1
(1.辽宁大学 商学院,辽宁沈阳 110136;2.沈阳工程学院 管理学院,辽宁 沈阳 110136)
在对企业孵化过程中的知识传递机制进行定量研究,采用合项思维方法,将协同学与种群生态学相结合,以企业孵化过程的自组织特性为切入点,构建了一类基础的企业孵化网络,在协同学哈肯模型的基础上,建立了多种群寄生关系模型,并对每一个节点加入知识传递时间延迟参数,然后进行模拟与仿真,在此过程中通过假设验证的方法对快弛豫参量和慢弛豫参量进行识别。在协同学中,慢弛豫参量对系统的影响起着决定性作用,即为系统的序参量。最后,在序参量功能作用的基础上,通过问题导入的方式,从种群进化的视角提出了企业孵化过程中的知识传递机制,即知识集群的构建、模式及边界。
企业孵化;序参量;知识传递
在种群生态学中,企业孵化过程被描述为种群间的寄生关系模型[1],企业孵化器作为宿主为在孵企业提供必要的物质、能量、信息等方面的支持,在孵企业作为寄生体,在没有形成独立生存能力的情况下,寄生于企业孵化器,获取物质、能量、信息等方面的支持,完成成长过程。在协同学中,采用哈肯模型来描述企业孵化过程的自组织、非线性等特征[2-3]。在远离平衡态的开放系统由无序向有序转化的过程中,系统不同的参量在临界点处的行为大不相同:有的参数阻尼大,衰减快,对转变的整个进程没有明显的影响;有的参数出现临界无阻尼现象,衰减缓慢,在演化过程中起着主要作用。哈肯根据参数在临界点附近变化的快慢将参量分为两类:一类是阻尼大衰减快的快弛豫参量,另一类是临界无阻尼的慢弛豫参量。这两类变量同时包含在决定系统演化的微分方程中,相互联系,相互制约,相互竞争。虽然慢参量只有一个或几个,但它却控制着系统演化的整个过程,决定着演化结果所具有的结构和功能,代表系统“序”或状态,它就是表征系统有序程度的序参量[4]。序参量是协同学中的重要概念。序参量对整个系统而言,具有集中信息、概括信息及传递信息的功能,是人们了解、认识系统的一把钥匙。知识既是一类信息,由此可见,序参量是研究企业孵化过程知识传递机制不可缺少的重要参数。本文将协同学与种群生态学相结合,以孵化过程的自组织非线性演化特性作为切入点,采用合项思维方式将种群生态学中描述种群间寄生关系的非线性模型与协同学中描述自组织系统非线性相变过程的序参量作用进行融合,对每一个节点加入知识传递时间延迟参数,并对企业孵化过程进行模拟和仿真,通过假设验证的方法对快弛豫参量和慢弛豫参量进行识别,在此基础上提出企业孵化的知识集群机制。
一、文献综述
关于企业孵化过程知识服务的研究很多,其中杜运力等人认为企业孵化的实质是知识配置,知识配置过程中的主导者即是企业孵化器,于是,构建了企业孵化器知识体系,然后采用基传算法建立了知识配置的量化模型,达到优化配置孵化知识,提高企业孵化器孵化能力的目的[5]。刘红丽认为企业孵化器是一类知识网络,网络中存在知识主体,而在孵企业处于接受知识转移的地位,以弥补创业过程中的知识缺口,促进在孵企业成长,并构建了小世界网络对此进行描述[6]。于晓丹认为科技兴中小企业作为在孵对象,其知
识需求应该成为企业孵化器构建知识体系的基础,并在此基础上构建了KPESF矩阵模型,将知识库引入企业孵化器知识服务系统中来[7]。
一些学者亦采用种群生态学方法来研究企业孵化过程。李永刚等认为企业寄生属于企业共生的一个阶段,寄生企业依靠“食取”寄主企业的资源或利润而得以生存。企业寄生关系是企业孵化过程的初始阶段[8];Scillitoe等构建了企业孵化的网络模型,分析了网络体系中存在的一种商业援助关系,认为学习者依据自身的偏好通过与孵化器进行管理咨询互动将会推动新企业的孵化过程[9]。
在对企业孵化及相关过程中序参量的识别研究中,常见两种方法,一种是指标评价,另一种是假设试算。前者张涵等构建了网络强度、合作公平等7个科技企业孵化网络绩效影响因素维度,对5个城市的孵化网络成员进行了实证研究[10];Schwartz等对来自德国的150家企业的相关数据进行实证分析,研究了不同企业间合作模式的差异,并在此基础上分析了企业孵化器的专业化和多元化运行机制[11]。后者如赵玉林利用协同学的微观方法,假设快、慢两类变量,采用绝热消去法通过计算进行区分,消去快变量,得到序参量方程[3]。李靖等在绝热消去法的基础上构建了一类物流网络协同投入状态参量方程组,对序参量进行甄别[12]。
综上,在企业孵化知识传递过程中,存在两种认知,一是在孵企业是知识缺乏的,企业孵化器在知识传递过程中比在孵企业本身更了解其知识需求,所以毫无疑问成为知识传递的主导者;二是企业孵化器中的在孵企业众多,企业孵化器由于无法了解每一个在孵企业的知识需求,所以其提供的知识服务具有一定的局限性,而在孵企业由于专业性更强,其知识需求应主导知识传递过程。至此,本文拟从定量的角度,采用序参量识别的方法,构建多种群寄生模型,通过假设模拟的方法,来找出知识传递过程中的主导者。
二、企业孵化知识传递过程的模拟与仿真
1.建模基础
(1)企业孵化知识传递过程的特点:
第一,企业孵化过程中,知识传递过程具有协同性。
第二,企业孵化过程中,知识传递过程存在动态相互作用。
第三,企业孵化过程中,知识传递过程具有自组织特性,是一个非线性系统。
第四,在孵企业是一类企业集聚,其演化过程具有自组织特性,是一个非线性系统。
第五,企业孵化器与被孵企业之间存在寄生关系特点。
(2)协同学中哈肯模型
涨落是自组织过程的基本特性。涨落意味着某种时间的出现具有偶然性,由于研究者掌握的条件不充分,结果可能具有一定的不确定性。精确确定这种不确定性是一个困难的问题,因此可以对此进行宏观讨论,可以用下述方程来进行描述;
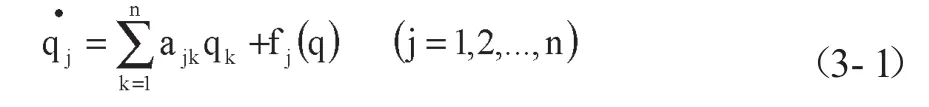
其中,fj(q)是q的一组非线性函数。对于稳定的非平衡定态点,其线性项系数矩阵的本征值具有负实部,即矩阵(ajk)是负定的。于是,进行线性变换,用一组新坐标使(ajk)对角化。
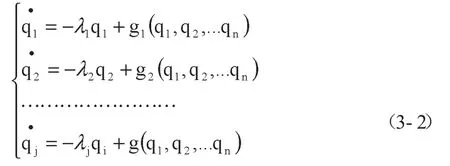
在运用哈肯模型描述系统演化过程时,往往初步假设某一参量为序参量[3],然后进行求解试算,判断假设是否成立。
(3)种群生态学中的多种群生态系统模型
种群生态系统具有自组织特性,多种群生态系统模型可以描述多种群之间的复杂关系,属于非线性系统,当解析解存在虚部时,可以通过观察其数值解图来分析系统的稳定性[13]。因此,本文根据哈肯模型的一般结构对可以描述多种群之间非线性自组织演进的种群寄生关系模型进行重构,实现本文概念框架所描述的企业孵化过程。
(4)时滞种群模型的优点
考虑到具有时滞的种群模型所考虑系统的未来状态,不仅取决于当前的状态,而且还依赖于系统过去的状态[14]。在许多情况下,这一事实较微分方程更真实地反映着实际情况[15],所以,本文拟构建带时滞模型来对企业孵化知识传递过程的序参量进行识别,时滞参数代表不同主体在知识传递过程中的响应时间。
综上,本文拟将种群生态学中的寄生关系模型,结合企业孵化过程的特点,按照式(3-2)进行重构,运用种群生态学的方法进行求解,运用协同学序参量识别方法对求解结果进行分析。
2.概念模型
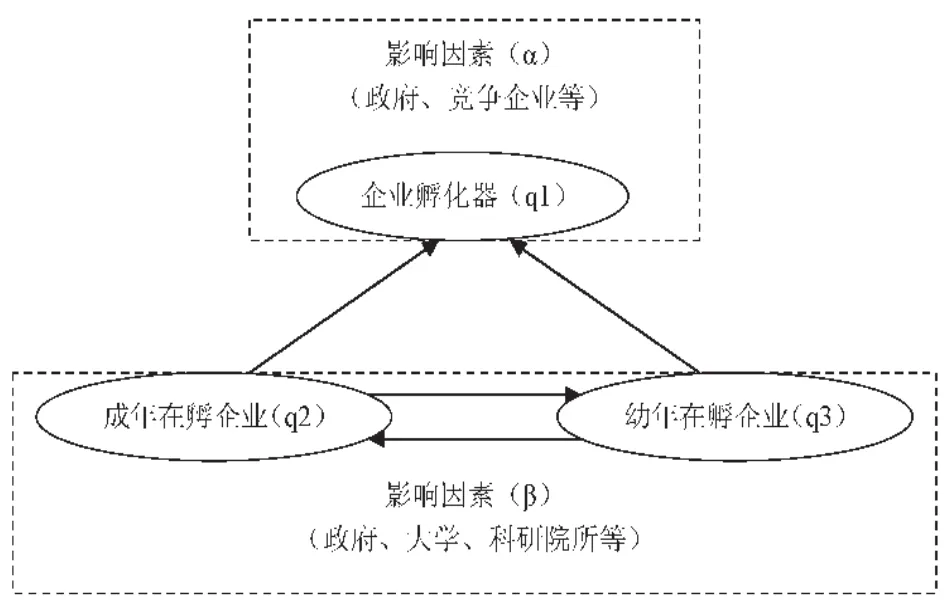
图3.1 企业孵化过程概念模型
(1)概念模型说明
上述模型由企业孵化器、成年在孵企业和幼年在孵企业三个种群构成企业孵化网络,企业孵化器是宿主,在孵企业是寄生体,成年在孵企业与幼年在孵企业存在资源竞争。在种群生态学中箭头的运用表示两种群之间的关系,信息传递默认为在两节点中是双向传递的。
(2)假设条件
假设1:影响因素对企业孵化器与在孵企业的作用是有差异的;
假设2:从目前的研究来看,企业孵化器的知识传递过程受到融资难[16]、人才匮乏[17]、政策支持有限[18]等因素的影响,所以表示企业孵化器的人工干预参数α在模拟过程中假设为负值。在孵企业的人工干预参数β在模拟过程中假设为正值;
假设3:序参量衰减缓慢的显著特性在本文中通过响应时间的存在和长短来进行描述与区分。成年在孵企业(г2)与幼年在孵企业(г3)的孵化时间差异主要表现为对知识传递的响应时间上,假设成年在孵企业处于即将脱离企业孵化器的边缘,孵化过程接近尾声,已基本完成成长过程,故对知识传递的响应时间较短,设г2=0.2,幼年在孵企业对知识传递的响应时间较长,设г3=1;企业孵化器作为孵化网络中最为成熟的主体,其响应时间最短,故设г1=0.1。
3.建模与仿真
首先,建立基础模型,其中q1代表企业孵化器,q2代表成年在孵企业、q3代表幼年在孵企业。
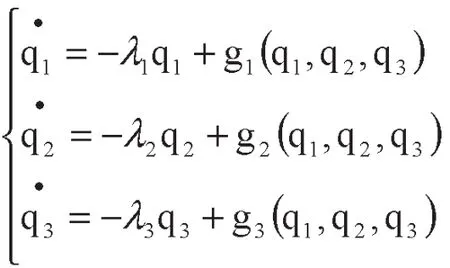
λi——种群知识的增长率或死亡率;qi——种群知识产出
然后采用假设试算的方法,进行快弛豫参量和慢弛豫参量的识别。
赋初值,设λ1=30,λ2=20,λ3=10,α=8,β=10,a=4,b=4.5,c=3,d=3.5
(1)假设企业孵化器q1为慢弛豫参量,在孵企业q2、q3为快弛豫参量,考虑到慢弛豫参量具有衰减缓慢的特点,所以设q1的时滞г1=0.1,q2、q3无时滞,建立模型。
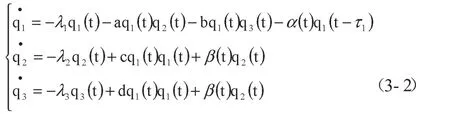
a、b、c、d——种群间相互作用系数
用matlab7.0对式(3-2)过程进行模拟仿真,为了更好地考察序参数对系统演进的作用,设初始值均为0。观察数值解图,发现三种群在上述参数作用下形成了新的状态,见图3.2(a)三种群在经历不同时间的振荡后,达到新的稳态。其中企业孵化器q1衰减相对缓慢,与假设的结果相同,证明q1可能是慢弛豫参量。而q2、q3均出现快速衰减的趋势,证明q2、q3可能是快驰豫参量。
(2)假设企业孵化器q1为快弛豫参量,在孵企业q2、q3为慢弛豫参量,设 q2、q3的时滞分别为г2=0.2,г3=1,建立模型。
(3-3)
用matlab7.0对式(3-3)过程进行模拟仿真,设初始值均为0。观察数值解图(x轴表示时间,y轴表示产出),见图3.2(b),发现三种群在经历不同时间的振荡后,q1在模拟时间段内始终没有达到新的稳态,而q2在达到稳态一段时间后,重新进入振荡状态,q3则达到了稳态。其中企业孵化器q1衰减相对缓慢,与假设的结果相反,证明q2、q3可能是慢弛豫参量的假设不成立。
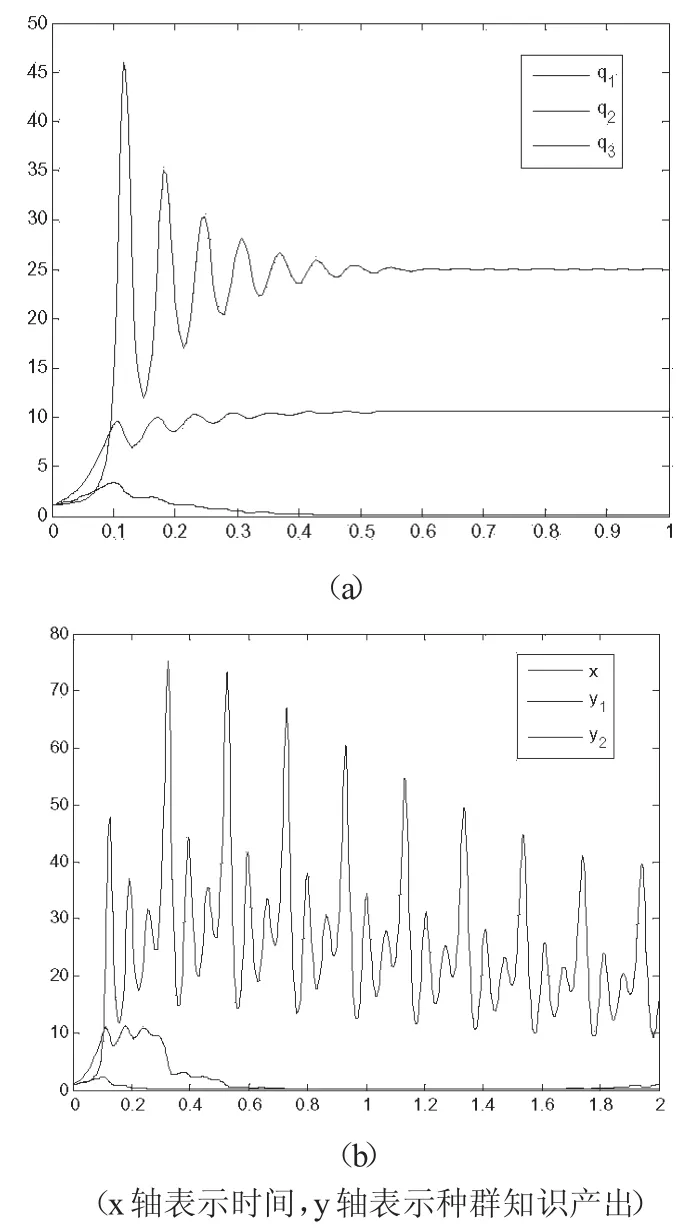
图3.2 企业孵化过程模拟与仿真结果(初始值从0开始)
通过上述模拟仿真与分析,发现企业孵化器可能是整个孵化过程的慢弛豫参量,即序参量。其控制着系统演化的整个过程,决定着演化结果所具有的结构和功能,也决定着系统“序”状态的实现。企业孵化器实质上是一种企业培育机制,下面将着重围绕企业孵化器这一序参量对企业孵化过程的知识传递机制进行分析。
三、基于问题导入的知识传递机制
本文主要研究三个问题:企业孵化过程中的知识传递机制与集群机制是否可以融合;知识传递机制与集群机制融合的模式是否存在;知识集群机制下,企业孵化器的边界如何界定。
问题一:企业孵化过程中的知识传递机制与集群机制是否可以融合。德兰根(DeLangen,2002)认为集群机制是集群内企业各种关系的总合,是一种协调、交互机制。企业孵化器作为一种企业培育机制,可以与集群机制相融合。知识传递机制是企业孵化过程中的重要影响因素,通过知识传递,可以有效地提高企业孵化过程中知识的融合、协调与互补,提高在孵企业的响应速度。产学研合作既包含相关机构在时间空间维度的聚集,又包含相关机构之间复杂的知识传递过程,所以可以作为企业孵化过程中集群机制与知识传递机制融合的切入点。企业孵化与产学研合作密不可分,产学研合作的知识传递过程对企业孵化过程中的知识创新起到从量变到质变的推动作用,知识传递表现为一类不确定随机事件,可能导致企业孵化过程临界状态的改变,例如新的在孵企业的入驻、企业孵化网络中新技术的研发等,而成为企业孵化过程有序结构衍生的内部诱因,使孵化过程偏离原来的状态或轨道,产出形成振荡。振荡的结果有两种,一种是在反馈过程中被放大为“巨涨落”,进而形成耗散结构,而另一种是在人工干预下很可能跃迁到新的稳态,例如图3.2(a)中的人工干预结果是使三种群的演进向稳态发展,而图3.2(b)的人工干预结果则是在短时间内观察不到企业孵化期稳态趋势的出现。因此,集群机制与知识传递机制的融合符合企业孵化过程的客观特点,可以在此基础上对企业孵化的结果做到能观能控。
问题二:知识传递机制与集群机制融合的模式是否存在。对上一个问题,知识传递过程中的不确定性所导致的耗散,可以参考日本创新研究的一类观点,即知识创新型集群[19]。知识创新型集群包括核心大学、公共研究机构、地方政府、地方研发型企业等多种主体,通过契约研究、联合研究等方式推动知识传递、知识创新等活动向前发展。其主体构成比三螺旋理论中Etzkowitz和Leydesdorff(2000)提出的大学、产业、政府三方创新模式以及社会网络理论下Smilor和Gill(1986)提出的大学、主要企业、创新企业、政府支持、专业支持和其他支持六个部分构成的创业网络模式更加丰富。日本知识集群模式最明显的特征是知识集群本部和知识集区事业推进工作组作为一类子系统而存在,增强了构成主体的功能性。
问题三:知识集群机制下,企业孵化器的边界如何界定。目前,从有限的关于知识集群的研究中,一些学者结合大学科技园、创业平台等实例,对知识集群作出了解释,Tallman认为知识集群中企业由于地理临近而能形成一支的架构性知识和系统性知识,从而实现创新性知识在区域中转移。如果将知识集群视为一类企业孵化器,则其运行机理与序参量衰减振荡缓慢的功能要求是可以一致的:知识传递过程中,会形成知识积累,这种积累可以视为一种能量积累,成为振荡的诱因,可以通过调整知识集群的功能和内部结构的方式来疏散和传递能量,如通过目标一致(共同研究课题)对系统偏离的不确定性进行约束,在此过程中孵化过程自然发生,日本的地方政府会联合本地产学研主体对知识集群功能和结构演进过程中的进一步规划进行评估,确保功能和内部结构演进的效果。这种知识集群的边界比传统的产业集群、孵化平台的边界更为宽广,资源(子系统)更加丰富,形式更加灵活,能够更好地体现知识资本在企业孵化过程中的作用。
[1]董秋云.生态视角下的中小企业集聚成长模式探讨[J].科技进步与对策,2008,25(3):108-111.
[2]张力.企业孵化器研究前沿与突破方向探析[J].外国经济与管理,2010,32(6):17-22.
[3]赵玉林.主导性高技术产业成长机制论[M].北京:科学出版社,2012:3.
[4]李柏洲,刘建波.企业进化系统的序参量探讨[J].中国科技论坛,2005(4):85-87.
[5]杜运力,陈智高.基于遗传算法的孵化器知识配置模型研究[J].科技进步与对策,2010,27(7):140-144.
[6]刘红丽.企业孵化器知识网络中的知识转移研究[D].华东理工大学,2014.
[7]于晓丹.企业孵化器知识网络中的知识转移研究[D].大连理工大学,2010.
[8]李永刚,施俊杰.企业孵化衍生与共生机理[J].南昌大学学报:人文社会科学版,2013,44(1):64-69.
[9]Scillitoe,J.andChakrabarti,A.K.Theroleofincubatorinteractions inassistingnewventures,Technovation,2010(30):155-167.
[10]张涵,赵黎明,乌兰娜.科技企业孵化网络关键影响因素与合作绩效的关系研究——基于SEM模型 [J].科学管理研究,2013,31(6):114-117.
[11]Schwartz,M.andHornych,C.Cooperationpatternsofincubator firmsandtheimpactofincubatorspecialization:empirical evidencefromGermany’,Technovation,2010(30):485-495.
[12]李靖,张永安.基于绝热消去法的物流网络协同序参量甄别研究[J].统计与决策,2012(7):183-185.
[13]夏安邦.系统建模理论与方法[M].北京:机械工业出版社,2008:33-45.
[14]焦宝聪.常微分方程通解[M].北京:清华大学出版社,2008.
[15]理查德·L.达夫特.组织生态学理论与应用组织理论与设计(第9版)[M].北京:清华大学出版社,2008:24-48.
[16]宋清.科技型创业企业孵化资源配置效率实证研究[J].中国科技论坛,2013(10):87-92.
[17]温凤媛.大企业开放式技术创新模式内涵及构成要素[J].沈阳师范大学学报:社会科学版,2013,37(6):81-83.
[18]林德昌,廖蓓秋,陆强,等.科技企业孵化器服务创新影响因素研究[J].科学学研究,2010,28(6):920-925,901.
[19]日本文部科学省.2010.平成22年度地域イノベーションクラスタープログラムパンフレット.http://www.mext.go. jp/a_menu/kagaku/chiiki/budget/1297966.htm,2011.
【责任编辑 王凤娥】
F125.1
A
1674-5450(2014)06-0066-04
2014-09-13
徐菁鸿,女,山东莱州人,辽宁大学经济学博士研究生,沈阳工程学院教师;郭燕青,男,吉林梅河口人,辽宁大学教授,博士研究生导师。
猜你喜欢
军事运筹与系统工程(2019年4期)2019-09-11
电子制作(2018年11期)2018-08-04
中国交通信息化(2017年3期)2017-06-08
知识就是力量(2017年2期)2017-01-21
华东科技(2016年5期)2016-03-15
物理实验(2015年9期)2015-02-28
创业家(2015年6期)2015-02-27
创业家(2015年6期)2015-02-27
数学年刊A辑(中文版)(2014年4期)2014-10-30
声学技术(2014年2期)2014-06-21
